
Oak Species Quercus robur L. and Quercus petraea Liebl. Identification Based on UHPLC-HRMS/MS Molecular Networks


Abstract
:
1. Introduction
2. Results and Discussion
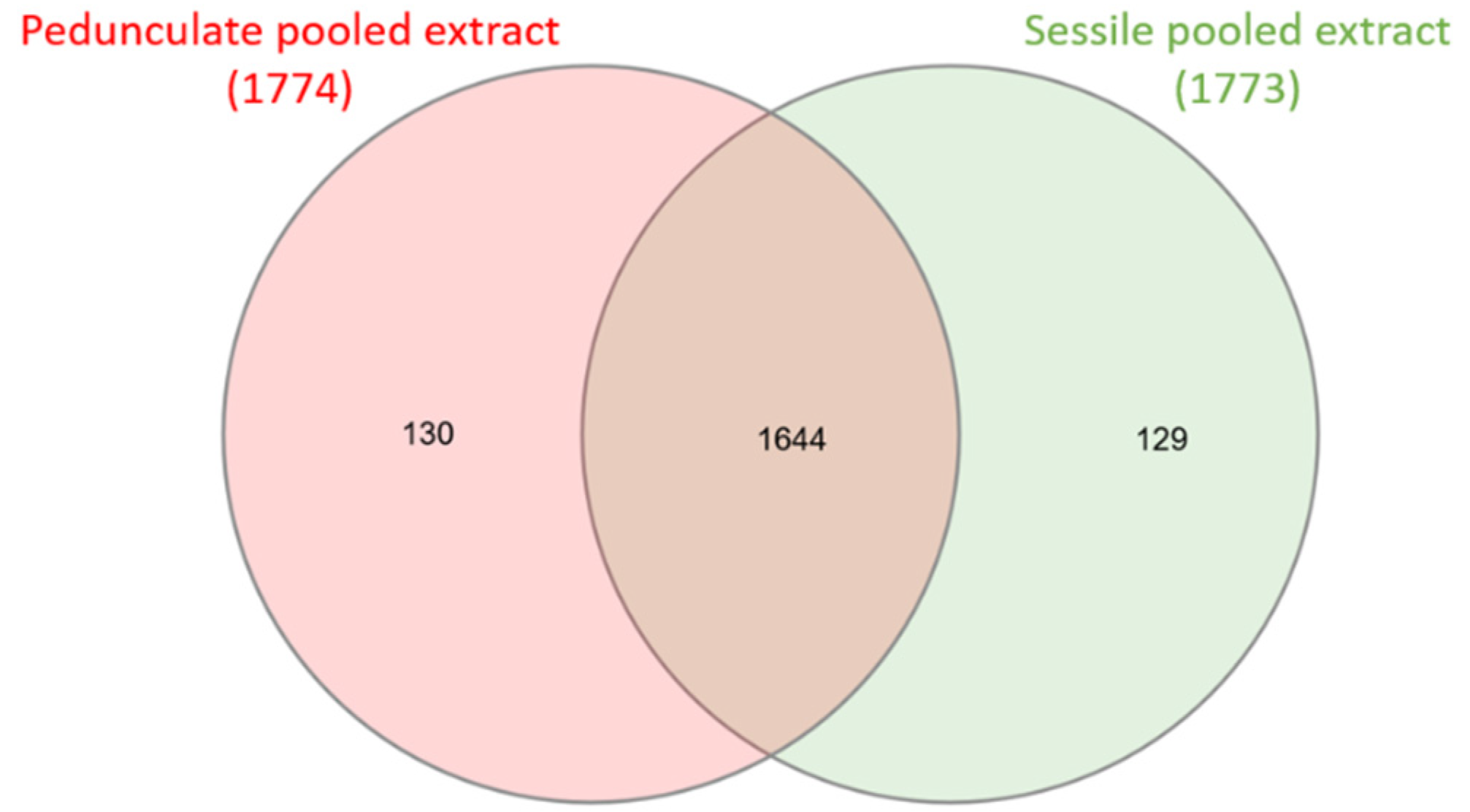
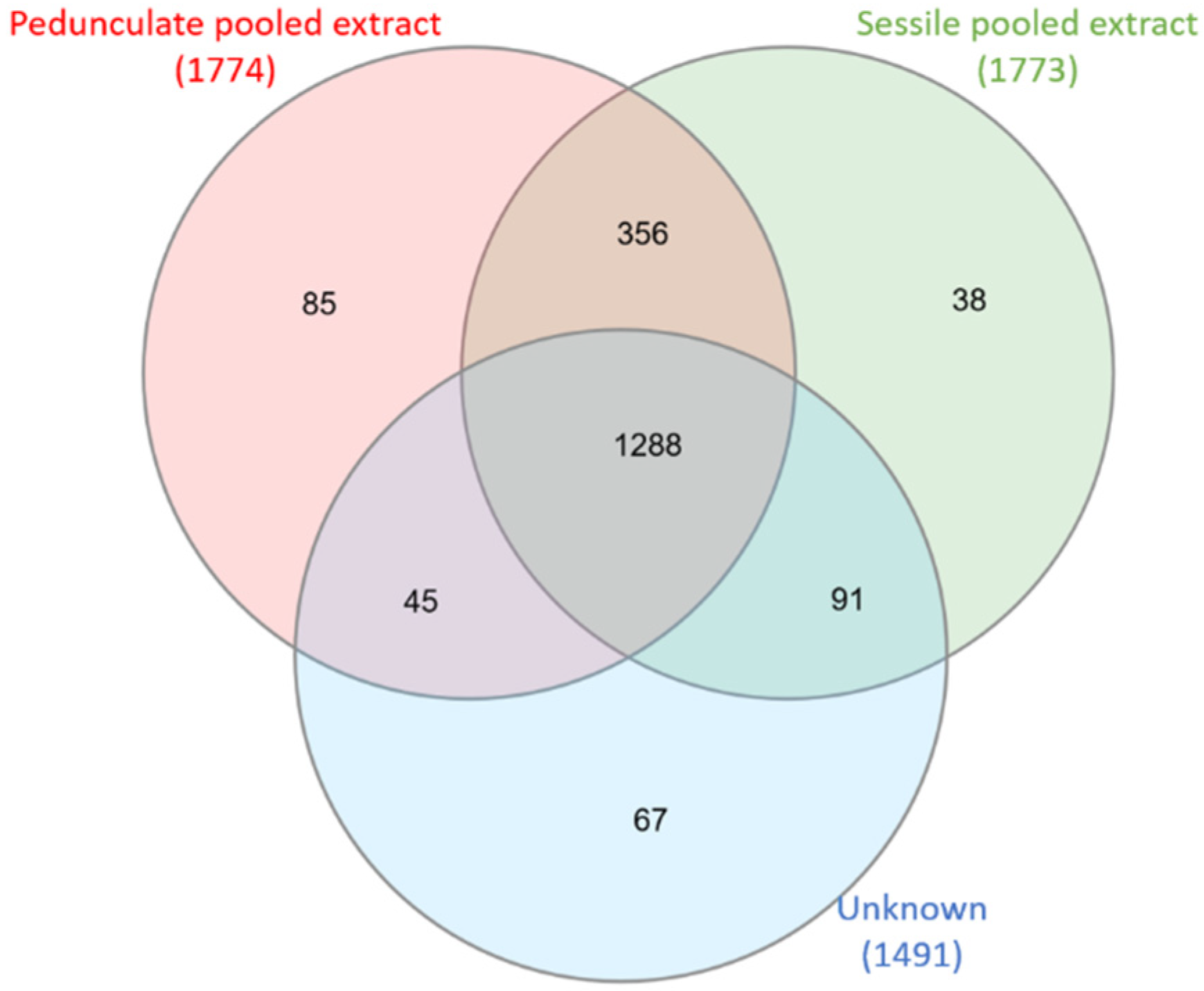
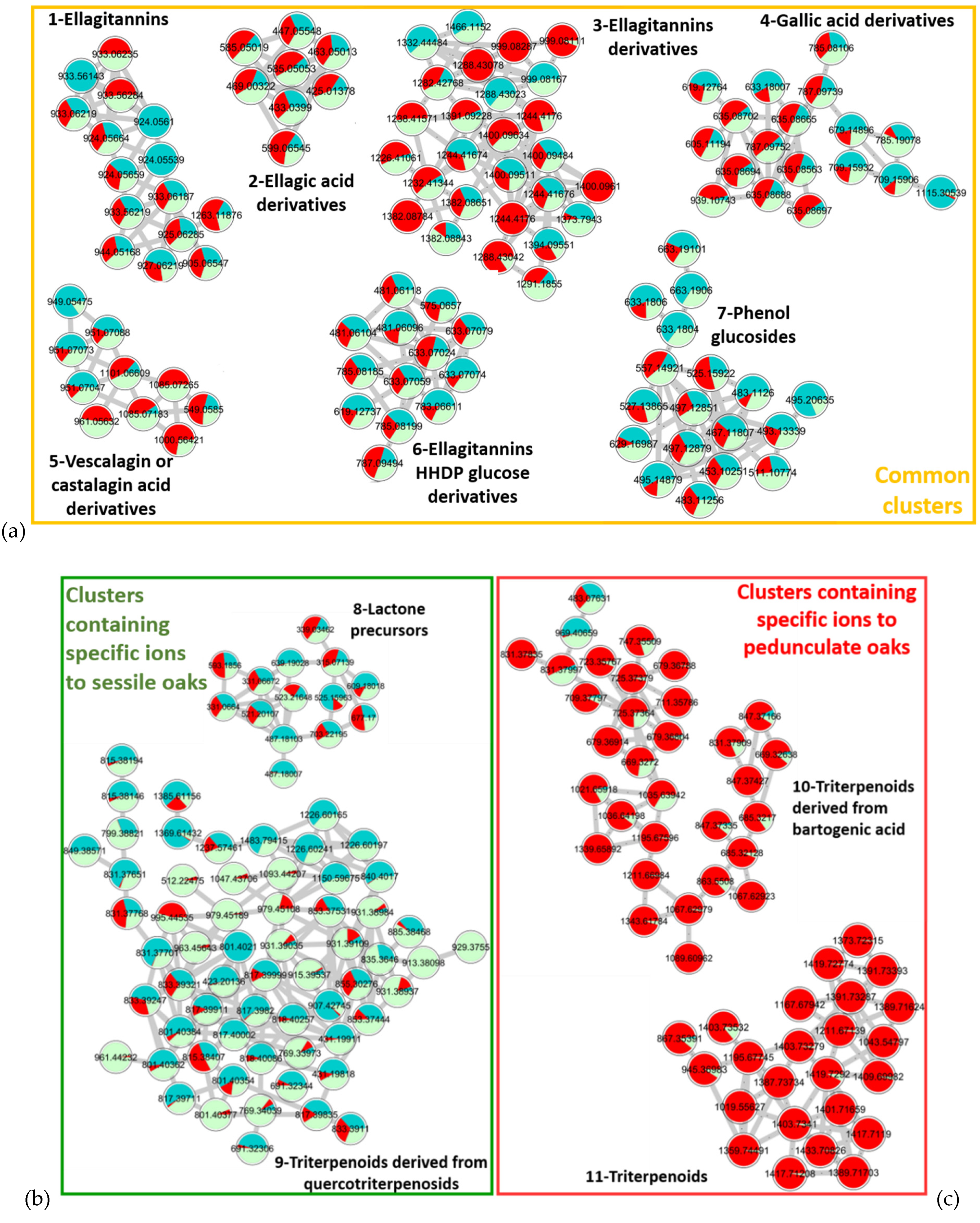
2.1. Comparison of Sessile and Pedunculate Oak Chemical Composition
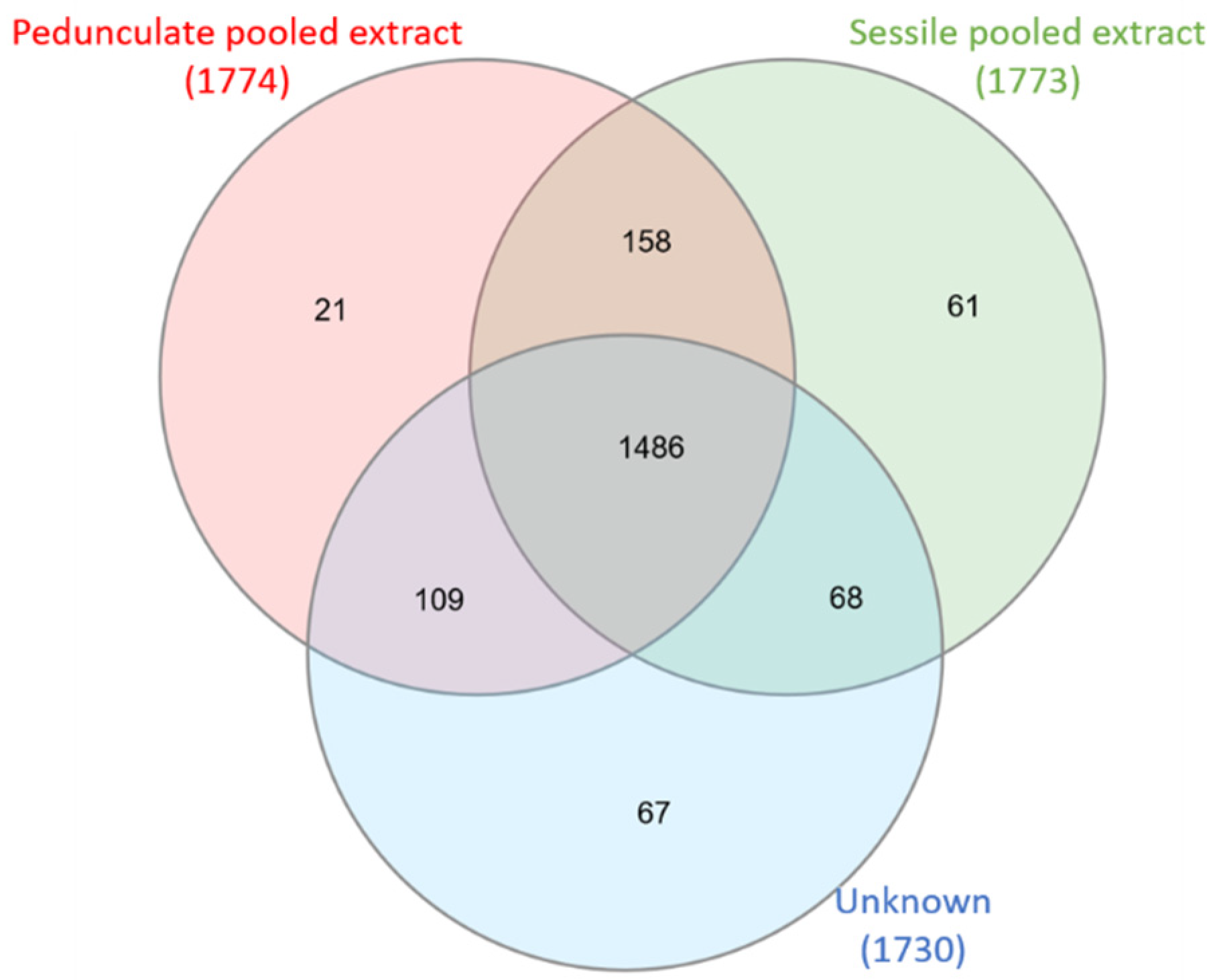
2.2. Species Assignment of an Unknown Sample
2.2.1. Addition of Pedunculate Oak Individuals to the Network
2.2.2. Addition of Sessile Oak Individuals to the Network
3. Materials and Methods
3.1. Chemicals
3.2. Oak Wood Extract Preparation
3.2.1. Oak Wood Sampling
3.2.2. Genetic Analyses for Species Assignation
3.2.3. Oak Wood Extraction
3.3. UHPLC-HRMS/MS Analysis
3.4. Venn Diagrams
3.5. Molecular Network Design
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kremer, A.; Dupouey, J.L.; Deans, J.D.; Cottrell, J.; Csaikl, U.M.; Finkeldey, R.; Espinel, S.; Jensen, J.S.; Kleinschmit, J.; Van Dam, B.; et al. Leaf morphological differentiation between Quercus robur and Quercus petraea is stable across western European mixed oak stands. Ann. For. Sci. 2002, 59, 777–787. [Google Scholar] [CrossRef]
- Boidron, J.-N.; Chatonnet, P.; Pons, M. Influence du bois sur certaines substances odorantes des vins. OENO One 1988, 22, 275–294. [Google Scholar] [CrossRef]
- Jarauta, I.; Cacho, J.; Ferreira, V. Concurrent Phenomena Contributing to the Formation of the Aroma of Wine during Aging in Oak Wood: An Analytical Study. J. Agric. Food Chem. 2005, 53, 4166–4177. [Google Scholar] [CrossRef] [PubMed]
- Spillman, P.J.; Sefton, M.A.; Gawel, R. The effect of oak wood source, location of seasoning and coopering on the composition of volatile compounds in oak-matured wines. Aust. J. Grape Wine Res. 2008, 10, 216–226. [Google Scholar] [CrossRef]
- Zamora, F. Barrel aging; Types of wood. In Red Wine Technology; Academic Press: Cambridge, MA, USA, 2019; pp. 125–147. [Google Scholar] [CrossRef]
