
Molecular Mechanisms of Parathyroid Disorders in Chronic Kidney Disease


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Post-Transcriptional Mechanisms Regulating Parathyroid Hormone Gene Expression in Secondary Hyperparathyroidism
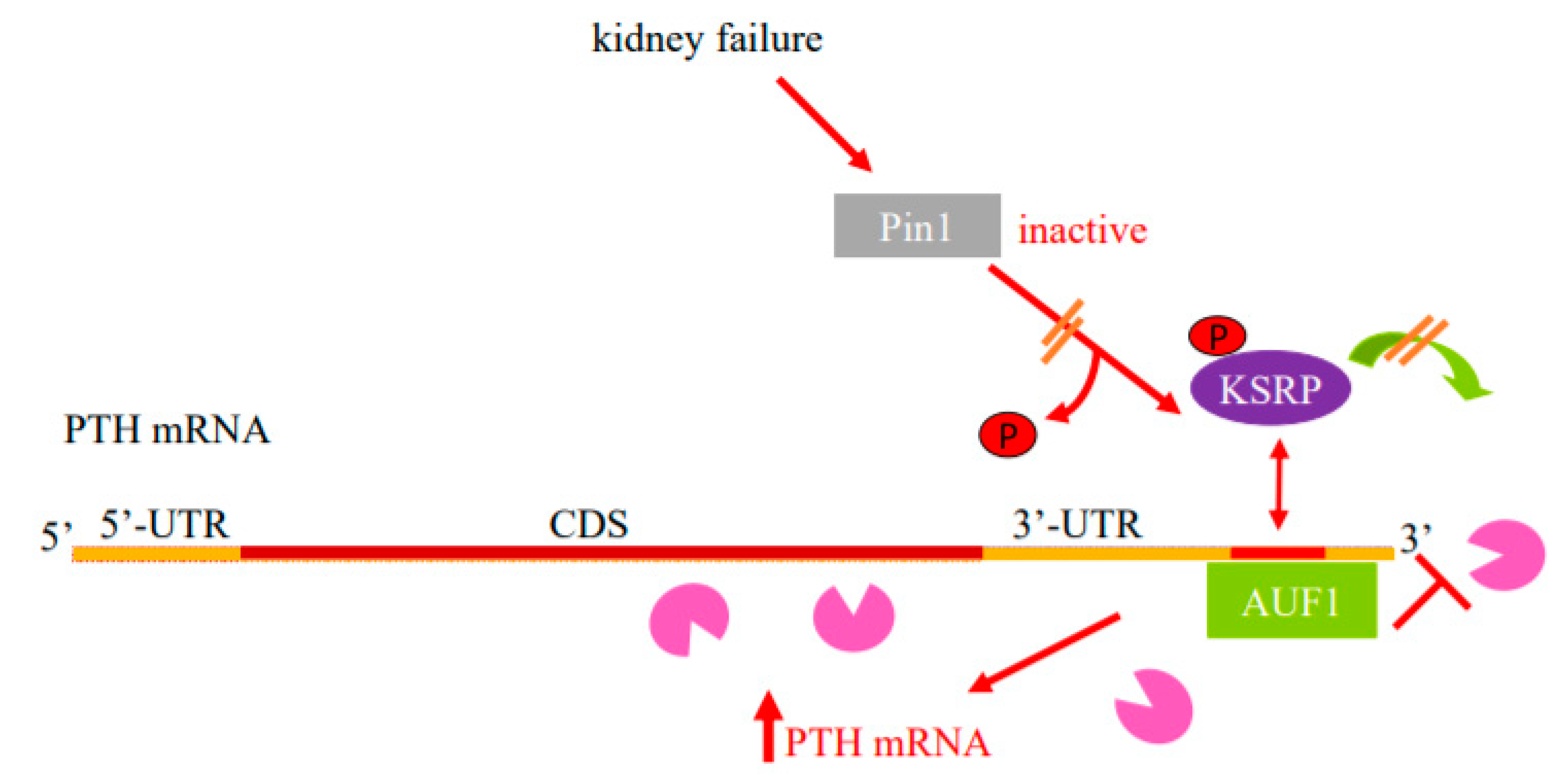
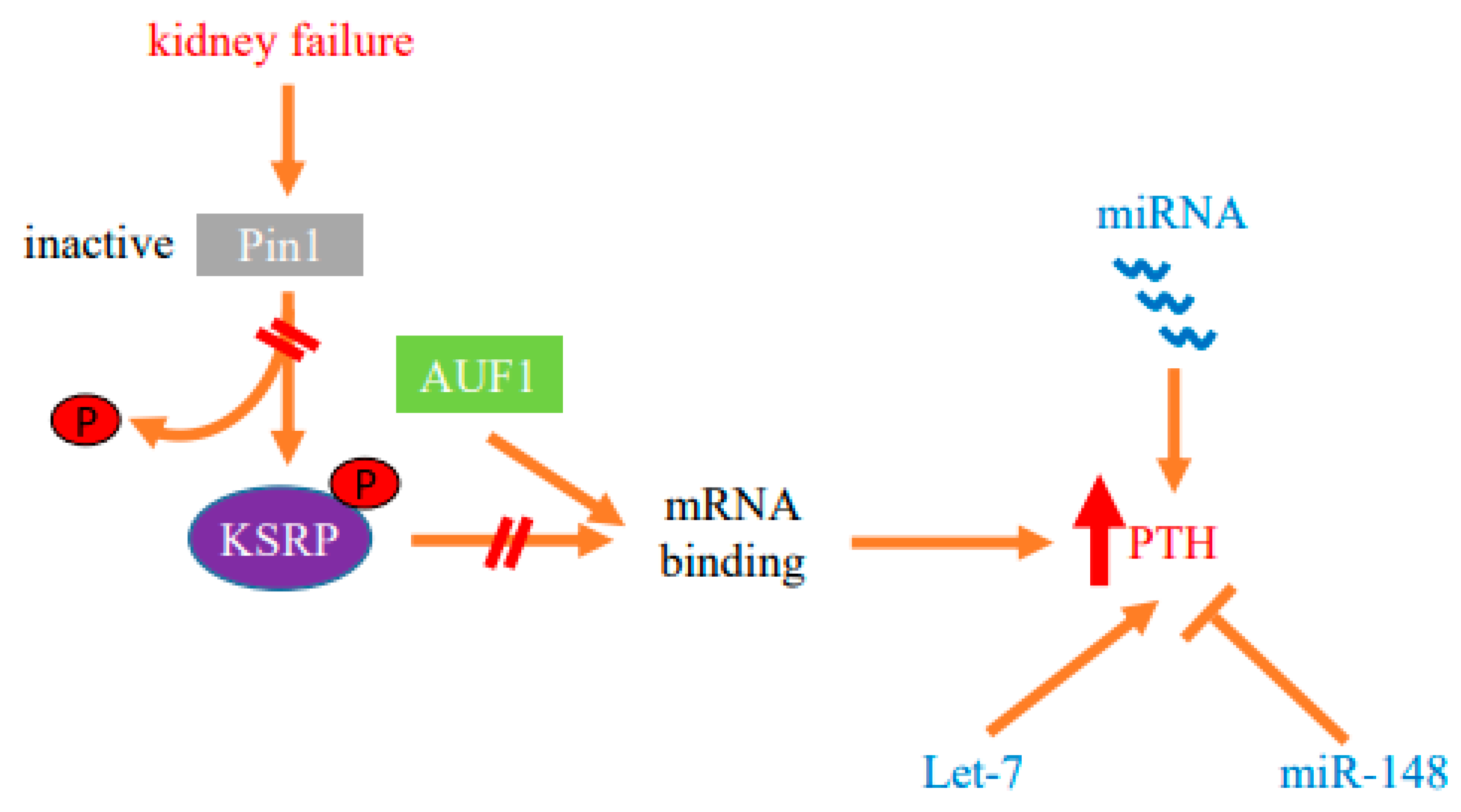
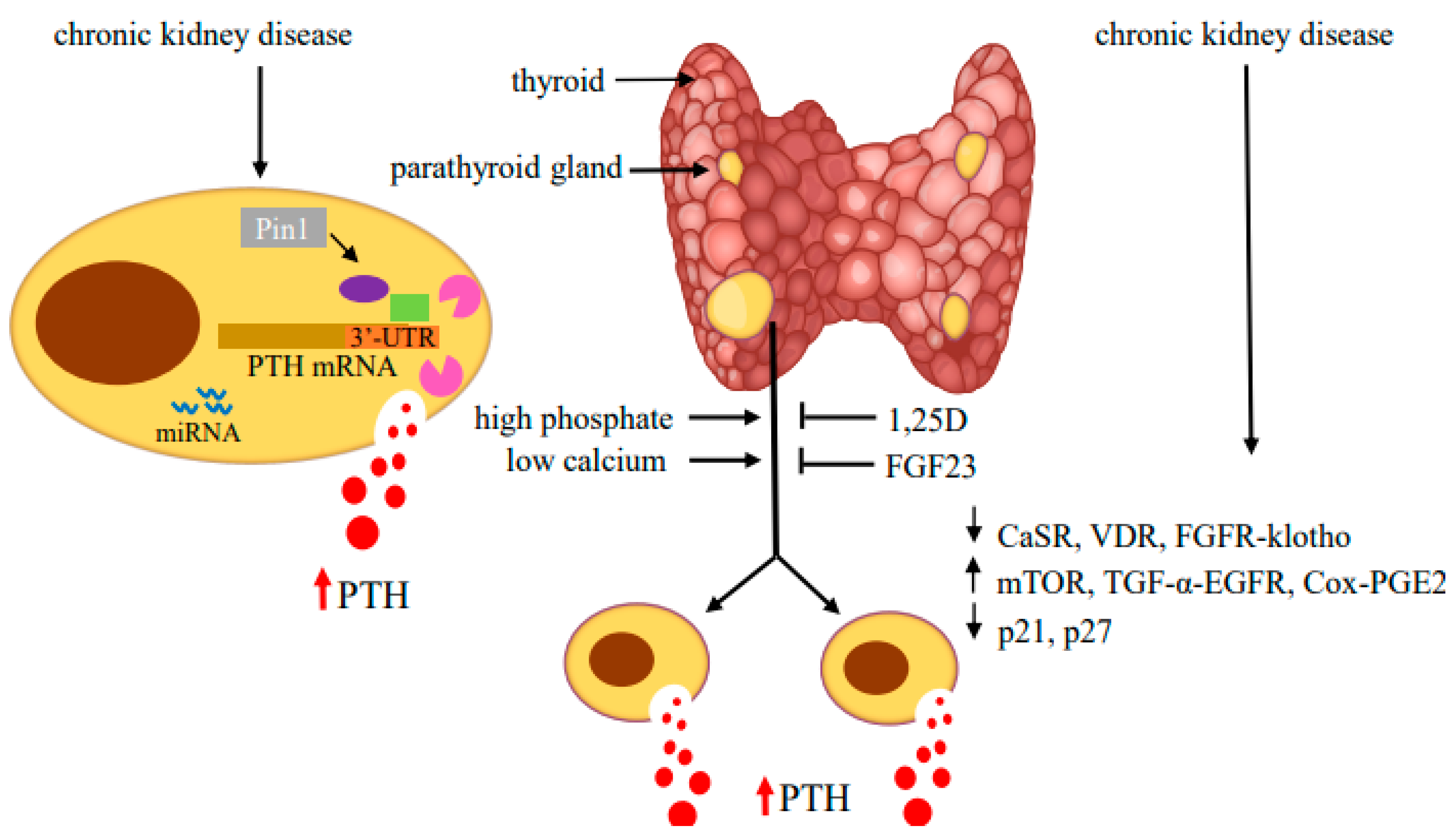
2.1. Regulation of PTH mRNA Stability by Protein–PTH mRNA Interactions and Pin1
2.2. Parathyroid microRNA Ablation in Experimental Secondary Hyperparathyroidism
2.3. Parathyroid miRNA Profiles
2.4. Function of Individual miRNAs in PTH Expression
3. Parathyroid Cell Proliferation in Secondary Hyperparathyroidism
3.1. Cell Cycle Regulation
3.2. TGF-α and EGFR
3.3. The mTORC1 Pathway
3.4. Nuclear Factor-Kappa B
3.5. Cyclooxygenase 2-PGE2
3.6. The Parathyroid Molecular Circadian Clock
4. Summary
Funding
Conflicts of Interest
Abbreviations
| PTH | parathyroid hormone |
| SHP | secondary hyperparathyroidism |
| CKD | chronic kidney disease |
| CaSR | Calcium sensing receptor |
| VDR | vitamin D receptor |
| FGF23 | fibroblast growth factor 23 |
| MiRNA | micro-RNA |
| MiRNome | miRNA transcriptome |
| 3′-UTR | 3′-untranslated region |
| ARE | AU-rich element |
| AUF1 | adenosine-uridine (AU)-rich binding factor 1 |
| KSRP | K-homology splicing regulatory protein |
| CPEB protein | cytoplasmic polyadenylation element binding protein |
| PT-Dicer−/− | parathyroid-specific gene deletion of Dicer |
| EGFR | epidermal growth factor receptor |
| Mtor | mammalian target of rapamycin |
| mTORC | mammalian target of rapamycin complex |
