
Identification of Indicators for Preterm Birth Using Retinoid Metabolites

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Clinical Charateristics of Subjects
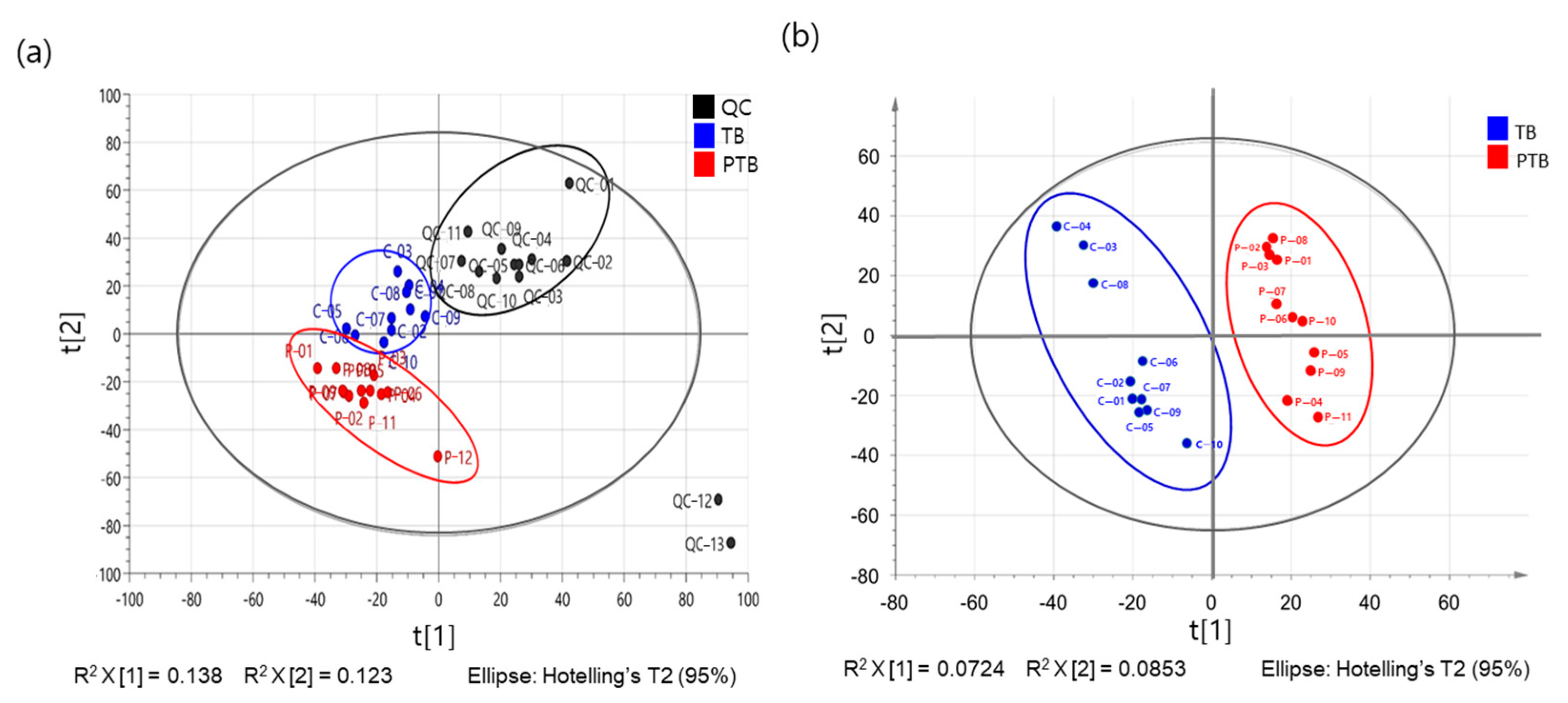
2.2. The Metabolome Profiling of Maternal Plasma Samples
2.3. The PTB Related with Metabolic Pathways
2.4. Analysis of Targeted Plasma Metabolite
2.5. Analysis of Plasma RBP
2.6. Predictive Performance for PTB
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Maternal Blood Sample Preparation
4.3. Metabolomic Profiling by LTQ-Orbitrap MS
4.4. Preperation of Standard Stock Solution for Verification of Retinoid Metabolites
4.5. Sample Preperation
4.6. LC-MS/MS Analysis
4.7. Plasma RBP Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chawanpaiboon, S.; Vogel, J.P.; Moller, A.B.; Lumbiganon, P.; Petzold, M.; Hogan, D.; Landoulsi, S.; Jampathong, N.; Kongwattanakul, K.; Laopaiboon, M.; et al. Global, regional, and national estimates of levels of preterm birth in 2014: A systematic review and modelling analysis. Lancet Glob. Health 2019, 7, e37–e46. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.K.; Velten, M. Maternal inflammation, growth retardation, and preterm birth: Insights into adult cardiovascular disease. Life Sci. 2011, 89, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Conde-Agudelo, A.; Papageorghiou, A.T.; Kennedy, S.H.; Villar, J. Novel biomarkers for the prediction of the spontaneous preterm birth phenotype: A systematic review and metaanalysis. BJOG Int. J. Obstet. Gynaecol. 2011, 118, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Sabour, S. Prediction of spontaneous preterm delivery in women with threatened preterm labour: A prospective cohort study of multiple proteins in maternal serum. BJOG Int. J. Obstet. Gynaecol. 2012, 119, 1544. [Google Scholar] [CrossRef] [PubMed]
- Laudanski, P.; Lemancewicz, A.; Kuc, P.; Charkiewicz, K.; Ramotowska, B.; Kretowska, M.; Jasinska, E.; Raba, G.; Karwasik-Kajszczarek, K.; Kraczkowski, J.; et al. Chemokines profiling of patients with preterm birth. Mediat. Inflamm. 2014, 2014, 185758. [Google Scholar] [CrossRef] [PubMed]
- Kuć, P.; Laudański, P.; Kowalczuk, O.; Chyczewski, L.; Laudański, T. Expression of selected genes in preterm premature rupture of fetal membranes. Acta Obstet. Gynecol. Scand. 2012, 91, 936–943. [Google Scholar] [CrossRef]
- Kmiecik, G.; Niklińska, W.; Kuć, P.; Pancewicz-Wojtkiewicz, J.; Fil, D.; Karwowska, A.; Karczewski, J.; Mackiewicz, Z. Fetal membranes as a source of stem cells. Adv. Med. Sci. 2013, 58, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine (US) Committee on Understanding Premature Birth and Assuring Healthy Outcomes. Preterm Birth: Causes, Consequences, and Prevention; Behrman, R.E., Butler, A.S., Eds.; National Academies Press: Washington, DC, USA, 2007; pp. 87–258.