- Zhang, B.; Cai, J.; Duan, C.-Q.; Reeves, M.J.; He, F. A Review of Polyphenolics in Oak Woods. Int. J. Mol. Sci. 2015, 16, 6978–7014. [Google Scholar] [CrossRef]
- Glabasnia, A.; Hofmann, T. Sensory-Directed Identification of Taste-Active Ellagitannins in American (Quercus alba L.) and European Oak Wood (Quercus robur L.) and Quantitative Analysis in Bourbon Whiskey and Oak-Matured Red Wines. J. Agric. Food Chem. 2006, 54, 3380–3390. [Google Scholar] [CrossRef] [PubMed]
- Chira, K.; Anguellu, L.; Da Costa, G.; Richard, T.; Pedrot, E.; Jourdes, M.; Teissedre, P.-L. New C-Glycosidic Ellagitannins Formed upon Oak Wood Toasting; Identification and Sensory Evaluation. Foods 2020, 9, 1477. [Google Scholar] [CrossRef]
- Winstel, D.; Marchal, A. Lignans in Spirits: Chemical Diversity, Quantification, and Sensory Impact of (±)-Lyoniresinol. Molecules 2018, 24, 117. [Google Scholar] [CrossRef] [Green Version]
- Marchal, A.; Cretin, B.N.; Sindt, L.; Waffo-Téguo, P.; Dubourdieu, D. Contribution of oak lignans to wine taste: Chemical identification, sensory characterization and quantification. Tetrahedron 2015, 71, 3148–3156. [Google Scholar] [CrossRef]
- Marchal, A.; Génin, E.; Waffo-Téguo, P.; Bibès, A.; Da Costa, G.; Mérillon, J.-M.; Dubourdieu, D. Development of an analytical methodology using Fourier transform mass spectrometry to discover new structural analogs of wine natural sweeteners. Anal. Chim. Acta 2015, 853, 425–434. [Google Scholar] [CrossRef]
- Marchal, A.; Waffo-Téguo, P.; Génin, E.; Mérillon, J.-M.; Dubourdieu, D. Identification of New Natural Sweet Compounds in Wine Using Centrifugal Partition Chromatography–Gustatometry and Fourier Transform Mass Spectrometry. Anal. Chem. 2011, 83, 9629–9637. [Google Scholar] [CrossRef] [PubMed]
- Gammacurta, M.; Waffo-Teguo, P.; Winstel, D.; Cretin, B.N.; Sindt, L.; Dubourdieu, D.; Marchal, A. Triterpenoids from Quercus petraea: Identification in Wines and Spirits and Sensory Assessment. J. Nat. Prod. 2019, 82, 265–275. [Google Scholar] [CrossRef]
