
Adipose Tissue and Biological Factors. Possible Link between Lymphatic System Dysfunction and Obesity


Abstract
:1. Adipose Tissue—Structure and Functions
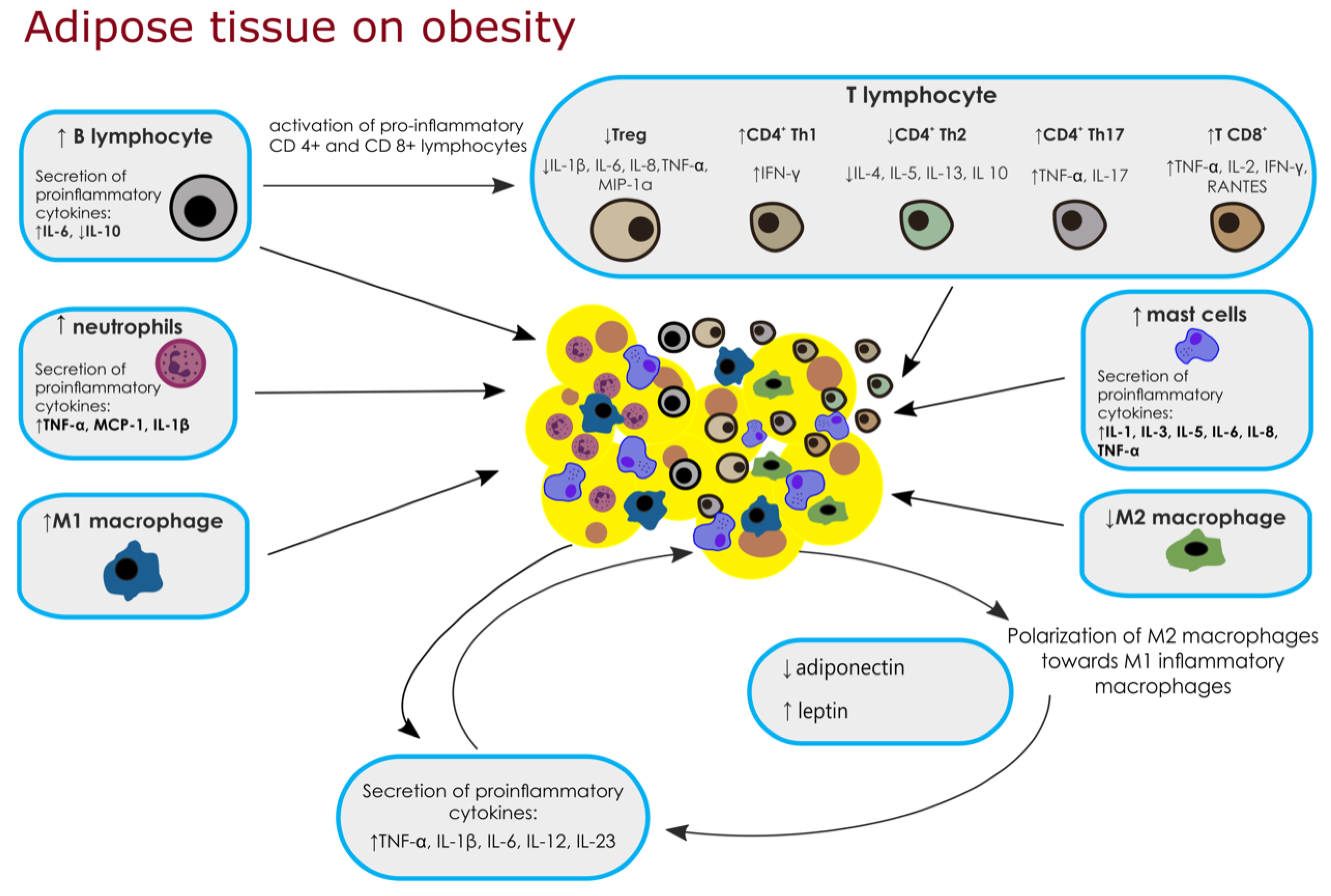
2. Adipose Tissue Immune Cells and Biological Factors
2.1. Macrophages
2.2. Neutrophils
2.3. Mast Cells
2.4. T Lymphocytes
2.4.1. Regulatory T Cells
2.4.2. CD4+ T Cells
2.4.3. Cytotoxic CD8+ T Cells
2.5. B Lymphocytes
3. Saturated Fatty Acids
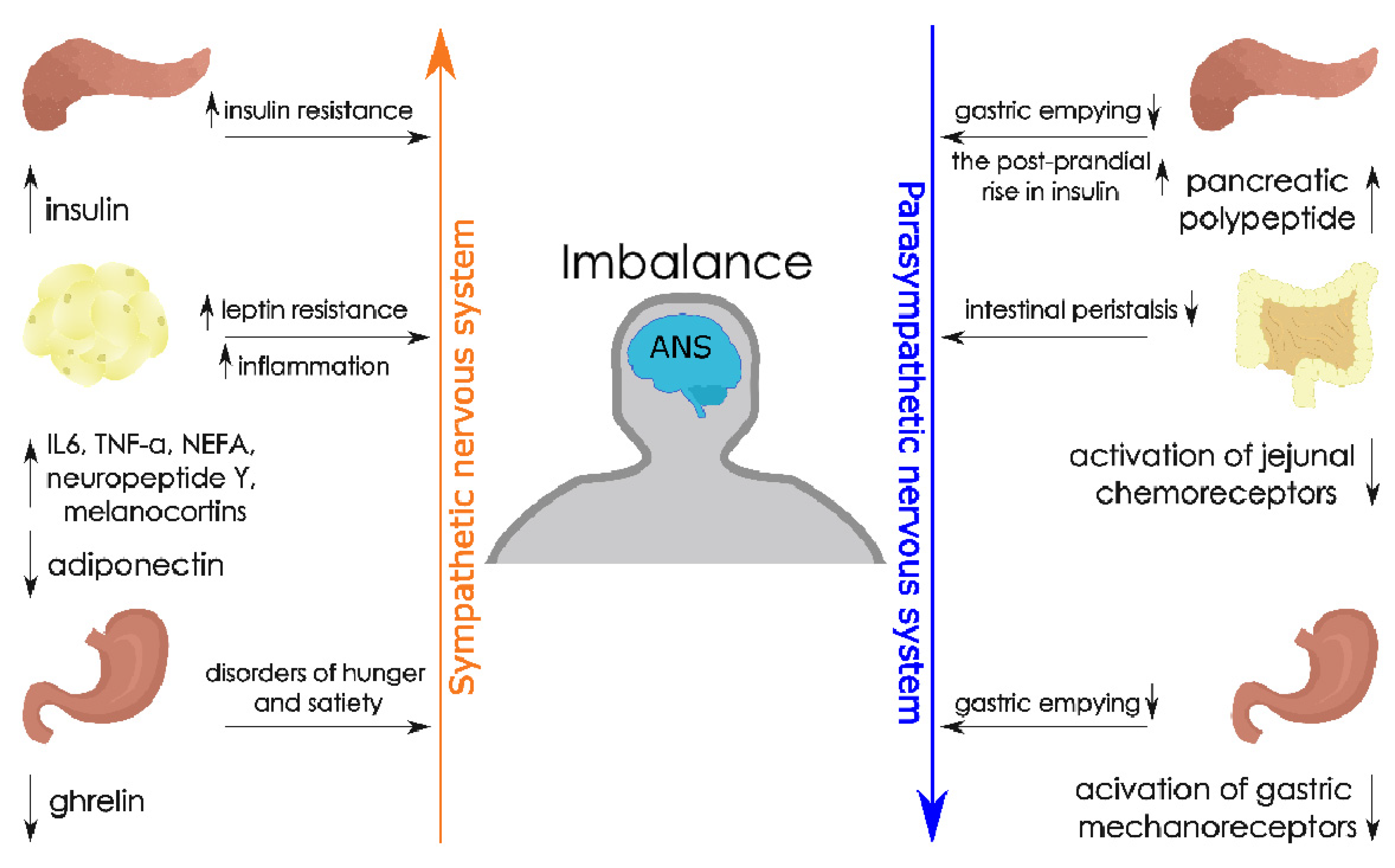
4. Adipose Tissue, Obesity and the Autonomic Nervous System
5. Lymphatic System—Physiology and Functions
5.1. Lymphatic System—Fat Absorption and Cholesterol Transport
5.2. Lymphatic System—Role in Regulating the Immune Response
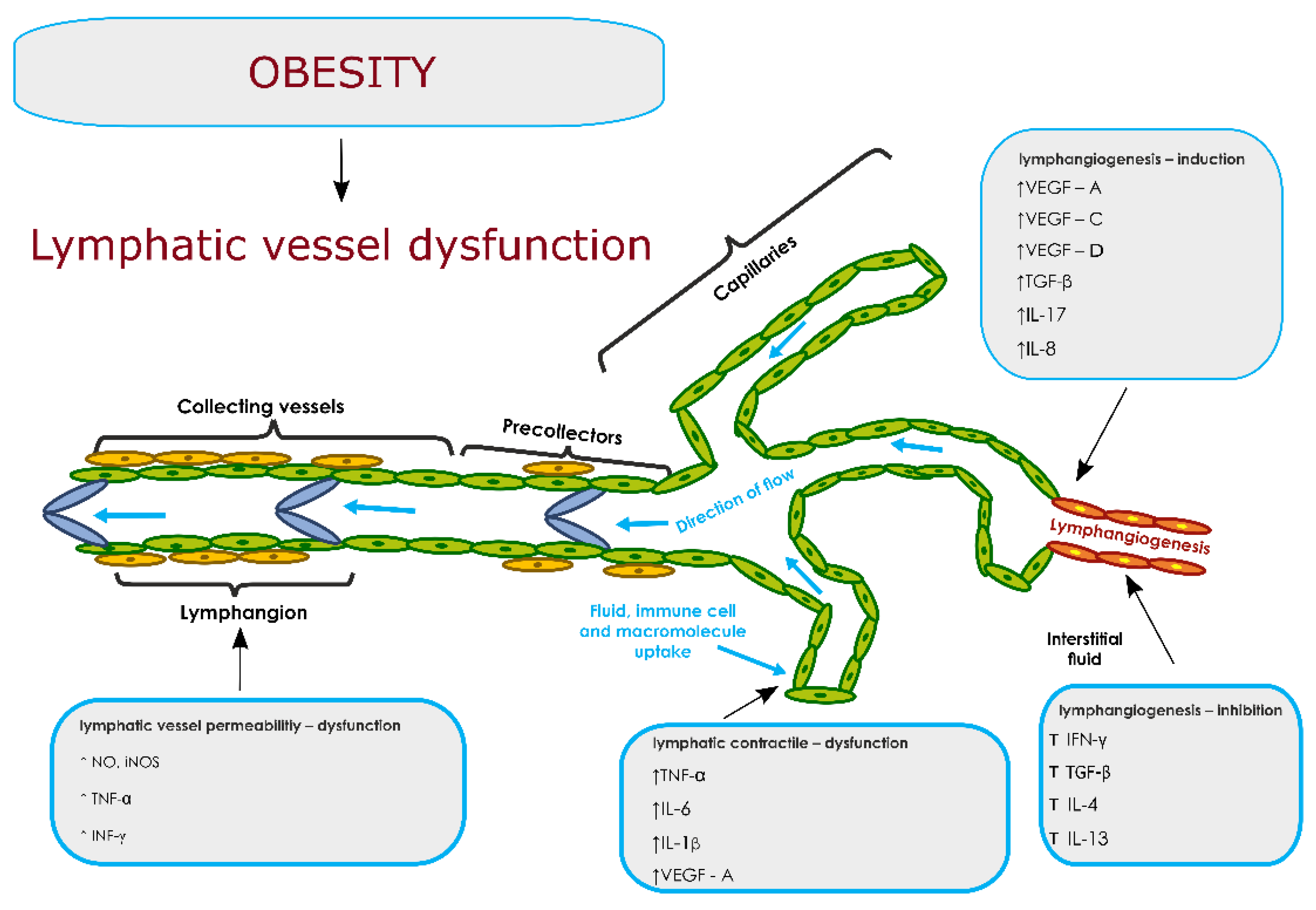
6. The Lymphatic System and Obesity
7. Dysfunction of the Lymphatic System: Pro- and Anti-Lymphangiogenic Factors in Obesity
7.1. Lymphatic System Dysfunctions—Permeability of Lymphatic Vessels
7.2. Lymphatic System Dysfunctions—Lymphatic Contractility
8. Manual Lymphatic Drainage as a New Therapeutic Option Supporting the Functions of the Lymphatic System in the Course of Obesity
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| WHO | World Health Organization |
| ANS | autonomic nervous system |
| BALT | Bronchus-associated lymphoid tissue |
| BAT | Brown adipose tissue |
| BMI | body mass index |
| CGRP | calcitonin gene-related peptide |
| DCs | dendritic cells |
| EEG | electroencephalogram |
| GALT | Gut-associated lymphoid tissue |
| gp38 | podoplanin |
| HRV | heart rate variability |
| IFN-γ | Interferon-γ |
| IL-1 | Interleukin-1 |
| IL-10 | Interleukin-10 |
| IL-12 | Interleukin-12 |
| IL-13 | Interleukin-13 |
| IL-17 | Interleukin-17 |
| IL-1β | Interleukin-1β |
| IL-2 | Interleukin-2 |
| IL-23 | Interleukin-23 |
| IL-3 | Interleukin-3 |
| IL-4 | Interleukin-4 |
| IL-5 | Interleukin-5 |
| IL-8 | Interleukin-8 |
| iNOS | induced nitric oxide synthase |
| IR | insulin resistance |
| JNK | c-Jun N-terminal kinases |
| LEC | lymphatic endothelial cell |
| LYVE-1 | Lymphatic Vessel Endothelial Receptor 1 |
| MALT | mucosa-associated lymphoid tissue |
| MIP-1α | macrophage inflammatory protein-1α |
| MLD | manual lymphatic drainage |
| NA-SR | noradrenaline spillover rate |
| NEFA | Non-esterified free fatty acids |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK | natural killers |
| NO | nitric oxide |
| PAI-1 | plasminogen activator inhibitor-1 |
| PNS | parasympathetic nervous system |
| Prox-1 | Prospero Homeobox 1 |
| RBP-4 | retinol-binding protein |
| SNA | sympathetic nerve activity |
| SNS | sympathetic nervous system |
| SP | substance P |
| SVF | Stromal vascular fraction |
| T2DM | Type 2 diabetes |
| TGF-β | Transforming growth factor-β |
| Th1 | T-helper-type 1 |
| Th17 | T-helper-type 17 |
| Th2 | T-helper-type 2 |
| TNF-α | Tumor necrosis factor-α |
| IL-6 | Interleukin-6 |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| Treg | regulatory T cells |
| VAT | Visceral adipose tissue |
| VEGF-A | vascular endothelial growth factor A |
| VEGF-C | vascular endothelial growth factor C |
| VEGF-D | vascular endothelial growth factor D |
| VEGFR-3 | vascular endothelial growth factor receptor 3 |
| VIP | vasoactive intestinal peptide |
| WAT | White adipose tissue |
References
- Bray, G.; Kim, K.; Wilding, J.P.H. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Barrera, J.; Sainz-Escárrega, V.H.; Medina-Urritia, A.X.; Jorge-Galarza, E.; Osorio-Alonso, H.; Torres-Tamayo, M. Dysfunctional adiposity index as a marker of adipose tissue morpho-functional abnormalities and metabolic disorders in apparently healthy subjects. Adipocyte 2021, 10, 142–152. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S. White, brown, beige and pink: A rainbow in the adipose organ. Curr. Opin. Endocr. Metab. Res. 2019, 4, 29–36. [Google Scholar] [CrossRef]
- Weschenfelder, C.; Schaan de Quadros, A.; Lorenzon Dos Santos, J.; Bueno Garofallo, S.; Marcadenti, A. Adipokines and Adipose Tissue-Related Metabolites, Nuts and Cardiovascular Disease. Metabolites 2020, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froy, O.; Garaulet, M. The Circadian Clock in White and Brown Adipose Tissue: Mechanistic, Endocrine, and Clinical Aspects. Endocr. Rev. 2018, 39, 261–273. [Google Scholar] [CrossRef]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat. Med. 2011, 17, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K.; Jachimowicz-Duda, O.; Wąż, P. The cut-off value for interleukin 34 as an additional potential inflammatory biomarker for the prediction of the risk of diabetic complications. Biomarkers 2016, 21, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Corbi, G.; Polito, R.; Monaco, M.L.; Cacciatore, F.; Scioli, M.; Ferrara, N.; Daniele, A.; Nigro, E. Adiponectin Expression and Genotypes in Italian People with Severe Obesity Undergone a Hypocaloric Diet and Physical Exercise Program. Nutrients 2019, 11, 2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, L.; Nowak, M.; Varghese, B.; Clark, J.; Hogan, A.E.; Toxavidis, V.; Balk, S.P. Adipose tissue invariant NKT cells protect against diet-induced obesity and metabolic disorder through regulatory cytokine production. Immunity 2012, 37, 574–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-Induced Changes in Adipose Tissue Microenvironment and Their Impact on Cardiovascular Disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.; et al. Evidence that TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab. 2018, 27, 1096–1110.e5. [Google Scholar] [CrossRef] [Green Version]