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef] [Green Version]
- Heng, Y.J.; Liong, S.; Permezel, M.; Rice, G.E.; Di Quinzio, M.K.; Georgiou, H.M. Human cervicovaginal fluid biomarkers to predict term and preterm labor. Front. Physiol. 2015, 6, 151. [Google Scholar] [CrossRef]
- Seol, H.J.; Sung, J.H.; Seong, W.J.; Kim, H.M.; Park, H.S.; Kwon, H.; Hwang, H.S.; Jung, Y.J.; Kwon, J.Y.; Oh, S.Y. Standardization of measurement of cervical elastography, its reproducibility, and analysis of baseline clinical factors affecting elastographic parameters. Obstet. Gynecol. Sci. 2020, 63, 42–54. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Culhane, J.F. Preterm birth and periodontal disease. N. Engl. J. Med. 2006, 355, 1925–1927. [Google Scholar] [CrossRef]
- You, Y.A.; Kwon, E.J.; Choi, S.J.; Hwang, H.S.; Choi, S.K.; Lee, S.M.; Kim, Y.J. Vaginal microbiome profiles of pregnant women in Korea using a 16S metagenomics approach. Am. J. Reprod. Immunol. 2019, 82, e13124. [Google Scholar] [CrossRef]
- Rubens, C.E.; Sadovsky, Y.; Muglia, L.; Gravett, M.G.; Lackritz, E.; Gravett, C. Prevention of preterm birth: Harnessing science to address the global epidemic. Sci. Transl. Med. 2014, 6, 262sr5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudtson, E.J.; Smith, K.; Mercer, B.M.; Miodovnik, M.; Thurnau, G.R.; Goldenberg, R.L.; Meis, P.J.; Moawad, A.H.; Vandorsten, J.P.; Sorokin, Y.; et al. National Institutes of Child Health and Human Development Maternal-Fetal Medicine Units Network. Serum homocysteine levels after preterm premature rupture of the membranes. Am. J. Obstet. Gynecol. 2004, 191, 537–541. [Google Scholar] [CrossRef]
- Yoo, J.Y.; You, Y.A.; Kwon, E.J.; Park, M.H.; Shim, S.; Kim, Y.J. Differential expression and methylation of integrin subunit alpha 11 and thrombospondin in the amnion of preterm birth. Obstet. Gynecol. Sci. 2018, 61, 565–574. [Google Scholar] [PubMed]
- Fortunato, S.J.; Menon, R. Distinct molecular events suggest different pathways for preterm labor and premature rupture of membranes. Am. J. Obstet. Gynecol. 2001, 184, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 17–42. [Google Scholar] [CrossRef]
- McDonald, H.; O’loughlin, J.; Jolley, P.; Vigneswaran, R.; McDonald, P.J. Vaginal infection and preterm labour. Br. J. Obstet. Gynaecol. 1991, 98, 427–435. [Google Scholar] [CrossRef]
- Freitas, A.C.; Bocking, A.; Hill, J.E.; Money, D.M.; the VOGUE Research Group. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome 2018, 6, 117. [Google Scholar] [CrossRef]
- Son, K.A.; Kim, M.; Kim, Y.M.; Kim, S.H.; Choi, S.J.; Oh, S.Y.; Roh, C.R.; Kim, J.H. Prevalence of vaginal microorganisms among pregnant women according to trimester and association with preterm birth. Obstet. Gynecol. Sci. 2018, 61, 38–47. [Google Scholar] [CrossRef]