- Vivas, N. Modélisation et calcul du bilan des apports d’oxygène au cours de l’élevage des vins rouges. IV-Elevage des vins rouges en conditions d’oxydations ménagées controlées. Progres. Agric. Vitic. 1999, 116, 305–311. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Glories, Y.; Maujean, A. Handbook of Enology, Volume 2: The Chemistry of Wine Stabilization and Treatments; John Wiley & Sons: Hoboken, NJ, USA, 2021; ISBN 978-1-119-58776-7. [Google Scholar]
- Del Alamo-Sanza, M.; Nevares, I. Oak wine barrel as an active vessel: A critical review of past and current knowledge. Crit. Rev. Food Sci. Nutr. 2017, 58, 2711–2726. [Google Scholar] [CrossRef]
- Prida, A.; Boulet, J.-C.; Ducousso, A.; Nepveu, G.; Puech, J.-L. Effect of species and ecological conditions on ellagitannin content in oak wood from an even-aged and mixed stand of Quercus robur L. and Quercus petraea Liebl. Ann. For. Sci. 2006, 63, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Prida, A.; Ducousso, A.; Petit, R.J.; Nepveu, G.; Puech, J.-L. Variation in wood volatile compounds in a mixed oak stand: Strong species and spatial differentiation in whisky-lactone content. Ann. For. Sci. 2007, 64, 313–320. [Google Scholar] [CrossRef]
- Doussot, F.; De Jéso, B.; Quideau, S.; Pardon, P. Extractives Content in Cooperage Oak Wood during Natural Seasoning and Toasting; Influence of Tree Species, Geographic Location, and Single-Tree Effects. J. Agric. Food Chem. 2002, 50, 5955–5961. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Savill, P.S. Variation of heartwood phenolics and oak lactones between the species and phenological types of Quercus petraea and Q. robur. Forestry 1996, 69, 47–56. [Google Scholar] [CrossRef]
- Kremer, A.; Petit, R.-J.; Ducousso, A. Biologie Évolutive et Diversité Génétique des Chênes Sessile et Pédonculé; INRAE: Cestas, France, 2002; p. 111. [Google Scholar] [CrossRef] [Green Version]
- Muir, G.; Fleming, C.C.; Schltterer, C. Species status of hybridizing oaks. Nature 2000, 405, 1016. [Google Scholar] [CrossRef] [PubMed]
