
The Response of Cowpea (Vigna unguiculata) Plants to Three Abiotic Stresses Applied with Increasing Intensity: Hypoxia, Salinity, and Water Deficit


Abstract
:1. Introduction
2. Results
2.1. Soil Parameters Following Treatments
2.2. Morphological and Physiological Parameters of Cowpea Plants Following Three Treatments
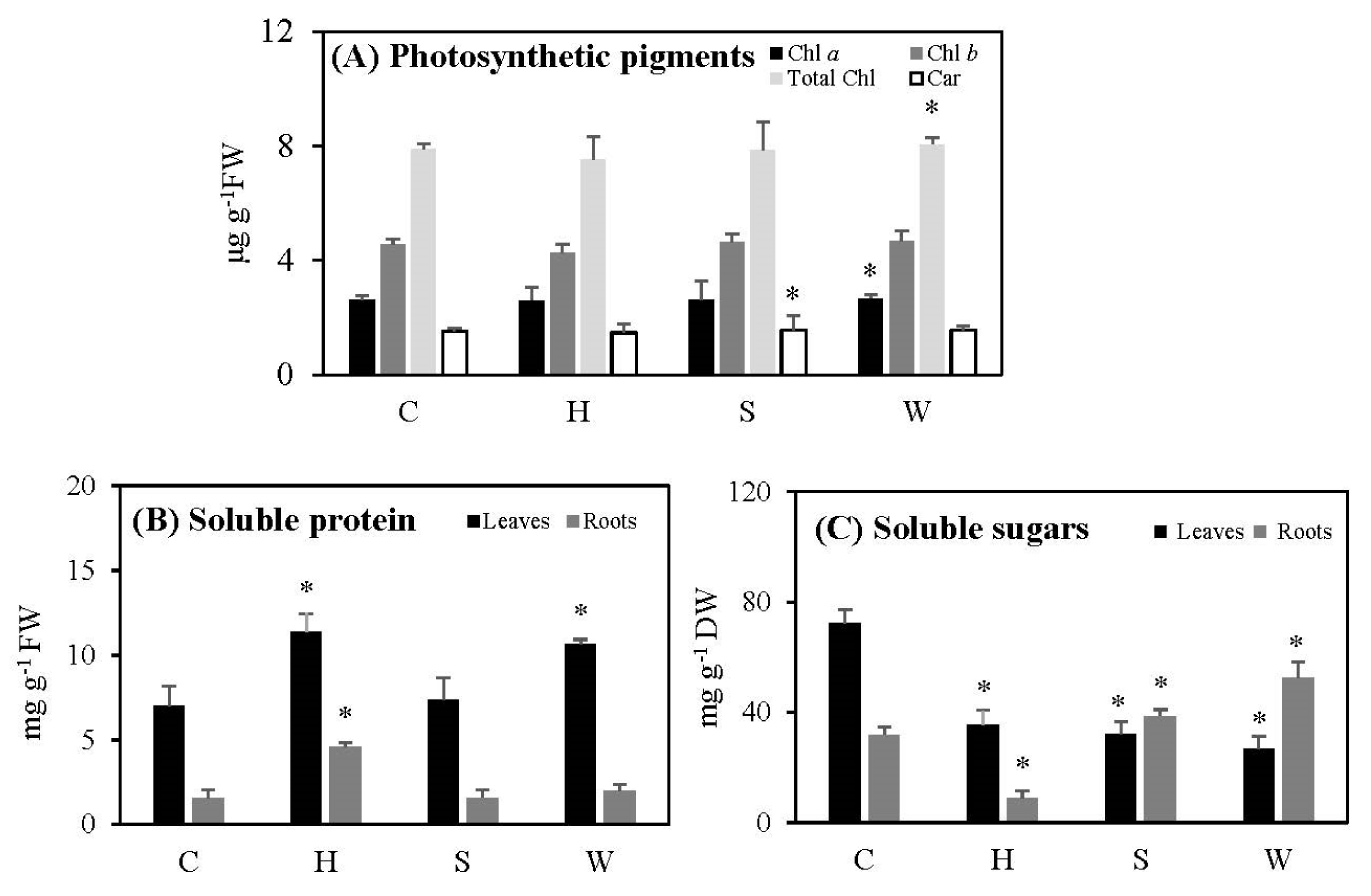
2.3. Photosynthetic Pigments, Total Soluble Sugars, and Protein Content Following the Three Treatments
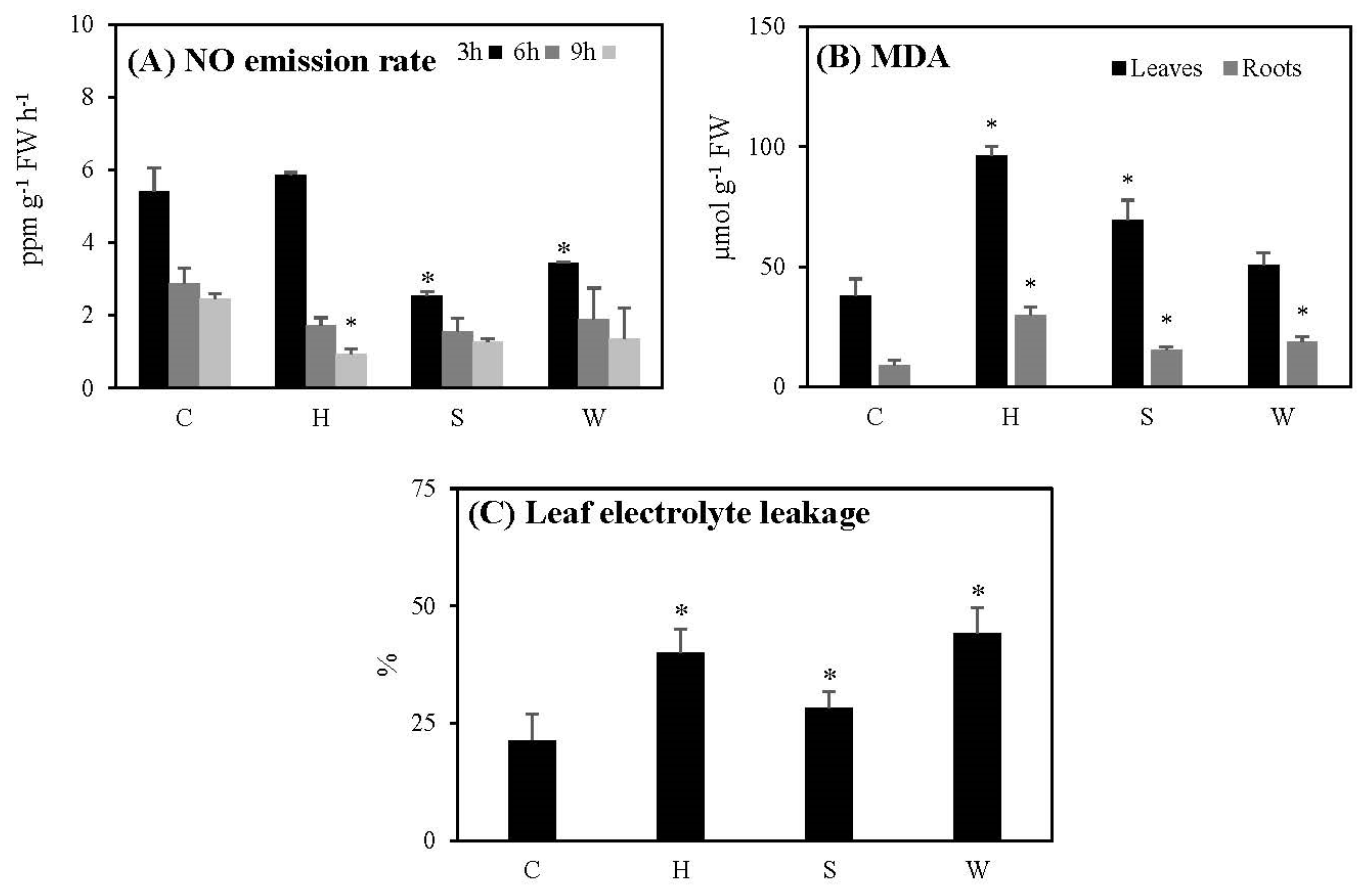
2.4. NO Production, Lipid Peroxidation, and Cell Damage Following the Three Treatments
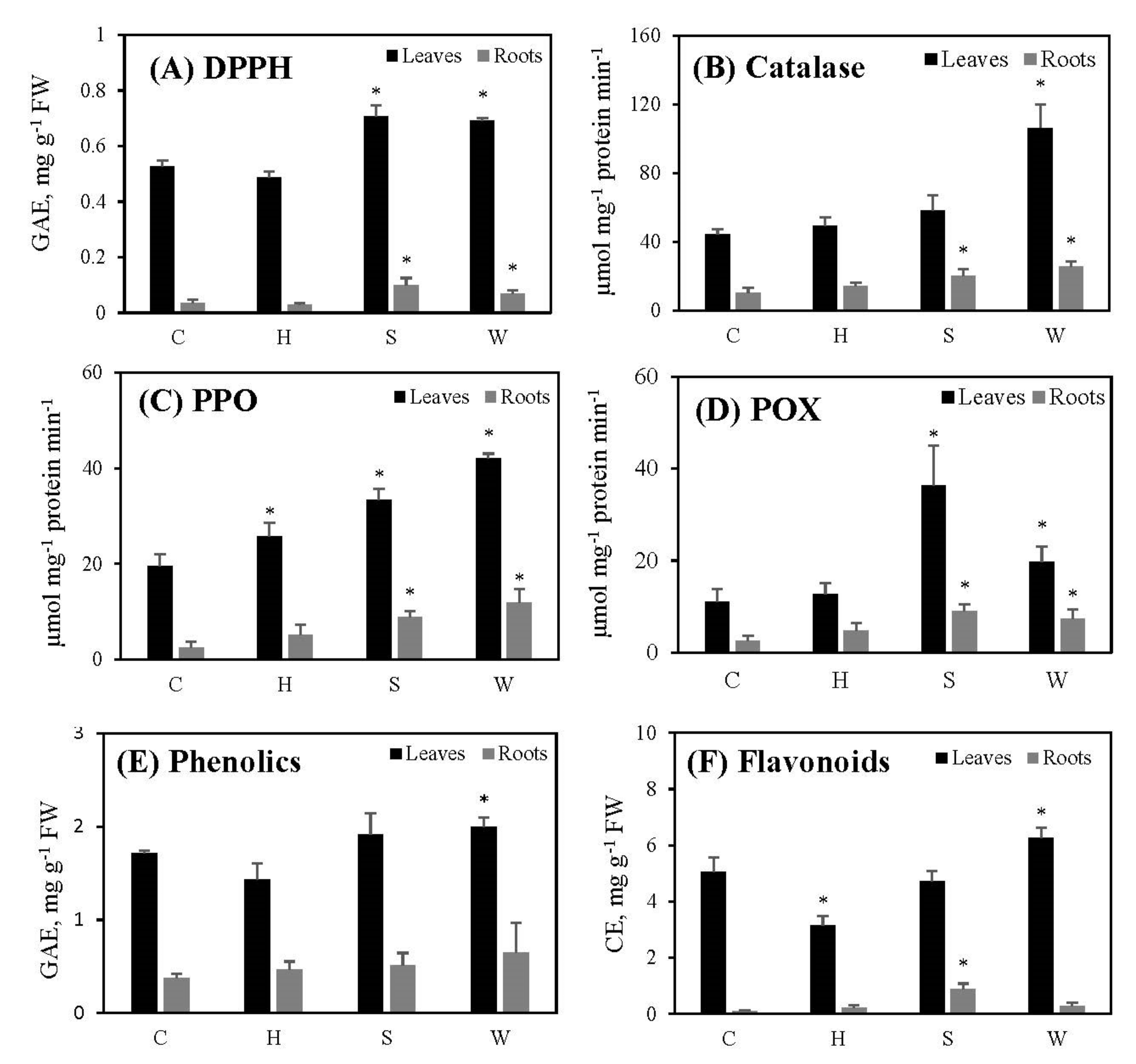
2.5. Antioxidant Capacity of Cowpea Plants Following Hypoxia and Re-Oxygenation
3. Discussion
3.1. Soil Parameters
3.2. Plant Growth
3.3. Photosynthetic Pigments, Sugars, and Protein Contents
3.4. Nitric Oxide (NO) Production
3.5. Cell Damage
3.6. Antioxidant Capacity
4. Material and Methods
4.1. Plant Materials and Growth Conditions
4.2. Treatments and Sampling
4.3. Soil Parameters
4.4. Growth Parameters
4.5. Nitric Oxide (NO) Emission, Lipid Peroxidation, and Leaf Electrolyte Leakage
4.6. Antioxidant Compounds and Enzyme Activities
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carvalho, B.M.; Viana, A.P.; dos Santos, P.H.D.; Generoso, A.L.; Corrêa, C.C.G.; Silveira, V.; Eiras, M.; Santos, E.A. Proteome of resistant and susceptible Passiflora species in the interaction with cowpea aphid-borne mosaic virus reveals distinct responses to pathogenesis. Euphytica 2019, 215, 167. [Google Scholar] [CrossRef]
- Salinas-Gamboa, R.; Johnson, S.D.; Sánchez-León, N.; Koltunow, A.M.; Vielle-Calzada, J.P. New observations on gametogenic development and reproductive experimental tools to support seed yield improvement in cowpea [Vigna unguiculata (L.) Walp]. Plant Reprod. 2016, 29, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A.E. Phenotyping cowpeas for adaptation to drought. Front. Physiol. 2012, 3, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timko, M.P.; Ehlers, J.D.; Roberts, P.A. Cowpea. In Genome Mapping and Molecular Breeding in Plants, Pulses, Sugar and Tuber Crops; Kole, C., Ed.; Springer: Berlin, Germany, 2007; Volume 3, pp. 49–67. [Google Scholar]