- Kanter, J.E.; Kramer, F.; Barnhart, S.; Averill, M.M.; Vivekanandan-Giri, A.; Vickery, T.; Li, L.O.; Becker, L.; Yuan, W.; Chait, A.; et al. Diabetes promotes an inflammatory macrophage phenotype and atherosclerosis through acyl-CoA synthetase 1. Proc. Natl. Acad. Sci. USA 2012, 109, 715–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, R.; Hilhorst, M.; Zhang, H.; Zeisbrich, M.; Berry, G.J.; Wallis, B.B.; Harrison, D.G.; Giacomini, J.C.; Goronzy, J.J.; Weyand, C.M. Glucose metabolism controls disease-specific signatures of macrophage effector functions. JCI Insight 2018, 3, e123047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.E.; Ko, M.S.; Yun, J.Y.; Kim, M.O.; Kim, J.H.; Park, H.S.; Kim, A.R.; Kim, H.J.; Kim, B.J.; Ahn, Y.E.; et al. Nitric Oxide Produced by Macrophages Inhibits Adipocyte Differentiation and Promotes Profibrogenic Responses in Preadipocytes to Induce Adipose Tissue Fibrosis. Diabetes 2016, 65, 2516–2528. [Google Scholar] [CrossRef] [Green Version]
- Nam, H.; Ferguson, B.S.; Stephens, J.M.; Morrison, R.F. Impact of obesity on IL-12 family gene expression in insulin responsive tissues. Biochim. Biophys. Acta 2012, 1832, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, V.T.; Shulman, G.I. Mechanisms for IR: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [Green Version]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, S.; Oh, D.Y.; Bandyopadhyay, G.; Li, D.; Xu, J.; McNelis, J.; Lu, M.; Li, P.; Yan, Q.; Zhu, Y.; et al. Neutrophils mediate insulin resistance in mice fed a high-fat diet through secreted elastase. Nat. Med. 2012, 18, 1407–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgazar-Carmon, V.; Rudich, A.; Hadad, N.; Levy, R. Neutrophils transiently infiltrate intra-abdominal fat early in the course of high-fat feeding. J. Lipid Res. 2008, 49, 1894–1903. [Google Scholar] [CrossRef] [Green Version]
- Dam, V.; Sikder, T.; Santosa, S. From neutrophils to macrophages: Differences in regional AT depots. Obes. Rev. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Liu, J.; Divoux, A.; Sun, J.; Zhang, J.; Clément, K.; Glickman, J.N.; Sukhova, G.K.; Wolters, P.J.; Du, J.; Gorgun, C.Z.; et al. Genetic deficiency and pharmacological stabilization of mast cells reduce diet-induced obesity and diabetes in mice. Nat. Med. 2009, 15, 940–945. [Google Scholar] [CrossRef] [Green Version]
- Divoux, A.; Moutel, S.; Poitou, C.; Lacasa, D.; Veyrie, N.; Aissat, A.; Arock, M.; Guerre-Millo, M.; Clément, K. Mast cells in human adipose tissue: Link with morbid obesity, inflammatory status, and diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E1677–E1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, S.; Kumari, R.; Prashar, Y.; Gill, N.S. Review of various molecular targets on mast cells and its relation to obesity: A future perspective. Diabetes Metab. Syndr. 2017, 11, 1001–1007. [Google Scholar] [CrossRef]
- Anaya, J.M.; Shoenfeld, Y.; Rojas-Villarraga, A.; Levy, R.A.; Cervera, R. Introduction to T and B lymphocytes. In Autoimmunity: From Bench to Bedside; El Rosario University Press: Bogota, Colombia, 2013. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK459471/ (accessed on 8 September 2021).
- Tiemessen, M.M.; Jagger, A.L.; Evans, H.G.; van Herwijnen, M.J.; John, S.; Taams, L.S. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 19446–19451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deiuliis, J.; Shah, Z.; Shah, N.; Needleman, B.; Mikami, D.; Narula, V.; Perry, K.; Hazey, J.; Kampfrath, T.; Kollengode, M.; et al. Visceral adipose inflammation in obesity is associated with critical alterations in tregulatory cell numbers. PLoS ONE 2011, 6, e16376. [Google Scholar] [CrossRef] [Green Version]
- Van der Zalm, I.; van der Valk, E.S.; Wester, V.L.; Nagtzaam, N.; van Rossum, E.; Leenen, P.; Dik, W.A. Obesity-associated T-cell and macrophage activation improve partly after a lifestyle intervention. Int. J. Obes. 2020, 44, 1838–1850. [Google Scholar] [CrossRef] [PubMed]
- Surendar, J.; Frohberger, S.J.; Karunakaran, I.; Schmitt, V.; Stamminger, W.; Neumann, A.L. Adiponectin Limits IFN-γ and IL-17 Producing CD4 T Cells in Obesity by Restraining Cell Intrinsic Glycolysis. Front. Immunol. 2019, 10, 2555. [Google Scholar] [CrossRef]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef]
- Misumi, I.; Starmer, J.; Uchimura, T.; Beck, M.A.; Magnuson, T.; Whitmire, J.K. Obesity Expands a Distinct Population of T Cells in Adipose Tissue and Increases Vulnerability to Infection. Cell Rep. 2019, 27, 514–524.e5. [Google Scholar] [CrossRef] [Green Version]
- Inzaugarat, M.E.; Billordo, L.A.; Vodánovich, F.; Cervini, G.M.; Casavalle, P.L.; Vedire, C.; Cherñavsky, A.C. Alterations in innate and adaptive immune leukocytes are involved in paediatric obesity. Pediatr. Obes. 2014, 9, 381–390. [Google Scholar] [CrossRef]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef]
- Kim, M.S.; Choi, M.S.; Han, S.N. High fat diet-induced obesity leads to proinflammatory response associated with higher expression of NOD2 protein. Nutr. Res. Pract. 2011, 5, 219–223. [Google Scholar] [CrossRef] [PubMed]
- De Furia, J.; Belkina, A.C.; Jagannathan-Bogdan, M.; Snyder-Cappione, J.; Carr, J.D.; Nersesova, Y.R.; Markham, D.; Strissel, K.J.; Watkins, A.A.; Zhu, M.; et al. B cells promote inflammation in obesity and type 2 diabetes through regulation of T-cell function and an inflammatory cytokine profile. Proc. Natl. Acad. Sci. USA 2013, 110, 5133–5138. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Ferracci, F.; Diaz, A.; Romero, M.; Lechner, S.; Blomberg, B. Obesity decreases B cell responses in young and elderly individuals. Obesity 2016, 24, 615–625. [Google Scholar] [CrossRef]
- Khaw, K.T.; Friesen, M.D.; Riboli, E.; Luben, R.; Wareham, N. Plasma Phospholipid Fatty Acid Concentration and Incident Coronary Heart Disease in Men and Women: The EPIC-Norfolk Prospective Study. PLoS Med. 2012, 9, e1001255. [Google Scholar] [CrossRef] [Green Version]
- Kurotani, K.; Sato, M.; Yasuda, K.; Kashima, K.; Tanaka, S.; Hayashi, T.; Shirouchi, B.; Akter, S.; Kashino, I.; Hayabuchi, H.; et al. Even- and odd-chain saturated fatty acids in serum phospholipids are differentially associated with adipokines. PLoS ONE 2017, 12, e178192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.L.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: Implications for the pathogenesis of obesity. J. Neurosci. 2009, 29, 359–370. [Google Scholar] [CrossRef]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Magnuson, A.M.; Regan, D.P.; Fouts, J.K.; Booth, A.D.; Dow, S.W.; Foster, M.T. Diet-induced obesity causes visceral, but not subcutaneous, lymph node hyperplasia via increases in specific immune cell populations. Cell Prolif. 2017, 50, e12365. [Google Scholar] [CrossRef] [Green Version]