- Guichoux, E.; Petit, R. Méthode de traçabilité géographique des bois de chêne; Déclaration d’invention (n DIRV-13-00566); INPI: Bordeaux, France, 2014. [Google Scholar]
- Marchal, A.; Prida, A.; Dubourdieu, D. New Approach for Differentiating Sessile and Pedunculate Oak: Development of a LC-HRMS Method to Quantitate Triterpenoids in Wood. J. Agric. Food Chem. 2016, 64, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Marchal, A.; Dubourdieu, D. Method for Identifying the Oak Species of an Oak Wood Sample. U.S. Patent 20160011166A1, 14 January 2016. [Google Scholar]
- Buche, G.; Colas, C.; Fougère, L.; Giordanengo, T.; Destandau, E. Untargeted UHPLC-Q-TOF-HRMS based determination of discrimating compounds for oak species Quercus robur L. and Quercus petraea Liebl. identification. Phytochem. Anal. 2020. [Google Scholar] [CrossRef]
- Gougeon, R.D.; Lucio, M.; De Boel, A.; Frommberger, M.; Hertkorn, N.; Peyron, D.; Chassagne, D.; Feuillat, F.; Cayot, P.; Voilley, A.; et al. Expressing Forest Origins in the Chemical Composition of Cooperage Oak Woods and Corresponding Wines by Using FTICR-MS. Chem. Eur. J. 2008, 15, 600–611. [Google Scholar] [CrossRef]
- Vayssières, M.P.; Plant, R.E.; Allen-Diaz, B.H. Classification trees: An alternative non-parametric approach for predicting species distributions. J. Veg. Sci. 2000, 11, 679–694. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.; Sanchez, L.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; A Kapono, C.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular Networking as a Dereplication Strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Simón, M.B.F.; Sanz, M.; Cadahía, E.; Poveda, A.P.; Broto-Cartagena, M. Chemical Characterization of Oak Heartwood from Spanish Forests of Quercus pyrenaica (Wild.). Ellagitannins, Low Molecular Weight Phenolic, and Volatile Compounds. J. Agric. Food Chem. 2006, 54, 8314–8321. [Google Scholar] [CrossRef] [PubMed]