- Agbicodo, E.M.; Fatokun, C.A.; Muranaka, S.; Visser, R.G.F. Breeding drought tolerant cowpea: Constraints, accomplishments, and future prospects. Euphytica 2009, 167, 353–370. [Google Scholar] [CrossRef] [Green Version]
- Merwad, A.R.M.A.; Desoky, E.S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Planchet, E.; Gupta, K.J.; Sonoda, M.; Kaiser, W.M. Nitric oxide emission from tobacco leaves and cell suspensions: Rate limiting factors and evidence for the involvement of mitochondrial electron transport. Plant J. 2005, 41, 732–743. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Hashem, A.; Abd Allah, E.F.; Ahmad, P. Arbuscular mycorrhiza in crop improvement under environmental stress. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Rasool, S., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 2, pp. 69–95. [Google Scholar] [CrossRef]
- Abeer, H.; Abd Allah, E.F.; Alqarawi, A.A.; Alwhibi Mona, S.; Alenazi, M.M.; Egamberdieva, D.; Ahmad, P. Arbuscular mycorrhizal fungi mitigates NaCl induced adverse effects on Solanum lycopersicum L. Pakistan J. Bot. 2015, 47, 327–340. [Google Scholar]
- Zimmermann, M.J.; Rocha, M.; Yolanda, T. Bean Crop: Factors Affecting Productivity; Brazilian Association for the Presence of Potash and Phosphate: Piracicaba, Brazil, 1988; pp. 317–340. [Google Scholar]
- Shavrukov, Y. Salt stress or salt shock: Which genes are we studying? J. Exp. Bot. 2013, 64, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Ghosh, P.; Roychoudhury, A. Salt acclimation differentially regulates the metabolites commonly involved in stress tolerance and aroma synthesis in indica rice cultivars. Plant Growth Regul. 2019, 88, 87–97. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Lippold, F.; Redestig, H.; Hannah, M.A.; Erban, A.; Krämer, U.; Kopka, J.; Udvardi, M.K. Integrative functional genomics of salt acclimatization in the model legume Lotus japonicus. Plant J. 2008, 53, 973–987. [Google Scholar] [CrossRef]
- Krishnamurthy, L.C.; Johansen, C.; Ito, O. Genotypic variation in root system development and its implication for drought resistance in Chickpea. In Roots and Nitrogen in Cropping Systems of the Semiarid Tropics; Ito, O., Johansen, C., Adu-Gyamfi, J.J., Katayama, K., Rao, J.V.K., Rego, T.J., Eds.; JIRCAS and ICRISAT: Hyderabad, India, 1996; pp. 235–250. [Google Scholar]