- Heilbronn, L.; Liu, B. Do adipose tissue macrophages promote insulin resistance or adipose tissue remodeling in humans? Horm. Mol. Biol. Clin. Investig. 2014, 20, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pongratz, G.; Straub, R.H. The sympathetic nervous response in inflammation. Arthritis Res. Ther. 2014, 16, 504. [Google Scholar] [CrossRef] [Green Version]
- Yun, A.J.; Lee, P.Y.; Bazar, K.A. Autonomic dysregulation as a basis of cardiovascular, endocrine, and inflammatory disturbances associated with obstructive sleep apnea and other conditions of chronic hypoxia, hypercapnia, and acidosis. Med. Hypotheses 2004, 62, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Guarino, D.; Nannipieri, M.; Iervasi, G.; Taddei, S.; Bruno, R.M. The Role of the Autonomic Nervous System in the Pathophysiology of Obesity. Front. Physiol. 2017, 8, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, S.B.; Gsponer, D.; Montoya-Zegarra, J.A.; Schneider, M.; Scholkmann, F.; Tacconi, C.; Noerrelykke, S.F.; Proulx, S.T.; Detmar, M. A Distinct Role of the Autonomic Nervous System in Modulating the Function of Lymphatic Vessels under Physiological and Tumor-Draining Conditions. Cell Rep. 2019, 27, 3305–3314.e13. [Google Scholar] [CrossRef]
- Costa, J.; Moreira, A.; Moreira, P.; Delgado, L.; Silva, D. Effects of weight changes in the autonomic nervous system: A systematic review and meta-analysis. Clin. Nutr. 2019, 38, 110–126. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Boesveldt, S.; Delplanque, S.; de Graaf, C.; de Wijk, R.A. Sensory-specific satiety: Added insights from autonomic nervous system responses and facial expressions. Physiol. Behav. 2017, 170, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Masi, E.B.; Valdés-Ferrer, S.I.; Steinberg, B.E. The vagusneurometabolic interface and clinical disease. Int. J. Obes. 2018, 42, 1101–1111. [Google Scholar] [CrossRef]
- Dockray, G.J. Gastrointestinal hormones and the dialogue between gut and brain. J. Physiol. 2014, 592, 2927–2941. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.; Gupta, V. Effect of obesity on autonomic nervous system. Int. J. Cur. Bio. Med. Sci. 2011, 1, 15–18. [Google Scholar]
- Monda, V.; Valenzano, A.; Monda, M. Modifications of Activity of Autonomic Nervous System, and Resting Energy Expenditure in Women Using Hormone-Replacement Therapy. Biol. Med. 2016, 8, 1. [Google Scholar] [CrossRef]
- Straznicky, N.E.; Lambert, G.W.; Lambert, E.A. Neuroadrenergic dysfunction in obesity: An overview of the effects of weight loss. Curr. Opin. Lipidol. 2010, 21, 21–30. [Google Scholar] [CrossRef]
- Nogueiras, R.; Pérez-Tilve, D.; Veyrat-Durebex, C.; Morgan, D.A.; Varela, L.; Haynes, W.G. Direct control of peripheral lipid deposition by CNS GLP-1 receptor signaling is mediated by the sympathetic nervous system and blunted in diet-induced obesity. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 5916–5925. [Google Scholar] [CrossRef]
- Smith, M.M.; Minson, C.T. Obesity and adipokines: Effects on sympathetic overactivity. J. Physiol. 2012, 590, 1787–1801. [Google Scholar] [CrossRef]
- Landsberg, L. Insulin-mediated sympathetic stimulation: Role in the pathogenesis of obesity-related hypertension (or, how insulin affects blood pressure, and why). J. Hypertens. 2001, 19, 523–528. [Google Scholar] [CrossRef]
- Kalil, G.Z.; Haynes, W.G. Sympathetic nervous system in obesity-related hypertension: Mechanisms and clinical implications. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2012, 35, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Van der Valk, E.S.; Savas, M.; van Rossum, E. Stress and Obesity: Are There More Susceptible Individuals? Curr. Obes. Rep. 2018, 7, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, A.A.; do Carmo, J.; Dubinion, J.; Hall, J.E. The role of the sympathetic nervous system in obesity-related hypertension. Curr. Hypertens. Rep. 2009, 11, 206–211. [Google Scholar] [CrossRef]
- Indumathy, J.; Pal, G.K.; Pal, P.; Ananthanarayanan, P.H.; Parija, S.C.; Balachander, J.; Dutta, T.K. Association of sympathovagal imbalance with obesity indices, and abnormal metabolic biomarkers and cardiovascular parameters. Obes. Res. Clin. Pract. 2015, 9, 55–66. [Google Scholar] [CrossRef]
- Huggett, R.J.; Burns, J.; Mackintosh, A.F.; Mary, D.A. Sympathetic neural activation in nondiabetic metabolic syndrome and its further augmentation by hypertension. Hypertension 2004, 44, 847–852. [Google Scholar] [CrossRef] [Green Version]
- Villarroya, F.; Vidal-Puig, A. Beyond the sympathetic tone: The new brown fat activators. Cell Metab. 2013, 17, 638–643. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy metabolism in the liver. Comprehens. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W. Sympathetic neuro-adipose connections mediate leptin-driven lipolysis. Cell 2015, 163, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Babic, T.; Travagli, R.A. Neural Control of the Pancreas. Pancreapedia Exocrine Pancreas Knowl. Base 2016. [Google Scholar] [CrossRef]
- Lambert, E.A.; Straznicky, N.E.; Dixon, J.B.; Lambert, G.W. Should the sympathetic nervous system be a target to improve cardiometabolic risk in obesity? Am. J. Physiol. Heart Circ. Physiol. 2015, 309, 244–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.E.; da Silva, A.A.; do Carmo, J.; Dubinion, J.; Hamza, S.; Munusamy, S.; Smith, G.; Stec, D.E. Obesity-induced hypertension: Role of sympathetic nervous system, leptin, and melanocortins. J. Biol. Chem. 2010, 285, 17271–17276. [Google Scholar] [CrossRef] [Green Version]