- Vivas, N.; Glories, Y.; Bourgeois, G.; Vitry, C. The Heartwood Ellagitannins of Different Oaks (Quercus Sp.) and Chestnut Species (Castanea Sativa Mill.). Quantity Analysis of Red Wines Aging in Barrels. J. Des Sci. Tech. Tonnelerie 1996, 25–75. [Google Scholar]
- Venter, P. Analysis of Complex Tannins by Multidimensional Techniques. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2019. [Google Scholar]
- Cerulli, A.; Napolitano, A.; Hošek, J.; Masullo, M.; Pizza, C.; Piacente, S. Antioxidant and In Vitro Preliminary Anti-Inflammatory Activity of Castanea sativa (Italian Cultivar “Marrone di Roccadaspide” PGI) Burs, Leaves and Chestnuts Extracts and their Metabolite Profiles by LC-ESI/LTQOrbitrap/MS/MS. Antioxidants 2021, 10, 278. [Google Scholar] [CrossRef]
- Mämmelä, P.; Savolainen, H.; Lindroos, L.; Kangas, J.; Vartiainen, T. Analysis of Oak Tannins by Liquid Chromatography-Electrospray Ionisation Mass Spectrometry. Available online: https://0-reader-elsevier-com.brum.beds.ac.uk/reader/sd/pii/S0021967300006245?token=2868AF0C859030365A67BD9580C8D918DE45468890B2BBF50E64B69B3C16913BF46EA1E94311438DAD78BA73AE69031D&originRegion=eu-west-1&originCreation=20210628133609 (accessed on 28 June 2021).
- Cammann, J.; Denzel, K.; Schilling, G.; Gross, G.G. Biosynthesis of gallotannins: β-Glucogallin-dependent formation of 1,2,3,4,6-pentagalloylglucose by enzymatic galloylation of 1,2,3,6-tetragalloylglucose. Arch. Biochem. Biophys. 1989, 273, 58–63. [Google Scholar] [CrossRef]
- Saucier, C.; Jourdes, M.; Glories, Y.; Quideau, S. Extraction, Detection, and Quantification of Flavano-Ellagitannins and Ethylvescalagin in a Bordeaux Red Wine Aged in Oak Barrels. J. Agric. Food Chem. 2006, 54, 7349–7354. [Google Scholar] [CrossRef] [PubMed]
- Centrifugal Partition Chromatography Applied to the Isolation of Oak Wood Aroma Precursors—Elsevier Enhanced Reader. Available online: https://0-reader-elsevier-com.brum.beds.ac.uk/reader/sd/pii/S0308814613005153?token=E993B2DAF29A3BBE1B6F2EB9836E50326FC688D5DA80E6036E1EB04F1BC6F141B831A98064361AB4E5DE019E76585EDB&originRegion=eu-west-1&originCreation=20210707140737 (accessed on 7 July 2021).