- Mishra, S.; Sahu, G.; Shaw, B.P. Insight into the cellular and physiological regulatory modulations of Class-I TCP9 to enhance drought and salinity stress tolerance in cowpea. Physiol. Plant. 2021, in press. [Google Scholar] [CrossRef]
- Ferreira-Neto, J.; Borges, A.; da Silva, M.D.; Morais, D.; Bezerra-Neto, J.P.; Bourque, G.; Kido, E.A.; Benko-Iseppon, A.M. The cowpea kinome: Genomic and transcriptomic analysis under biotic and abiotic stresses. Front. Plant Sci. 2021, 12, 667013. [Google Scholar] [CrossRef]
- Galmés, J.; Medrano, H.; Flexas, J. Acclimation of Rubisco specificity factor to drought in tobacco: Discrepancies between in vitro and in vivo estimations. J. Exp. Bot. 2006, 57, 3659–3667. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Carriquí, M.; Coopman, R.E.; Gago, J.; Galmés, J.; Martorell, S.; Morales, F.; Diaz Espejo, A. Stomatal and mesophyll conductances to CO2 in different plant groups: Underrated factors for predicting leaf photosynthesis responses to climate change? Plant Sci. 2014, 226, 41–48. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Abd Allah, E.F. Mycorrhizal Association and ROS in Plants. In Oxidative Damage to Plants; Ahmad, P., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 453–475. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-talk between nitric oxide, hydrogen peroxide and calcium in salt-stressed Chenopodium quinoa Willd. at seed germination stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef]
- Urrestarazu, M.; Mazuela, P.M. Effect of slow-release oxygen supply by fertigation on horticultural crops under soilless culture. Sci. Hortic. 2005, 106, 484–490. [Google Scholar] [CrossRef]
- Dat, J.; Folzer, H.; Parent, C.; Badot, P.M.; Capelli, N. Hypoxia stress. Current understanding and perspectives. In Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues; Teixeira da Silva, J.A., Ed.; Global Science Books: Isleworth, UK, 2006; Volume 3, pp. 664–674. [Google Scholar]
- Mane, A.; Saratale, G.; Karadge, B.; Samant, J. Studies on the effects of salinity on growth, polyphenol content and photosynthetic response in Vetiveria zizanioides (L.) Nash. Emirates J. Food Agric. 2017, 23, 59–70. [Google Scholar] [CrossRef]
- Santiago, S.; Martha, S.; Sofía, T.S.; Jorge, M. The role of nitric oxide in nitrogen fixation by legumes. Front. Plant Sci. 2020, 11, 521. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Papadopoulou, E.; Rennenberg, H. Interaction of flooding with carbon metabolism, of forest trees. Plant Biol. 2004, 6, 299–306. [Google Scholar] [CrossRef]