- Breit, S.; Kupferberg, A.; Rogler, G.; Hasler, G. Vagus Nerve as Modulator of the Brain-Gut Axis in Psychiatric and Inflammatory Disorders. Front. Psychiatry 2018, 9, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, K.N.; Verheijden, S.; Boeckxstaens, G.E. The Vagus Nerve in Appetite Regulation, Mood, and Intestinal Inflammation. Gastroenterology 2017, 152, 730–744. [Google Scholar] [CrossRef] [Green Version]
- German, J.; Kim, F.; Schwartz, G.J.; Havel, P.J.; Rhodes, C.J.; Schwartz, M.W.; Morton, G.J. Hypothalamic leptin signaling regulates hepatic insulin sensitivity via a neurocircuit involving the vagus nerve. Endocrinology 2009, 150, 4502–4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanida, M.; Yamamoto, N.; Morgan, D.A.; Kurata, Y.; Shibamoto, T.; Rahmouni, K. Leptin receptor signaling in the hypothalamus regulates hepatic autonomic nerve activity via phosphatidylinositol 3-kinase and AMP-activated protein kinase. J. Neurosci. 2015, 35, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikramuddin, S.; Blackstone, R.P.; Brancatisano, A.; Toouli, J.; Shah, S.N.; Wolfe, B.M.; Fujioka, K.; Maher, J.W.; Swain, J.; Que, F.G.; et al. Effect of reversible intermittent intra-abdominal vagal nerve blockade on morbid obesity: The ReCharge randomized clinical trial. JAMA 2014, 312, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Babic, T.; Browning, K.N. The role of vagal neurocircuits in the regulation of nausea and vomiting. Eur. J. Pharm. 2014, 722, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triggiani, A.I.; Valenzano, A.; Ciliberti, M.A.; Moscatelli, F.; Villani, S.; Monda, M.; Messina, G.; Federici, A.; Babiloni, C.; Cibelli, G. Heart rate variability is reduced in underweight and overweight healthy adult women. Clin. Physiol. Funct. Imaging 2017, 37, 162–167. [Google Scholar] [CrossRef]
- Mignini, F.; Sabbatini, M.; Coppola, L.; Cavallotti, C. Analysis of nerve supply pattern in human lymphatic vessels of young and old men. Lymphat. Res. Biol. 2012, 10, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Cleypool, C.; Mackaaij, C.; Lotgerink Bruinenberg, D.; Schurink, B.; Bleys, R. Sympathetic nerve distribution in human lymph nodes. J. Anat. 2021, 239, 282–289. [Google Scholar] [CrossRef]
- Hsu, M.C.; Itkin, M. Lymphatic Anatomy. Tech. Vasc. Interv. Radiol. 2016, 19, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Lee, S.; Hong, Y.K. The new era of the lymphatic system: No longer secondary to the blood vascular system. Cold Spring Harb. Perspect. Med. 2012, 2, a006445. [Google Scholar] [CrossRef] [Green Version]
- Ruddle, N.H.; Akirav, E.M. Secondary lymphoid organs: Responding to genetic and environmental cues in ontogeny and the immune response. J. Immunol. 2009, 183, 2205–2212. [Google Scholar] [CrossRef]
- Schulte-Merker, S.; Sabine, A.; Petrova, T.V. Lymphatic vascular morphogenesis in development, physiology, and disease. J. Cell Biol. 2011, 16, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Ratajska, A.; Jankowska-Steifer, E.; Czarnowska, E.; Flaht, A.; Radomska-Leśniewska, D. Morphogenesis, structure and properties of lymphatic vessels. Postep. Hig. Med. Dosw. 2012, 19, 901–912. [Google Scholar] [CrossRef]
- Albrecht, I.; Christofori, G. Molecular mechanisms of lymphangiogenesis in development and cancer. Int. J. Dev. Biol. 2011, 55, 483–494. [Google Scholar] [CrossRef]
- Danussi, C.; Spessotto, P.; Petrucco, A.; Wassermann, B.; Sabatelli, P.; Montesi, M.; Doliana, R.; Bressan, G.M.; Colombatti, A. Emilin1 deficiency causes structural and functional defects of lymphatic vasculature. Mol. Cell. Biol. 2008, 28, 4026–4039. [Google Scholar] [CrossRef] [Green Version]
- Alders, M.; Hogan, B.M.; Gjini, E.; Salehi, F.; Al-Gazali, L.; Hennekam, E.A.; Holmberg, E.E.; Mannens, M.M.; Mulder, M.F. Mutations in CCBE1 cause generalized lymph vessel dysplasia in humans. Nat. Genet. 2009, 41, 1272–1274. [Google Scholar] [CrossRef]
- Jiang, X.; Nicolls, M.R.; Tian, W.; Rockson, S.G. Lymphatic Dysfunction, Leukotrienes, and Lymphedema. Ann. Rev. Physiol. 2018, 10, 49–70. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.C.; Baluk, P.; Srinivasan, R.S.; Oliver, G.; McDonald, D.M. Plasticity of button-like junctions in the endothelium of airway lymphatics in development and inflammation. Am. J. Pathol. 2012, 180, 2561–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieweg, O.E.; Tanis, P.J.; Leong, S.P.L. Anatomy and Physiology of Lymphatic Circulation: Application to Lymphatic Mapping. In Atlas of Lymphoscintigraphy and Sentinel Node Mapping; Mariani, G., Vidal-Sicart, S., Valdés Olmos, R., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Karkkainen, M.J.; Saaristo, A.; Jussila, L.; Karila, K.A.; Lawrence, E.C.; Pajusola, K.; Bueler, H.; Eichmann, A.; Kauppinen, R.; Kettunen, M.I.; et al. A model for gene therapy of human hereditary lymphedema. Proc. Natl. Acad. Sci. USA 2001, 98, 12677–12682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitti, M.D.; Hespe, G.E.; Kataru, R.P.; García Nores, G.D.; Savetsky, I.L.; Torrisi, J.S.; Gardenier, J.C.; Dannenberg, A.J.; Mehrara, B.J. Obesity-induced lymphatic dysfunction is reversible with weight loss. J. Physiol. 2016, 594, 7073–7087. [Google Scholar] [CrossRef] [PubMed]
- Gashev, A.A.; Zawieja, D.C. Hydrodynamic regulation of lymphatic transport and the impact of aging. Pathophysiol. Off. J. Int. Soc. Pathophysiol. 2010, 17, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, W.; Engeset, A.; Jaeger, P.M.; Sokolowski, J.; Theodorsen, L. Flow and composition of leg lymph in normal men during venous stasis, muscular activity and local hyperthermia. Actaphysiol. Scand. 1977, 99, 149–155. [Google Scholar] [CrossRef]