- Park, S.; You, Y.A.; Yun, H.; Choi, S.J.; Hwang, H.S.; Choi, S.K.; Lee, S.M.; Kim, Y.J. Cervicovaginal fluid cytokines as predictive markers of preterm birth in symptomatic women. Obstet. Gynecol. Sci. 2020, 63, 455–463. [Google Scholar] [CrossRef]
- Ansari, A.; Lee, H.; You, Y.A.; Jung, Y.; Park, S.; Kim, S.M.; Hwang, G.S.; Kim, Y.J. Identification of Potential Biomarkers in the Cervicovaginal Fluid by Metabolic Profiling for Preterm Birth. Metabolites 2020, 10, 349. [Google Scholar] [CrossRef]
- Han, Y.W.; Ikegami, A.; Bissada, N.F.; Herbst, M.; Redline, R.W.; Ashmead, G.G. Transmission of an uncultivated Bergeyella strain from the oral cavity to amniotic fluid in a case of preterm birth. J. Clin. Microbiol. 2006, 44, 1475–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.A.; Yoo, J.Y.; Kwon, E.J.; Kim, Y.J. Blood Microbial Communities During Pregnancy Are Associated With Preterm Birth. Front. Microbiol. 2019, 10, 1122. [Google Scholar] [CrossRef]
- Horgan, R.; Clancy, O.; Myers, J.; Baker, P.N. Gynaecology. An overview of proteomic and metabolomic technologies and their application to pregnancy research. BJOG Int. J. Obstet. Gynaecol. 2009, 116, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabolomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Bastos Maia, S.; Rolland Souza, A.S.; Costa Caminha, M.F.; Lins da Silva, S.; Callou Cruz, R.S.B.L.; Carvalho Dos Santos, C.; Batista Filho, M. Vitamin A and Pregnancy: A Narrative Review. Nutrients 2019, 11, 681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mawson, A.R. A Role for the Liver in Parturition and Preterm Birth. J. Transl. Sci. 2016, 2, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Berggren Söderlund, M.; Fex, G.A.; Nilsson-Ehle, P. Concentrations of retinoids in early pregnancy and in newborns and their mothers. Am. J. Clin. Nutr. 2005, 81, 633–636. [Google Scholar] [CrossRef]
- Quadro, L.; Hamberger, L.; Gottesman, M.E.; Colantuoni, V.; Ramakrishnan, R.; Blaner, W.S. Transplacental delivery of retinoid: The role of retinol-binding protein and lipoprotein retinyl ester. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E844–E851. [Google Scholar] [CrossRef]
- Suhre, K.; Schmitt-Kopplin, P. MassTRIX: Mass translator into pathways. Nucleic Acids Res. 2008, 36, W481–W484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- McParland, P.C.; Taylor, D.J. Recent Advances in Obstetrics and Gynaecology; Bonnar, J., Dunlop, W., Eds.; Royal Society of Medicine Press: London, UK, 2005; Volume 23, Chapter 3; p. 27. [Google Scholar]
- Menon, R. Spontaneous preterm birth, a clinical dilemma: Etiologic, pathophysiologic and genetic heterogeneities and racial disparity. Acta Obstet. Gynecol. Scand. 2008, 87, 590–600. [Google Scholar] [CrossRef]