- Herrera, A. Responses to flooding of plant water relations and leaf gas exchange in tropical tolerant trees of a black-water wetland. Front. Plant Sci. 2013, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- León, J.; Castillo, M.C.; Gayubas, B. The hypoxia–reoxygenation stress in plants. J. Exp. Bot. 2021, 72, 5841–5856. [Google Scholar] [CrossRef]
- Onwugbuta-Enyi, J. Water balance and proximate composition in cowpea (Vigna unguiculata (L.) Walp) seedlings exposed to drought and flooding stress. J. Appl. Sci. Environ. Manag. 2004, 8, 55–57. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; López-Aguilar, R.; Kaya, C.; Larrinaga-Mayoral, J.A.; Flores-Hernández, A. Comparative effects of NaCl and polyethyleneg mergence and seedling growth of cowpea. J. Agron. Crop Sci. 2002, 188, 235–247. [Google Scholar] [CrossRef]
- Le, L.T.T.; Kotula, L.; Siddique, K.H.M.; Colmer, T.D. Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2021, 22, 1909. [Google Scholar] [CrossRef]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.I.A.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A.S.; et al. Exogenous Sodium Nitroprusside Mitigates Salt Stress in Lentil (Lens culinaris Medik.) by Affecting the Growth, Yield, and Biochemical Properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef]
- Maamallan, S.; Prakash, M.; Sathiyanarayanan, G.; Rameshkumar, S. Effect of Seed Hardening and Pelleting on Germination and Seedling Attributes of Cowpea under Saline Condition. Legume Res. 2021, 44, 723–729. [Google Scholar] [CrossRef]
- Dadson, R.B.; Hashem, F.M.; Javaid, I.; Joshi, J.; Allen, A.L.; Devine, T.E. Effect of water stress on the yield of cowpea (Vigna unguiculata [L.] Walp.) genotypes in the Delmarva Region of the United States. J. Agron. Crop Sci. 2005, 191, 210–217. [Google Scholar] [CrossRef]
- Singh, H.; Reddy, M.S. Effect of inoculation with phosphate solubilizing fungus on growth and nutrient uptake of wheat and maize plants fertilized with rock phosphate in alkaline soils. Eur. J. Soil Biol. 2011, 47, 30–34. [Google Scholar] [CrossRef]