- Scallan, J.P.; Zawieja, S.D.; Castorena-Gonzalez, J.A.; Davis, M.J. Lymphatic pumping: Mechanics, mechanisms and malfunction. J. Physiol. 2016, 594, 5749–5768. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Davis, M.J.; Muthuchamy, M. Emerging trends in the pathophysiology of lymphatic contractile function. Semin. Cell Dev. Biol. 2015, 38, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriondo, A.; Solari, E.; Marcozzi, C.; Negrini, D. Lymph flow pattern in pleural diaphragmatic lymphatics during intrinsic and extrinsic isotonic contraction. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H60–H70. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, A.; Solari, E.; Marcozzi, C.; Negrini, D. Diaphragmatic lymphatic vessel behavior during local skeletal muscle contraction. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H193–H205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solari, E.; Marcozzi, C.; Negrini, D.; Moriondo, A. Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow. Biology 2020, 9, 463. [Google Scholar] [CrossRef]
- Clodius, L. Lymphoedema. In Plastic Surgery, Vol 6: The Trunk and Lower Extremity; McCarthy, J.G., Ed.; W.B. Sunders: Philadelphia, PA, USA, 1990; pp. 4093–4101. [Google Scholar]
- Greene, A.K. Obesity-induced lymphedema: Clinical and lymphoscintigraphic features. Plast. Reconstr. Surg. 2015, 6, 1715–1719. [Google Scholar] [CrossRef]
- Varaliová, Z.; Vlasák, R.; Čížková, T.; Gojda, J.; Potočková, J.; Šiklová, M.; Krauzová, E.; Štěpán, M.; Bülow, J.; Štich, V.; et al. Lymphatic drainage affects lipolytic activity of femoral adipose tissue in women. Int. J. Obes. 2020, 44, 1974–1978. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Oliver, G. Development of the mammalian lymphatic vasculature. J. Clin. Investig. 2014, 124, 888–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vranova, M.; Halin, C. Lymphatic Vessels in Inflammation. J. Clin. Cell Immunol. 2014, 5, 250. [Google Scholar] [CrossRef]
- Randolph, G.J.; Ivanov, S.; Zinselmeyer, B.H.; Scallan, J.P. The Lymphatic System: Integral Roles in Immunity. Ann. Rev. Immunol. 2017, 35, 31–52. [Google Scholar] [CrossRef] [Green Version]
- Cromer, W.; Wang, W.; Zawieja, S.D.; von der Weid, P.Y.; Newell-Rogers, M.K.; Zawieja, D.C. Colonic insult impairs lymph flow, increases cellular content of the lymph, alters local lymphatic microenvironment, and leads to sustained inflammation in the rat ileum. Inflamm. Bowel Dis. 2015, 21, 1553–1563. [Google Scholar] [CrossRef] [Green Version]
- Dixon, J.B. Lymphatic lipid transport: Sewer or subway? Trends Endocrinol. Metab. 2010, 21, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, C.; Li, W.; Fulp, B.; Platt, A.M.; Gautier, E.L.; Westerterp, M.; Bittman, R.; Tall, A.R.; Chen, S.H.; Thomas, M.J.; et al. Lymphatic vasculature mediates macrophage reverse cholesterol transport in mice. J. Clin. Investig. 2013, 123, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifarelli, V.; Eichmann, A. The Intestinal Lymphatic System: Functions and Metabolic Implications. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, D.W.; Herbst, K.L. Lipedema: A Relatively Common Disease with Extremely Common Misconceptions. Plastic and reconstructive surgery. Glob. Open 2016, 4, e1043. [Google Scholar] [CrossRef]
- Cooke, C.J.; Nanjee, M.N.; Stepanova, I.P.; Olszewski, W.L.; Miller, N.E. Variations in lipid and apolipoprotein concentrations in human leg lymph: Effects of posture and physical exercise. Atherosclerosis 2004, 173, 39–45. [Google Scholar] [CrossRef]
- Huang, L.H.; Elvington, A.; Randolph, G.J. The role of the lymphatic system in cholesterol transport. Front. Pharmacol. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuorio, T.; Nurmi, H.; Moulton, K.; Kurkipuro, J.; Robciuc, M.R.; Öhman, M.; Heinonen, S.E.; Samaranayake, H.; Heikura, T.; Alitalo, K. Lymphatic vessel insufficiency in hypercholesterolemic mice alters lipoprotein levels and promotes atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1162–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milasan, A.; Smaani, A.; Martel, C. Early rescue of lymphatic function limits atherosclerosis progression in Ldlr(-/-) mice. Atherosclerosis 2019, 283, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Milasan, A.; Ledoux, J.; Martel, C. Lymphatic network in atherosclerosis: The underestimated path. Future Sci. OA 2015, 1, FSO61. [Google Scholar] [CrossRef] [Green Version]
- Streich, K.; Smoczek, M.; Hegermann, J.; Dittrich-Breiholz, O.; Bornemann, M.; Siebert, A.; Bleich, A.; Buettner, M. Dietary lipids accumulate in macrophages and stromal cells and change the microarchitecture of mesenteric lymph nodes. J. Adv. Res. 2020, 24, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, M.B.; Sevick-Muraca, E.M. Cytokines are systemic effectors of lymphatic function in acute inflammation. Cytokine 2013, 64, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; von der Weid, P.Y. Lymphatic system: An active pathway for immune protection. Semin. Cell Dev. Biol. 2015, 38, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevaskis, N.L.; Kaminskas, L.M.; Porter, C.J. From sewer to savior—Targeting the lymphatic system to promote drug exposure and activity. Nat. Rev. Drug Discov. 2015, 14, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Hampton, H.R.; Bailey, J.; Tomura, M.; Brink, R.; Chtanova, T. Microbe-dependent lymphatic migration of neutrophils modulates lymphocyte proliferation in lymph nodes. Nat. Commun. 2015, 6, 7139. [Google Scholar] [CrossRef]
- Neeland, M.R.; Elhay, M.J.; Nathanielsz, J.; Meeusen, E.N.; de Veer, M.J. Incorporation of CpG into a liposomal vaccine formulation increases the maturation of antigen-loaded dendritic cells and monocytes to improve local and systemic immunity. J. Immunol. 2014, 192, 3666–3675. [Google Scholar] [CrossRef] [Green Version]
- Hirosue, S.; Vokali, E.; Raghavan, V.R.; Rincon-Restrepo, M.; Lund, A.W. Steady-state antigen scavenging, cross-presentation, and CD8+ T cell priming: A new role for lymphatic endothelial cells. J. Immunol. 2014, 192, 5002–5011. [Google Scholar] [CrossRef] [Green Version]
- Weitman, E.S.; Aschen, S.Z.; Farias-Eisner, G.; Albano, N.; Cuzzone, D.A.; Ghanta, S.; Zampell, J.C.; Thorek, D.; Mehrara, B.J. Obesity impairs lymphatic fluid transport and dendritic cell migration to lymph nodes. PLoS ONE 2013, 8, e70703. [Google Scholar] [CrossRef]