| TGF-α | transforming growth factor α |
| 1:25D | 1,25 dihydroxy-vitamin D |
| NF-kB | nuclear factor kappa B |
| COX | Cyclooxygenase |
| PGE2 | prostaglandin E2 |
| EP2 | PGE2 receptor 2 subtype |
| FGFR | fibroblast growth factor receptor |
| rpS6p−/− | ribosomal protein S6 (rpS6) harboring alanine substitutions at all serine phosphorylation sites |
| MAPK | mitogen-activated protein kinase |
| PRAD1 | parathyroid adenomatosis 1 |
| MEN1 | multiple endocrine neoplasia type 1 |
| AKT | protein kinase B |
References
- Hannan, M.F.; Kallay, E.; Chang, W.; Brandi, M.L.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. Nat. Rev. Endocrinol. 2019, 15, 33–51. [Google Scholar] [CrossRef]
- Silver, J.; Levi, R. Regulation of pth synthesis and secretion relevant to the management of secondary hyperparathyroidism in chronic kidney disease. Kidney Int. Suppl. 2005, 67, S8–S12. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/15882315 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Kumar, R.; Thompson, J.R. The regulation of parathyroid hormone secretion and synthesis. J. Am. Soc. Nephrol. 2011, 22, 216–224. Available online: https://pubmed.ncbi.nlm.nih.gov/21164021 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Hofer, M.A.; Brown, E.M. Extracellular calcium sensing and signalling. Nat. Rev. Mol. Cell Biol. 2003, 4, 530–538. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12838336 (accessed on 20 December 2021). [CrossRef]
- Brown, E.M.; MacLeod, R.J. Extracellular calcium sensing and extracellular calcium signaling. Physiol. Rev. 2001, 81, 239–297. [Google Scholar] [CrossRef]