- Singh, S.K.; Kakani, V.G.; Surabhi, G.K.; Reddy, K.R. Cowpea (Vigna unguiculata [L.] Walp.) genotypes response to multiple abiotic stresses. J. Photochem. Photobiol. B 2010, 100, 135–146. [Google Scholar] [CrossRef]
- Saradadevi, R.; Bramley, H.; Siddique, K.H.M.; Edwards, E.; Palta, J.A. Contrasting stomatal regulation and leaf ABA concentrations in wheat genotypes when split root systems were exposed to terminal drought. Field Crops Res. 2014, 162, 77–86. [Google Scholar] [CrossRef]
- Kumar, R.S.; Singh, S.; Nandwal, A.S.; Waldia, R.S.; Sharma, M.K. Genotypic differences in water status, membrane integrity, ionic content, N2-fixing efficiency and dry matter of mung bean nodules under saline irrigation. Physiol. Mol. Biol. Plants 2008, 14, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rane, J.; Raina, S.K.; Govindasamy, V.; Bindumadhava, H.; Hanjagi, P.; Giri, R.; Jangid, K.K.; Kumar, M.; Nair, R.M. Use of phenomics for differentiation of mungbean (Vigna radiata L. Wilczek) genotypes varying in growth rates per unit of water. Front. Plant Sci. 2021, 12, 692564. [Google Scholar] [CrossRef] [PubMed]
- Hajihashemi, S.; Brestic, M.; Landi, M.; Skalicky, M. Resistance of Fritillaria imperialis to freezing stress through gene expression, osmotic adjustment and antioxidants. Sci. Rep. 2020, 10, 10427. [Google Scholar] [CrossRef]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric oxide: A multitasked signaling gas in plants. Mol. Plant 2015, 8, 506–520. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Ratcliffe, R.G.; Gupta, K.J. Plant mitochondria: Source and target for nitric oxide. Mitochondrion 2014, 19, 329–333. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.B.; Al Mahmud, J.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd Allah, E.F.; Gucel, S.; Tran, L.S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Shao, R.; Wang, K.; Shangguan, Z. Cytokinin-induced photosynthetic adaptability of Zea mays L. to drought stress associated with nitric oxide signal: Probed by ESR spectroscopy and fast OJIP fluorescence rise. J. Plant Physiol. 2010, 167, 472–479. [Google Scholar] [CrossRef]
- Blokhina, O.B.; Fagerstedt, K.V.; Chirkova, T.V. Relationships between lipid peroxidation and anoxia tolerance in a range of species during post anoxic reaeration. Physiol. Plant. 1999, 105, 625–632. [Google Scholar] [CrossRef]