- Kataru, R.P.; Park, H.J.; Baik, J.E.; Li, C.; Shin, J.; Mehrara, B.J. Regulation of Lymphatic Function in Obesity. Front. Physiol. 2020, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Jalkanen, S.; Salmi, M. Lymphatic endothelial cells of the lymph node. Nat. Rev. Immunol. 2020, 20, 566–578. [Google Scholar] [CrossRef]
- Hampton, H.R.; Chtanova, T. Lymphatic Migration of Immune Cells. Front. Immunol. 2019, 10, 1168. [Google Scholar] [CrossRef]
- Ariyagunarajah, R.; Chen, H.H. To be or not to be obese: Impact of obesity on lymphatic function. J. Physiol. 2017, 595, 1449–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savetsky, I.L.; Albano, N.J.; Cuzzone, D.A.; Gardenier, J.C.; Torrisi, J.S.; GarcíaNores, G.D.; Nitti, M.D.; Hespe, G.E.; Nelson, T.S.; Kataru, R.P.; et al. Lymphatic Function Regulates Contact Hypersensitivity Dermatitis in Obesity. J. Investig. Dermatol. 2015, 135, 2742–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobedo, N.; Proulx, S.T.; Karaman, S.; Dillard, M.E.; Johnson, N.; Detmar, M.; Oliver, G. Restoration of lymphatic function rescues obesity in Prox1-haploinsufficient mice. JCI Insight 2016, 1, e85096. [Google Scholar] [CrossRef]
- Greene, A.K. Lower-extremity lymphedema and elevated. N. Engl. J. Med. 2012, 366, 2136–2137. [Google Scholar] [CrossRef] [PubMed]
- James, B.R.; Tomanek-Chalkley, A.; Askeland, E.J.; Kucaba, T.; Griffith, T.S.; Norian, L.A. Diet-induced obesity alters dendritic cell function in the presence and absence of tumor growth. J. Immunol. 2012, 189, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21(st) Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef] [PubMed]
- Aschen, S.; Zampell, J.C.; Elhadad, S.; Weitman, E.M.; Mehrara, B.J. Regulation of adipogenesis by lymphatic fluid stasis: Part II. Expression of adipose differentiation genes. Plast. Reconstr. Surg. 2012, 129, 838–847. [Google Scholar] [CrossRef]
- Escobedo, N.; Oliver, G. The Lymphatic Vasculature: Its Role in Adipose Metabolism and Obesity. Cell Metab. 2017, 26, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, S.; Scallan, J.P.; Kim, K.W.; Werth, K.; Johnson, M.W.; Saunders, B.T. CCR7 and IRF4-dependent dendritic cells regulate lymphatic collecting vessel permeability. J. Clin. Investig. 2016, 126, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.N.; Zawieja, D.C.; Goodman, A.H.; Granger, H.J. Characterization of intact mesenteric lymphatic pump and its responsiveness to acute edemagenic stress. Am. J. Physiol. 1989, 257, 2059–2069. [Google Scholar] [CrossRef]
- Von der Weid, P.Y. Review article: Lymphatic vessel pumping and inflammation–the role of spontaneous constrictions and underlying electrical pacemaker potentials. Aliment. Pharmacol. Ther. 2001, 15, 1115–1129. [Google Scholar] [CrossRef]
- Varricchi, G.; Granata, F.; Loffredo, S.; Genovese, A.; Marone, G. Angiogenesis and lymphangiogenesis in inflammatory skin disorders. J. Am. Acad. Dermatol. 2015, 73, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Cuzzone, D.A.; Weitman, E.S.; Albano, N.J.; Ghanta, S.; Savetsky, I.L.; Gardenier, J.C.; Joseph, W.J.; Torrisi, J.S.; Bromberg, J.F.; Olszewski, W.L.; et al. IL-6 regulates adipose deposition and homeostasis in lymphedema. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1426–H1434. [Google Scholar] [CrossRef]
- Von Der Weid, P.Y.; Rehal, S. Regulatory mechanisms in lymphatic gut. Ann. N. Y. Acad. Sci. 2010, 1207, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Azhar, S.H.; Lim, H.Y.; Tan, B.K.; Angeli, V. The Unresolved Pathophysiology of Lymphedema. Front. Physiol. 2020, 11, 137. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Cheng, G.; Conner, D.A.; Huang, Y.; Kucherlapati, R.S.; Munn, L.L. Impaired lymphatic contraction associated with immunosuppression. Proc. Natl. Acad. Sci. USA 2011, 108, 18784–18789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrisi, J.S.; Hespe, G.E.; Cuzzone, D.A.; Savetsky, I.L.; Nitti, M.D.; Gardenier, J.C.; GarcíaNores, G.D.; Jowhar, D.; Kataru, R.P.; Mehrara, B.J. Inhibition of Inflammation and iNOS Improves Lymphatic Function in Obesity. Sci. Rep. 2016, 6, 19817. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.S.; Karaman, S.; Proulx, S.T.; Ochsenbein, A.M.; Luciani, P.; Leroux, J.C.; Wolfrum, C.; Detmar, M. Chronic high-fat diet impairs collecting lymphatic vessel function in mice. PLoS ONE 2014, 9, e94713. [Google Scholar] [CrossRef]
- Liao, S.; von der Weid, P.Y. Inflammation-induced lymphangiogenesis and lymphatic dysfunction. Angiogenesis 2014, 17, 325–334. [Google Scholar] [CrossRef]
- D’Alessio, S.; Tacconi, C.; Danese, S. Targeting lymphatics in inflammatory bowel disease. Oncotarget 2015, 6, 34047–34048. [Google Scholar] [CrossRef] [PubMed]
- Rockson, S.G. The lymphatics and the inflammatory response: Lessons learned from human lymphedema. Lymphat. Res. Biol. 2013, 11, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, A.; Avraham, T.; Zampell, J.C.; Haviv, Y.S.; Weitman, E.; Mehrara, B.J. Adipose-derived stem cells promote lymphangiogenesis in response to VEGF-C stimulation or TGF-β1 inhibition. Future Oncol. 2011, 7, 1457–1473. [Google Scholar] [CrossRef] [Green Version]
- Oka, M.; Iwata, C.; Suzuki, H.I.; Kiyono, K.; Morishita, Y.; Watabe, T.; Komuro, A.; Kano, M.R.; Miyazono, K. Inhibition of endogenous TGF-beta signaling enhances lymphangiogenesis. Blood 2008, 111, 4571–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataru, R.P.; Jung, K.; Jang, C.; Yang, H.; Schwendener, R.A.; Baik, J.E.; Han, S.H.; Alitalo, K.; Koh, G.Y. Critical role of CD11b+ macrophages and VEGF in inflammatory lymphangiogenesis, antigen clearance, and inflammation resolution. Blood 2009, 113, 5650–5659. [Google Scholar] [CrossRef] [Green Version]
- Wada, H.; Ura, S.; Kitaoka, S.; Satoh-Asahara, N.; Horie, T.; Ono, K.; Takaya, T.; Takanabe-Mori, R.; Akao, M.; Abe, M.; et al. Distinct characteristics of circulating vascular endothelial growth factor-a and C levels in human subjects. PLoS ONE 2011, 6, e29351. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, S.K.; Jin, Y.; Goyal, S.; Lee, H.S.; Fuchsluger, T.A.; Lee, H.K.; Dana, R. A novel pro-lymphangiogenic function for Th17/IL-17. Blood 2011, 118, 4630–4634. [Google Scholar] [CrossRef] [Green Version]