- Sindt, L.; Gammacurta, M.; Waffo-Teguo, P.; Dubourdieu, D.; Marchal, A. Taste-Guided Isolation of Bitter Lignans from Quercus petraea and Their Identification in Wine. J. Nat. Prod. 2016, 79, 2432–2438. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kouno, I. Whisky Lactone Precursors from the Wood of Platycarya strobilacea. J. Nat. Prod. 1996, 59, 997–999. [Google Scholar] [CrossRef]
- Masson, E.; Baumes, R.; Le Guerneve, C.; Puech, J.-L. Identification of a Precursor of β-Methyl-γ-octalactone in the Wood of Sessile Oak (Quercus petraea (Matt.) Liebl.). J. Agric. Food Chem. 2000, 48, 4306–4309. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, Y.; Wilkinson, K.L.; Elsey, G.M.; Raunkjær, M.; Sefton, M.A. Identification of Natural Oak Lactone Precursors in Extracts of American and French Oak Woods by Liquid Chromatography—Tandem Mass Spectrometry. J. Agric. Food Chem. 2007, 55, 9195–9201. [Google Scholar] [CrossRef]
- Arramon, G.; Saucier, C.; Colombani, D.; Glories, Y. Identification of triterpene saponins in Quercus robur L. and Q. petraea Liebl. heartwood by LC-ESI/MS and NMR. Phytochem. Anal. 2002, 13, 305–310. [Google Scholar] [CrossRef]
- Pérez, A.J.; Pecio, Ł.; Kowalczyk, M.; Kontek, R.; Gajek, G.; Stopinsek, L.; Mirt, I.; Oleszek, W.; Stochmal, A. Triterpenoid Components from Oak Heartwood (Quercus Robur) and Their Potential Health Benefits. J. Agric. Food Chem. 2017, 65, 23. [Google Scholar] [CrossRef]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; Da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Feature-Based Molecular Networking in the GNPS Analysis Environment—Nature Methods. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/s41592-020-0933-6 (accessed on 5 July 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
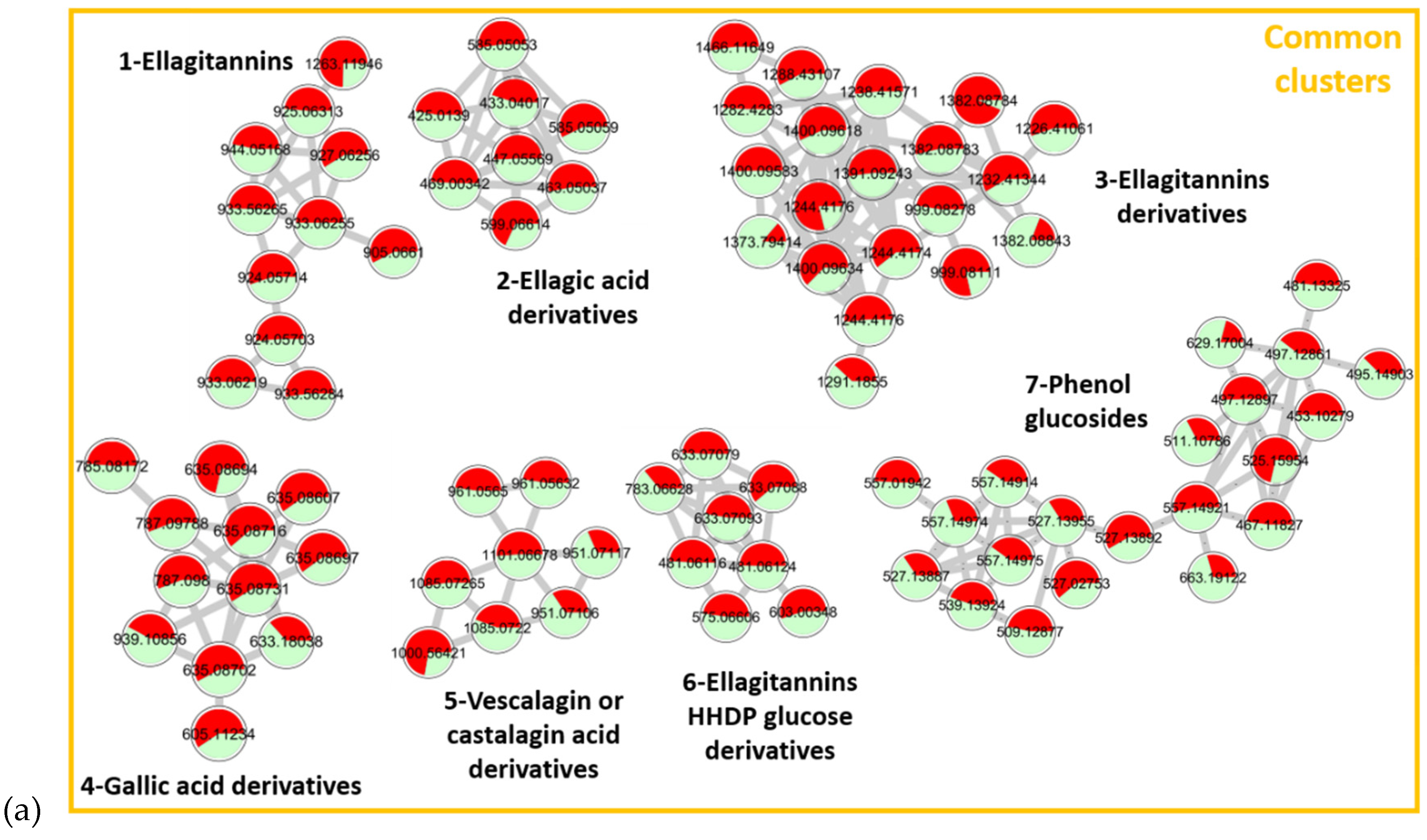
| Cluster Number | RT (min) | Measured m/z [M−H]− or [M−2H]2− | Formula [M] | Error (ppm) | MS/MS Fragments [M−H]− | Proposed Annotation for Molecules | Molecular Family of the Cluster |
|---|---|---|---|---|---|---|---|