- Nair, A.S.; Abraham, T.K.; Jaya, D.S. Studies on the changes in lipid peroxidation and antioxidants in drought stress induced cowpea (Vigna unguiculata L.) varieties. J. Environ. Biol. 2008, 29, 689–691. [Google Scholar]
- Noctor, G.; Foyer, C. Ascorbate and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Bansal, R.J.; Srivastava, J.P. Antioxidative defense system in pigeonpea roots under waterlogging stress. Acta Physiol. Plant. 2012, 34, 515–522. [Google Scholar] [CrossRef]
- Garnczarska, M.; Bednarski, W. Effect of a short-term hypoxic treatment followed by re-aeration on free radicals level and antioxidative enzymes in lupine roots. Plant Physiol. Biochem. 2004, 42, 233–240. [Google Scholar] [CrossRef]
- Prashanth, S.R.; Sadhasivam, V.; Parida, A. Over expression of cytosolic copper/zinc superoxide dismutase from a mangrove plant Avicennia marina in indica rice var Pusa Basmati-1 confers abiotic stress tolerance. Transgenic Res. 2008, 7, 281–291. [Google Scholar] [CrossRef]
- Cochrane, D.W.; Shah, J.K.; Hebelstrup, K.H.; Igamberdiev, A.U. Expression of phytoglobin affects nitric oxide metabolism and energy state of barley plants exposed to anoxia. Plant Sci. 2017, 265, 124–130. [Google Scholar] [CrossRef]
- Zafari, S.; Hebelstrup, K.H.; Igamberdiev, A.U. Transcriptional and metabolic changes associated with phytoglobin expression during germination of barley seeds. Int. J. Mol. Sci. 2020, 21, 2796. [Google Scholar] [CrossRef] [Green Version]
- Bang-Andreasen, T.; Nielsen, J.T.; Voriskova, J.; Heise, J.; Rønn, R.; Hansen, H.C.B.; Jacobsen, C.S. Wood ash induced pH changes strongly affect soil bacterial numbers and community composition. Front. Microbiol. 2017, 8, 1400. [Google Scholar] [CrossRef]
- Ahmad, R.; Kim, M.D.; Back, K.H.; Kim, H.S.; Lee, H.S.; Kwon, S.Y.; Murata, N.; Chung, W.I.; Kwak, S.S. Stress-induced expression of choline oxidase in potato plant chloroplasts confers enhanced tolerance to oxidative, salt, and drought stresses. Plant Cell Rep. 2008, 27, 687–698. [Google Scholar] [CrossRef]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric analysis of chlorophylls and carotenoids from commonly grown fern species by using various extracting solvents. Res. J. Chem. Sci. 2014, 4, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Ferris, R.; Long, L.; Bunn, S.M. Leaf stomatal and epidermal cell development: identification of putative quantitative trait loci in relation to elevated carbon dioxide concentration in poplar. Tree Physiol. 2002, 22, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [Green Version]