- Choi, I.; Lee, Y.S.; Chung, H.K.; Choi, D.; Ecoiffier, T.; Lee, H.N.; Kim, K.E.; Lee, S.; Park, E.K.; Maeng, Y.S.; et al. Interleukin-8 reduces post-surgical lymphedema formation by promoting lymphatic vessel regeneration. Angiogenesis 2013, 16, 29–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savetsky, I.L.; Ghanta, S.; Gardenier, J.C.; Torrisi, J.S.; GarcíaNores, G.D.; Hespe, G.E.; Nitti, M.D.; Kataru, R.P.; Mehrara, B.J. Th2 cytokines inhibit lymphangiogenesis. PLoS ONE 2015, 10, 0126908. [Google Scholar] [CrossRef] [PubMed]
- Zampell, J.C.; Avraham, T.; Yoder, N.; Fort, N.; Yan, A.; Weitman, E.S.; Mehrara, B.J. Lymphatic function is regulated by a coordinated expression of lymphangiogenic and anti-lymphangiogenic cytokines. Am. J. Physiol. Cell Physiol. 2012, 302, C392–C404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avraham, T.; Daluvoy, S.; Zampell, J.; Yan, A.; Haviv, Y.S.; Rockson, S.G.; Mehrara, B.J. Blockade of transforming growth factor-beta1 accelerates lymphatic regeneration during wound repair. Am. J. Pathol. 2010, 177, 3202–3214. [Google Scholar] [CrossRef]
- Shimizu, Y.; Shibata, R.; Ishii, M.; Ohashi, K.; Kambara, T.; Uemura, Y.; Yuasa, D.; Kataoka, Y.; Kihara, S.; Murohara, T.; et al. Adiponectin-mediated modulation of lymphatic vessel formation and lymphedema. J. Am. Heart Assoc. 2013, 2, e000438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cromer, W.E.; Zawieja, S.D.; Tharakan, B.; Childs, E.W.; Newell, M.K.; Zawieja, D.C. The effects of inflammatory cytokines on lymphatic endothelial barrier function. Angiogenesis 2014, 17, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Schwager, S.; Detmar, M. Inflammation and Lymphatic Function. Front. Immunol. 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Cifarelli, V.; Appak-Baskoy, S.; Peche, V.S.; Kluzak, A.; Shew, T.; Narendran, R.; Pietka, K.M.; Cella, M.; Walls, C.W.; Czepielewski, R.; et al. Visceral obesity and insulin resistance associate with CD36 deletion in lymphatic endothelial cells. Nat. Commun. 2021, 12, 3350. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.A.; Vasile, E.; Feng, D.; Sundberg, C.; Brown, L.F.; Detmar, M.J.; Lawitts, J.A.; Benjamin, L.; Tan, X.; Manseau, E.J.; et al. Vascular permeability factor/vascular endothelial growth factor induces lymphangiogenesis as well as angiogenesis. J. Exp. Med. 2002, 196, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.J.; Davis, A.M.; Ku, C.W.; Gashev, A.A. Myogenic constriction and dilation of isolated lymphatic vessels. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Telinius, N.; Baandrup, U.; Rumessen, J.; Pilegaard, H.; Hjortdal, V.; Aalkjaer, C.; Boedtkjer, D.B. The human thoracic duct is functionally innervated by adrenergic nerves. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, 206–213. [Google Scholar] [CrossRef]
- D’Andrea, V.; Bianchi, E.; Taurone, S.; Mignini, F.; Cavallotti, C.; Artico, M. Cholinergic innervation of human mesenteric lymphatic vessels. Folia Morphol. 2013, 72, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Krieglstein, C.F.; Anthoni, C.; Cerwinka, W.H.; Stokes, K.Y.; Russell, J.; Grisham, M.B.; Granger, D.N. Role of blood- and tissue-associated inducible nitric-oxide synthase in colonic inflammation. Am. J. Pathol. 2007, 170, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Schönbein, G.W. Nitric oxide (NO) side of lymphatic flow and immune surveillance. Proc. Natl. Acad. Sci. USA 2012, 109, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Muthuchamy, M.; Zawieja, D. Molecular regulation of lymphatic contractility. Ann. N. Y. Acad. Sci. 2008, 1131, 89–99. [Google Scholar] [CrossRef]
- Lukacs-Kornek, V.; Malhotra, D.; Fletcher, A.L.; Acton, S.E.; Elpek, K.G.; Tayalia, P.; Collier, A.R.; Turley, S.J. Regulated release of nitric oxide by nonhematopoieticstroma controls expansion of the activated T cell pool in lymph nodes. Nat. Immunol. 2011, 12, 1096–1104. [Google Scholar] [CrossRef]
- Breslin, J.W.; Gaudreault, N.; Watson, K.D.; Reynoso, R.; Yuan, S.Y.; Wu, M.H. Vascular endothelial growth factor-C stimulates the lymphatic pump by a VEGF receptor-3-dependent mechanism. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Bouta, E.M.; Kuzin, I.; de Mesy Bentley, K.; Wood, R.W.; Rahimi, H.; Ji, R.C.; Ritchlin, C.T.; Bottaro, A.; Xing, L.; Schwarz, E.M. Brief Report: Treatment of Tumor Necrosis Factor-Transgenic Mice With Anti-Tumor Necrosis Factor Restores Lymphatic Contractions, Repairs Lymphatic Vessels, and May Increase Monocyte/Macrophage Egress. Arthritis Rheumatol. 2017, 69, 1187–1193. [Google Scholar] [CrossRef] [Green Version]
- Polzer, K.; Baeten, D.; Soleiman, A.; Distler, J.; Gerlag, D.M.; Tak, P.P.; Schett, G.; Zwerina, J. Tumour necrosis factor blockade increases lymphangiogenesis in murine and human arthritic joints. Ann. Rheum. Dis. 2008, 67, 1610–1616. [Google Scholar] [CrossRef]
- Földi, M.; Strossenreuther, R. Foundations of Manual Lymph Drainage, 3rd ed.; Mosby Publisher, Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Loskotová, A.; Loskotová, J.; Suchanek, I.; Brychta, P.; Lipový, B. Myofascial-manual lymphatic drainage for burn trauma: A service evaluation. Br. J. Commun. Nurs. 2017, 1, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.M.; Yeun, Y.R.; Kim, H.Y.; Kim, S.J. Effects of manual lymph drainage for abdomen on the brain activity of subjects with psychological stress. J. Phys. Ther. Sci. 2017, 29, 491–494. [Google Scholar] [CrossRef] [PubMed]