| 1 | 4.10 | 933.06251− | C41H26O26 | −0.1 | 631.0582 C27H19O18 300.9998 C14H5O8 | Vescalagin or Castalagin [32] | Ellagitannins |
| 3.16 | 924.05712− | C82H50O51 | −1.4 | 631.0.581C27H19O18 300.9993 C14H5O8 | Roburin D or A [32] | ||
| 2 | 7.15 | 433.04011− | C19H14O12 | −1.5 | 300.9910 C14H5O8 | Ellagic acid pentoside [33] | Ellagic acid derivatives |
| 7.40 | 447.05561− | C20H16O12 | 0.8 | 300.9913 C14H5O8 | Methyl ellagic acid pentoside [34] | ||
| 3 | 3.46 | 999.08112− | C111H172O31 | 0.1 | 1065.1018 C46H33O30 975.0744 C43H27O27 933.0650 C41H25O26 631.0575 C27H19O18 | Grandinin or Roburin E derivative | Ellagitannin derivatives |
| 3.33 | 1227.41063− | x | x | 1065.1093 C46H33O30 933.0631 C41H25O26 915.0539 C41H23O25 783.0690 C34H23O22 631.0574 C27H19O18 | Glycosylated grandinin | ||
| 4 | 6.26 | 635.08731− | C27H24O18 | −0.5 | 483.0678 C20H19O14 465.0678 C20H17O13 313.0568 C13H13O9 169.0148 C7H5O5 | Trigalloyl glucose [35] | Gallic acid derivatives |
| 7.16 | 939.11581− | C34H28O22 | −0.3 | 769.0892 C34H25O21 617.0753 C34H17O12 447.0573 C20H15O12 169.0151 C7H5O5 | Penta-galloylglucose [36] | ||
| 5 | 3.57 | 1101.06671− | C48H30O31 | −0.9 | 1057.0797 C47H29O29 933.0605 C41H25O26 631.0574 C27H19O18 425.0141 C20H9O11 | Vescavaloninic acid [37] | Vescalagin or castalagin acid derivatives |
| 3.71 | 961.06411− | C42H26O27 | −0.6 | 917.0683 C41H25O25 873.0778 C40H25O23 615.0631 C27H19O17 491.062 C25H15O11 | Deoxy-carboxy vescalagin [7] | ||
| 6 | 1.89 | 481.06111− | C20H18O14 | 0.1 | 300.9951 C14H5O8 275.0161 C13H7O7 | HHDP glucose [34] | Ellagitannins HHDP glucose derivatives |
| 3.87 | 783.06621− | C34H24O22 | −0.6 | 481.0640 C20H17O14 300.9994 C14H5O8 | Pedunculagin [36] | ||
| 7 | 7.30 | 497.12891− | C22H26O13 | −0.9 | 313.0574 C13H13O9 183.0655 C9H11O4 169.0136 C7H5O5 | 3,4,5-trimethoxyphenyl-(6′-O-galloyl)-O-β-glucopyranoside [38] | Phenol glucosides |
| 5.58 | 453.10271− | C20H22O12 | 0.6 | 313.0569 C13H13O9 327.0706 C14H15O9 183.0293 C8H7O5 169.0152 C7H5O5 | 3-methoxy-4-hydroxyphenol 1-O-β-d-(6′-O-galloyl)glucopyranoside [39] | ||
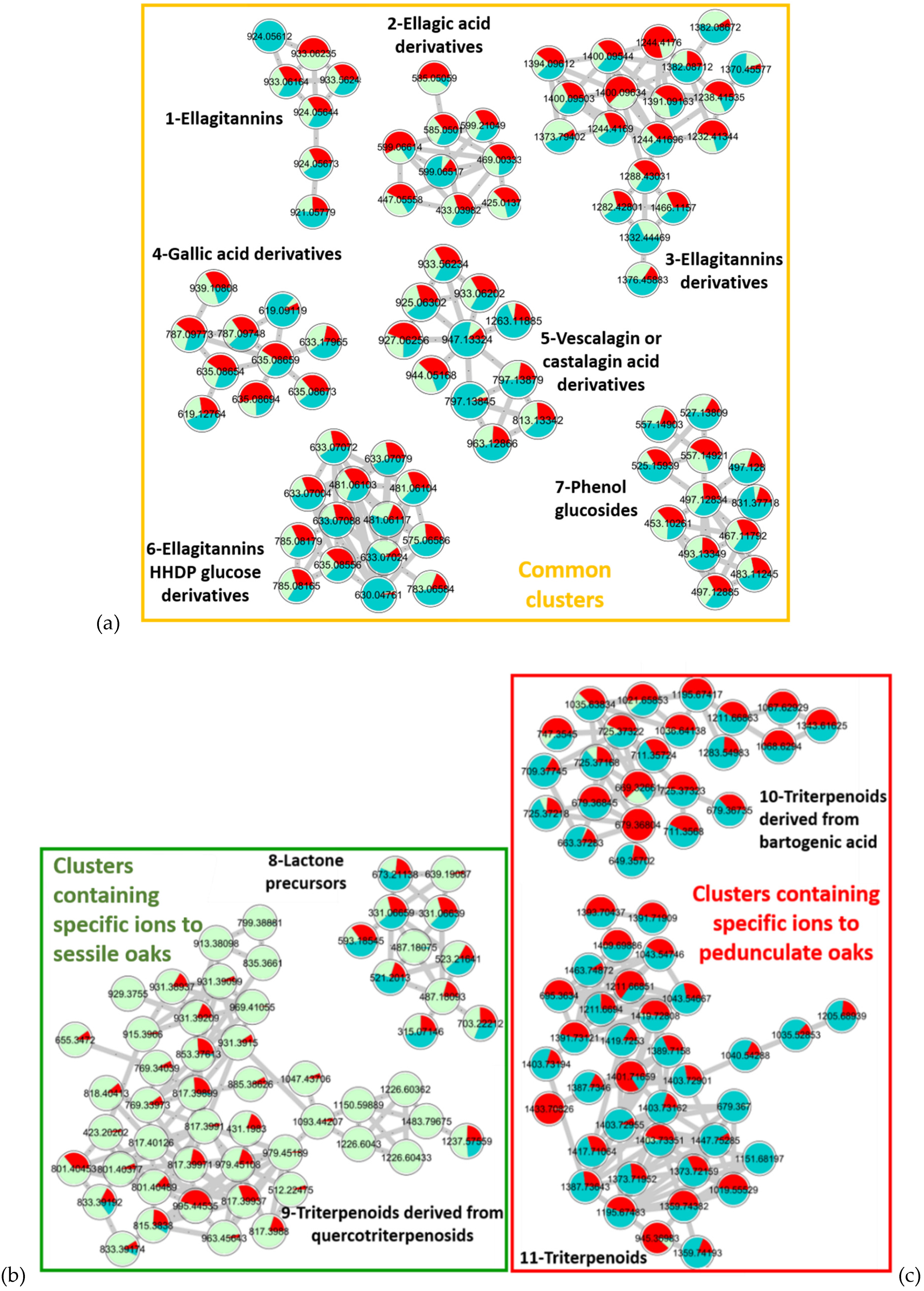