- Farrant, J.M. Mechanisms of desiccation tolerance in Angiosperm resurrection plants. In Plant Desiccation Tolerance; Jenks, M.A., Wood, A.J., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 51–90. ISBN 0813812631. [Google Scholar]
- Qiao, Y.; Zhang, H.; Dong, B.; Shi, C.; Li, Y.; Zhai, H.; Liu, M. Effects of elevated CO2 concentration on growth and water use efficiency of winter wheat under two soil water regimes. Agric. Water Manag. 2010, 97, 1742–1748. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Biochem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shah, J.K.; Cochrane, D.W.; De Paepe, R.; Igamberdiev, A.U. Respiratory complex I deficiency results in low nitric oxide levels, induction of hemoglobin and upregulation of fermentation pathways. Plant Physiol. Biochem. 2013, 63, 185–190. [Google Scholar] [CrossRef]
- Heath, R.L.; Parker, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N.; Hackley, E. New substrate for fluorometric determination of oxidative enzymes. Ann. Chem. 1967, 39, 271–278. [Google Scholar] [CrossRef]
- Raymond, J.; Pakariyathan, N.; Azanza, J.L. Purification and some properties of polyphenol oxidases from sunflowers seed. Phytochemistry 1993, 34, 927–931. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histoenzymology; Kalyani Publishers: New Delhi, India, 1980; p. 286. [Google Scholar]
- Vyas, P.; Debnath, S.C.; Igamberdiev, A.U. Metabolism of glutathione and ascorbate in lingonberry cultivars during in vitro and ex vitro propagation. Biol. Plant. 2013, 57, 603–612. [Google Scholar] [CrossRef]
- Chandrasekara, N.; Shahidi, F. Effect of roasting on phenolic content and antioxidant activities of whole cashew nuts, kernels, and testa. J. Agric. Food Chem. 2011, 59, 5006–5014. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity using the DPPH free radical method. Lebensm. Wissenchaft Technol. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Parameter | Control | Post-Hypoxia | Salt Stress | Water Deficit |
|---|---|---|---|---|
| pH | 5.46 ± 0.04 | 5.85 ± 0.02 * | 6.12 ± 0.03 * | 5.66 ± 0.03 * |
| Conductivity (µS/cm) | 352.0 ± 5.9 | 198.8 ± 8.3 * | 6332.8 ± 209.3 * | 681.8 ± 30.8 * |