- Drüeke, T.B. Modulation and action of the calcium-sensing receptor. Nephrol. Dial. Transplant. 2004, 19, v20–v26. [Google Scholar] [CrossRef] [Green Version]
- Slatopolsky, E.; Caglar, S.; Pennell, J.P.; Taggart, D.D.; Canterbury, J.M.; Reiss, E.; Bricker, N.S. On the pathogenesis of hyperparathyroidism in chronic experimental renal insufficiency in the dog. J. Clin. Investig. 1971, 50, 492–499. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/5545116 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Kilav, R.; Silver, J.; Naveh-Many, T. Parathyroid hormone gene expression in hypophosphatemic rats. J. Clin. Investig. 1995, 96, 327–333. [Google Scholar] [CrossRef]
- Ben Dov, I.Z.; Galitzer, H.; Lavi-Moshayoff, V.; Goetz, R.; Kuro-o, M.; Mohammadi, M.; Sirkis, R.; Naveh-Many, T.; Silver, J. The parathyroid is a target organ for fgf23 in rats. J. Clin. Investig. 2007, 117, 4003–4008. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17992255 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Naveh-Many, T.; Silver, J. The pas de trois of vitamin d, fgf23, and pth. J. Am. Soc. Nephrol. 2016, 28, 393–395. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/27807212 (accessed on 20 December 2021). [CrossRef]
- Centeno, P.P.; Herberger, A.; Mun, H.-C.; Tu, C.; Nemeth, E.F.; Chang, W.; Conigrave, A.D.; Ward, D.T. Phosphate acts directly on the calcium-sensing receptor to stimulate parathyroid hormone secretion. Nat. Commun. 2019, 10, 4693. [Google Scholar] [CrossRef]