| 8 | 9.01 | 487.18211− | C22H32O12 | 0.1 | 211.0266 C9H7O6 168.0074 C7H4O5● 124.0233 C6H4O3● | 3-Methyl-4-[[6-O-(3,4,5-trihydroxybenzoyl)-β-d-glucopyranosyl]oxy] octanoic acid [40,41,42] | Lactone precursors |
| 10.40 | 639.19081− | C29H36O16 | −0.2 | 271.0454 C11H11O8 211.0244 C9H7O6 169.0145 C7H5O5 125.0249 C6H5O3 | Not identified | ||
| 9 | 10.60 | 817.39911− | C43H62O15 | −0.6 | 655.3483 C37H51O10 611.3593 C36H51O8 169.0139 C7H5O5 | Quercotriterpenosid [11,12] | Triterpenoids derived from quercotriterpenosids |
| 9.95 | 979.45181− | C49H72O20 | −0.6 | 817.4011 C43H61O15 755.4010 C42H59O12 | Quercotriterpenosid derivative [11,12] | ||
| 10 | 10.90 | 679.36801− | C36H56O12 | −0.5 | 517.3207 C30H45O7 455.3171 C29H43O4 437.3075 C29H41O3 | Glucosylated bartogenic acid [43,44] | Triterpenoids derived from bartogenic acid |
| 14.30 | 669.32721− | C37H50O11 | −2.8 | 517.3173 C30H45O7 455.3171 C29H43O4 437.3060 C29H41O3 | Galloyl bartogenic acid [43,44] | ||
| 11 | 13.50 | 1373.72481− | C72H110O25 | −0.1 | 695.3649 C36H55O13 647.3443 C35H51O11 485.2909 C29H41O6 471.3113 C29H43O5 | Roburosid B or C [43,44] | Triterpenoids |
| 13.80 | 1403.73411− | C72H110O24 | 0.2 | 679.3705 C36H55O12 517.3173 C30H45O7 | Roburosid A [43,44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buche, G.; Colas, C.; Fougère, L.; Destandau, E. Oak Species Quercus robur L. and Quercus petraea Liebl. Identification Based on UHPLC-HRMS/MS Molecular Networks. Metabolites 2021, 11, 684. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100684
Buche G, Colas C, Fougère L, Destandau E. Oak Species Quercus robur L. and Quercus petraea Liebl. Identification Based on UHPLC-HRMS/MS Molecular Networks. Metabolites. 2021; 11(10):684. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100684
Chicago/Turabian StyleBuche, Gaëlle, Cyril Colas, Laëtitia Fougère, and Emilie Destandau. 2021. "Oak Species Quercus robur L. and Quercus petraea Liebl. Identification Based on UHPLC-HRMS/MS Molecular Networks" Metabolites 11, no. 10: 684. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100684