| Total dissolved solids (ppm) | 185.8 ± 6.3 | 100.3 ± 4.7 * | 3166.3 ± 104.5 * | 382.5 ± 44.7 * |
| Parameter | Control | Post-Hypoxia | Salinity | Water Deficit | |
|---|---|---|---|---|---|
| Shoot length (cm) | 112.0 ± 6.7 | 100.9 ± 4.0 * | 77.6 ± 8.4 * | 109.2 ± 9.6 | |
| Root length (cm) | 9.2 ± 1.4 | 6.6 ± 0.8 * | 6.7 ± 0.7 * | 17.3 ± 4.1 * | |
| Number of compound leaves | 10 ± 1.6 | 11 ± 1.0 | 7 ± 1.9 * | 7 ± 1.3 * | |
| Leaflet area (cm2) | 97.3 ± 8.4 | 96.7 ± 8.9 | 95.4 ± 13.9 | 97.9 ± 7.5 | |
| Fresh weight (g) | Leaves | 8.9 ± 1.0 | 5.7 ± 1.3 * | 7.3 ± 0.8 * | 5.3 ± 0.7 * |
| Roots | 1.5 ± 0.4 | 1.2 ± 0.8 | 1.5 ± 0.3 | 1.5 ± 0.3 | |
| Whole plants | 18.2 ± 2.1 | 11.9 ± 2.98 * | 15.6 ± 0.9 * | 13.2 ± 1.3 * | |
| Dry weight (g) | Leaves | 1.1 ± 0.1 | 0.5 ± 0.1 * | 0.9 ± 0.1 * | 1.0 ± 0.1 |
| Roots | 0.2 ± 0.1 | 0.1 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.1 | |
| Whole plants | 2.3 ± 0.3 | 1.2 ± 0.3 * | 2.2 ± 0.1 | 2.5 ± 0.2 | |
| Stomatal density (mm−2) | 25.0 ± 3.2 | 25.7 ± 3.6 | 30.7 ± 6.5 * | 53.6 ± 5.9 * | |
| Stomatal index (%) | 32.6 ± 4.4 | 30.5 ± 4.3 | 35.6 ± 4.1 | 37.7 ± 4.2 * | |
| Leaf relative water content (%) | 89.6 ± 10.1 | 88.1 ± 4.2 | 87.9 ± 5.8 | 47.7 ± 0.4 * | |
| Water-use efficiency (g DW kg−1 H2O) | 2.8 ± 0.1 | 3.6 ± 0.1 * | 2.1 ± 0.1 * | 1.7 ± 0.1 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayawardhane, J.; Goyali, J.C.; Zafari, S.; Igamberdiev, A.U. The Response of Cowpea (Vigna unguiculata) Plants to Three Abiotic Stresses Applied with Increasing Intensity: Hypoxia, Salinity, and Water Deficit. Metabolites 2022, 12, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12010038
Jayawardhane J, Goyali JC, Zafari S, Igamberdiev AU. The Response of Cowpea (Vigna unguiculata) Plants to Three Abiotic Stresses Applied with Increasing Intensity: Hypoxia, Salinity, and Water Deficit. Metabolites. 2022; 12(1):38. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12010038
Chicago/Turabian StyleJayawardhane, Jayamini, Juran C. Goyali, Somaieh Zafari, and Abir U. Igamberdiev. 2022. "The Response of Cowpea (Vigna unguiculata) Plants to Three Abiotic Stresses Applied with Increasing Intensity: Hypoxia, Salinity, and Water Deficit" Metabolites 12, no. 1: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12010038