- Lizewska, B.; Teul, J.; Kuc, P.; Lemancewicz, A.; Charkiewicz, K.; Goscik, J.; Kacerovsky, M.; Menon, R.; Miltyk, W.; Laudanski, P. Maternal Plasma Metabolomic Profiles in Spontaneous Preterm Birth: Preliminary Results. Mediat. Inflamm. 2018, 2018, 9362820. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.Y.; Hyeon, D.Y.; Shin, Y.; Kim, S.M.; You, Y.A.; Kim, D.; Hwang, D.; Kim, Y.J. Integrative analysis of transcriptomic data for identification of T-cell activation-related mRNA signatures indicative of preterm birth. Sci. Rep. 2021, 11, 2392. [Google Scholar] [CrossRef] [PubMed]
- Grizotte-Lake, M.; Zhong, G.; Duncan, K.; Kirkwood, J.; Iyer, N.; Smolenski, I.; Isoherranen, N.; Vaishnava, S. Commensals Suppress Intestinal Epithelial Cell Retinoic Acid Synthesis to Regulate Interleukin-22 Activity and Prevent Microbial Dysbiosis. Immunity 2018, 49, 1103–1115.e6. [Google Scholar] [CrossRef] [Green Version]
- Iyer, N.; Vaishnava, S. Vitamin A at the interface of host-commensal-pathogen interactions. PLoS Pathog. 2019, 15, e1007750. [Google Scholar] [CrossRef] [Green Version]
- Mei, Z.; Li, H.; Serdula, M.K.; Flores-Ayala, R.C.; Wang, L.; Liu, J.M.; Grummer-Strawn, L.M. C-reactive protein increases with gestational age during pregnancy among Chinese women. Am. J. Hum. Biol. 2016, 28, 574–579. [Google Scholar] [CrossRef]
- Miller, E.M. Changes in serum immunity during pregnancy. Am. J. Hum. Biol. 2009, 21, 401–403. [Google Scholar] [CrossRef] [Green Version]
- Sapin, V.; Begue, R.J.; Dastugue, B.; Chambon, P.; Dolle, P. Retinoids and mouse placentation. Trophobl. Res. 1998, 12, 57–76. [Google Scholar] [CrossRef]
- Soprano, D.R.; Blaner, W.S. Plasma retinol-binding protein. In The Retinoids, Biology, Chemistry and Medicine; Sporn, M.B., Roberts, A.B., Goodman, D.S., Eds.; Raven: New York, NY, USA, 1994; pp. 257–282. [Google Scholar]
- Horwitz, J.; Heller, J. Interactions of all-trans, 9-, 11-, and 13-cis-retinal, all-trans-retinyl acetate, and retinoic acid with human retinol-binding protein and prealbumin. J. Biol. Chem. 1973, 248, 6317–6324. [Google Scholar] [CrossRef]
- Kawagushi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of Vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Blanchon, L.; Marceau, G.; Borel, V.; Prat, C.; Herbet, A.; Bouvier, D.; Gallot, D.; Sapin, V. Implications of retinoid pathway in human fetal membranes: Study of target genes. Gynecol. Obstet. Fertil. 2011, 39, 370–372. [Google Scholar] [CrossRef]
- Dey, T.; Mukherjee, A.; Chakraborty, S. A practical overview of case-control studies in clinical practice. Chest 2020, 158, S57–S64. [Google Scholar] [CrossRef] [PubMed]