- de Godoy, J.M.; Santana, K.R.; Godoy, M. Lymphoscintigraphic evaluation of manual lymphatic therapy: The Godoy & Godoy technique. Phlebology 2015, 30, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Tan, I.C.; Maus, E.A.; Rasmussen, J.C.; Marshall, M.V.; Adams, K.E.; Fife, C.E.; Smith, L.A.; Chan, W.; Sevick-Muraca, E.M. Assessment of lymphatic contractile function after manual lymphatic drainage using near-infrared fluorescence imaging. Arch. Phys. Med. Rehabil. 2011, 92, 756–764.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaedú, R.; Mello, M.; Tucunduva, R.A.; daSilveira, J.; Takahashi, M.; Valente, A. Postoperative Orthognathic Surgery Edema Assessment With and Without Manual Lymphatic Drainage. J. Craniofacial Surg. 2017, 28, 1816–1820. [Google Scholar] [CrossRef]
- Müller, M.; Klingberg, K.; Wertli, M.M.; Carreira, H. Manual lymphatic drainage and quality of life in patients with lymphoedema and mixed oedema: A systematic review of randomised controlled trials. Quality of life research. Int. J. Qual. Life Asp. Treat. Care Rehabil. 2018, 27, 1403–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, B.; Gaitatzis, K.; Janse de Jonge, X.; Blackwell, R.; Koelmeyer, L.A. Manual lymphatic drainage treatment for lymphedema: A systematic review of the literature. J. Cancer Surviv. Res. Pract. 2021, 15, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Belgrado, J.P.; Vandermeeren, L.; Vankerckhove, S.; Valsamis, J.B.; Malloizel-Delaunay, J.; Moraine, J.J.; Liebens, F. Near-Infrared Fluorescence Lymphatic Imaging to Reconsider Occlusion Pressure of Superficial Lymphatic Collectors in Upper Extremities of Healthy Volunteers. Lymphat. Res. Biol. 2016, 14, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Kramp, M.E. Combined manual therapy techniques for the treatment of women with infertility: A case series. J. Am. Osteopath. Assoc. 2012, 112, 680–684. [Google Scholar]
- Kutlay, S.; Ozdemir, E.C.; Pala, Z.; Ozen, S.; Sanli, H. Complete Decongestive Therapy Is an Option for the Treatment of Rosacea Lymphedema (Morbihan Disease): Two Cases. Phys. Ther. 2019, 1, 406–410. [Google Scholar] [CrossRef]
- Arngrim, N.; Simonsen, L.; Holst, J.J.; Bülow, J. Reduced adipose tissue lymphatic drainage of macromolecules in obese subjects: A possible link between obesity and local tissue inflammation? Int. J. Obes. 2013, 37, 748–750. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, D.F.; de Oliveira, P.; Gimenes, A.S.; Moreno, M.A. Postural drainage and manual lymphatic drainage for lower limb edema in women with morbid obesity after bariatric surgery: A randomized controlled trial. Am. J. Phys. Med. Rehabil. 2013, 92, 697–703. [Google Scholar] [CrossRef]
- Antoniak, K.; Hansdorfer-Korzon, R.; Wojtowicz, D.; Koziński, M.; Zorena, K. The influence of manual therapy on biometric parameters in patients with improper body weight. In Proceedings of the 3rd International Electronic Conference on Environmental Research and Public Health —Public Health Issues in the Context of the COVID-19 Pandemic, online, 11–25 January 2021; MDPI: Basel, Switzerland, 2021. [Google Scholar] [CrossRef]
- Antoniak, K. Physiotherapeutic treatment as an aid in lymphatic function in obese patients. In Webinar on Obesity and Weight Loss; PHRONESIS LLC.: Malvern, PA, USA, 2021; pp. 23–24. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Cell Type | Synthesis of Factors | Metabolic Effects | Source |
|---|---|---|---|
| ↑ Macrophages Polarization of M2 macrophages towards M1 inflammatory macrophages | ↑ TNF-α, IL-1β, IL-6, IL-12, IL-23 ↑ iNOS activation and ↑ NO ↓ IL-10 | ↑ inflammation | [13,14,15,16,17,20] |
| ↑ insulin resistance | |||
| ↑ risk of atherosclerosis development | |||
| ↑ risk of hypertension development | |||
| ↑ Neutrophils | ↑ TNF-α, MCP-1, IL-1β | ↑ macrophage infiltration | [20,21,22] |
| ↑ insulin resistance | |||
| ↑ Mast cells | ↑ IL-1, IL-3, IL-5, IL-6 IL-8, TNF-α, histamines, heparins, tryptases, chymases | ↑ macrophage infiltration remodeling of adipose tissue | [23,24] |
| ↓ Treg | ↓ IL-1β, IL-6, IL-8, TNF-α, MIP-1α | ↑ inflammation | [27,28,29] |
| ↑ insulin resistance | |||
| ↑ M1 macrophage infiltration | |||
| ↓ glucose tolerance | |||
| ↑ CD4+ Th1 | ↑ IFN-γ | ↑ insulin resistance | [27,29,30] |
| ↑ risk of atherosclerosis development | |||
| ↑ risk of hypertension development | |||
| change in phenotype of M2 macrophages towards M1 macrophages | |||
| ↓ CD4+ Th2 | ↓ IL-4, IL-5, IL-13, IL-10 | ↓ glucose tolerance | [32,33,34] |
| ↑ CD4+ Th17 | ↑ TNF-α and IL-17 | ↑ insulin resistance | [32,33,34] |
| ↑ risk of atherosclerosis development | |||
| ↑ risk of hypertension development | |||
| change in phenotype of M2 macrophages towards M1 macrophages | |||
| ↑ T CD8+ | ↑ TNF- α, IL-2, IFN- γ and RANTES chemokines | ↑ insulin resistance | [32,33,34] |
| ↓ B Cells | ↑ IL-6 | activation of pro-inflammatory CD4+ and CD8+ lymphocytes | [34,35,36,37] |
| ↓ IL-10 | change in phenotype of M2 macrophages towards M1 macrophages | ||
| ↑ Saturated Fatty Acids—as a metabolites odd-chain saturated fatty acids even-chain saturated fatty acids | ↑ MCP-1 and TNF-α | change in phenotype of M2 macrophages towards M1 macrophages | [40] |
| ↑ resistin | odd-chain saturated fatty acids are associated with a favorable adipokine profile, whereas even-chain saturated fatty acids are associated with an unfavorable profile. | [39] | |
| ↓ adiponectin | |||
| ↑ visfatin | |||
| ↓ leptin and PAI-1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoniak, K.; Hansdorfer-Korzon, R.; Mrugacz, M.; Zorena, K. Adipose Tissue and Biological Factors. Possible Link between Lymphatic System Dysfunction and Obesity. Metabolites 2021, 11, 617. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090617
Antoniak K, Hansdorfer-Korzon R, Mrugacz M, Zorena K. Adipose Tissue and Biological Factors. Possible Link between Lymphatic System Dysfunction and Obesity. Metabolites. 2021; 11(9):617. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090617
Chicago/Turabian StyleAntoniak, Klaudia, Rita Hansdorfer-Korzon, Małgorzata Mrugacz, and Katarzyna Zorena. 2021. "Adipose Tissue and Biological Factors. Possible Link between Lymphatic System Dysfunction and Obesity" Metabolites 11, no. 9: 617. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11090617