- Barupal, D.K.; Haldiya, P.K.; Wohlgemuth, G.; Kind, T.; Kothari, S.L.; Pinkerton, K.E.; Fiehn, O. MetaMapp: Mapping and visualizing metabolomic data by integrating information from biochemical pathways and chemical and mass spectral similarity. BMC Bioinform. 2012, 13, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Term Birth (n = 39) | Preterm Birth (n = 20) | p-Value |
|---|---|---|---|
| Maternal age | 33.3 ± 0.7 | 32.7 ± 1.0 | 0.626 |
| GAS | 23.0 ± 0.8 | 27.6 ± 1.6 | 0.010 * |
| preBMI | 21.0 ± 0.4 | 21.7 ± 0.7 | 0.397 |
| Parity | 0.488 | ||
| Nulliparous | 16 (42.1) | 8 (40.0) | |
| Multiparous | 21 (57.9) | 12 (60.0) | |
| Gravidity | 0.416 | ||
| 0 | 25 (64.1) | 11 (55.0) | |
| ≥1 | 14 (35.9) | 9 (45.0) | |
| WBC (×103/mL) | 8.8 ± 0.4 | 11.6 ± 0.6 | <0.001 * |
| C-reactive protein (mg/dL) | 0.3 ± 0.1 | 0.8 ± 0.2 | 0.034 * |
| pregBMI | 26.8 ± 0.6 | 26.4 ± 0.8 | 0.397 |
| GAD | 39.0 ± 0.2 | 33.3 ± 1.0 | <0.001 * |
| Delivery mode | 0.026 † | ||
| Normal delivery | 23 (59.0) | 5 (25.0) | |
| Cesarean section | 16 (41.0) | 15 (75.0) | |
| Birth weight (Kg) | 3.2 ± 0.1 | 2.2 ± 0.2 | <0.001 * |
| Gender, n (%) | 0.651 | ||
| Male | 25 (64.1) | 14 (70.0) | |
| Female | 14 (35.9) | 6 (30.0) | |
| APGAR 1 min | 9.7 ± 0.1 | 7.9 ± 0.5 | 0.002 * |
| APGAR 5 min | 9.9 ± 0.1 | 8.9 ± 0.4 | 0.009 * |
| Term Birth (n = 24, mg/L) | Preterm Birth (n = 16, mg/L) | p-Value |
|---|---|---|
| 63.4 ± 4.2 | 122.9 ± 16.7 * | 0.012 |
| Metabolite | AUC | p-Value | SENS | SPEC | PPV | NPV | Accuracy |
|---|---|---|---|---|---|---|---|
| At-retinal | 0.808 | <0.001 | 75.0% | 84.2% | 68.2% | 86.5% | 79.7% |
| 13cis-RA | 0.826 | <0.001 | 85.0% | 68.4% | 58.6% | 90.0% | 74.6% |
| 9cis-RA | 0.679 | 0.026 | 45.0% | 92.1% | 69.2% | 76.1% | 74.6% |
| Retinyl palmitate | 0.670 | 0.035 | 40.0% | 94.7% | 72.7% | 75.0% | 74.6% |
| RBP | 0.736 | 0.012 | 62.5% | 95.8% | 90.9% | 79.3% | 82.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, Y.-A.; Hwang, S.-Y.; Kim, S.-M.; Park, S.; Lee, G.-I.; Park, S.; Ansari, A.; Lee, J.; Kwon, Y.; Kim, Y.-J. Identification of Indicators for Preterm Birth Using Retinoid Metabolites. Metabolites 2021, 11, 443. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11070443
You Y-A, Hwang S-Y, Kim S-M, Park S, Lee G-I, Park S, Ansari A, Lee J, Kwon Y, Kim Y-J. Identification of Indicators for Preterm Birth Using Retinoid Metabolites. Metabolites. 2021; 11(7):443. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11070443
Chicago/Turabian StyleYou, Young-Ah, Soo-Yeon Hwang, Soo-Min Kim, Seojeong Park, Ga-In Lee, Sunwha Park, AbuZar Ansari, Jeongae Lee, Youngjoo Kwon, and Young-Ju Kim. 2021. "Identification of Indicators for Preterm Birth Using Retinoid Metabolites" Metabolites 11, no. 7: 443. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11070443