- Silver, J.; Naveh-Many, T.; Mayer, H.; Schmelzer, H.J.; Popovtzer, M.M. Regulation by vitamin d metabolites of parathyroid hormone gene transcription in vivo in the rat. J. Clin. Investig. 1986, 78, 1296–1301. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.; Marks, R.; Naveh-Many, T. Regulation of parathyroid hormone and calcitonin gene expression by l,25-dihydroxyvitamin d3 and calcium. In Calcium Regulation and Bone Metabolism: Basic and Clinical Aspects; Cohn, D.V., Glorieux, F.H., Martin, T.J., Eds.; Excerpta Medica: Amsterdam, The Netherlands, 1990; pp. 140–145. [Google Scholar]
- Pike, J.W.; Meyer, M.B.; Watanuki, M.; Kim, S.; Zella, L.A.; Fretz, J.A.; Yamazaki, M.; Shev, N.K. Perspectives on mechanisms of gene regulation by 1,25-dihydroxyvitamin d3 and its receptor. J. Steroid Biochem. Mol. Biol. 2007, 103, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.; Ashok, S.; Koszewski, N.J. Vitamin d receptor interactions with the rat parathyroid hormone gene: Synergistic effects between two negative vitamin d response elements. J. Bone Miner. Res. 1999, 14, 1828–1837. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/0010571682 (accessed on 20 December 2021). [CrossRef]
- Krajisnik, T.; Bjorklund, P.; Marsell, R.; Ljunggren, O.; Akerstrom, G.; Jonsson, K.B.; Westin, G.; Larss, T.E.on. Fibroblast growth factor-23 regulates parathyroid hormone and 1alpha-hydroxylase expression in cultured bovine parathyroid cells. J. Endocrinol. 2007, 195, 125–131. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17911404 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Naveh-Many, T.; Rahamimov, R.; Livni, N.; Silver, J. Parathyroid cell proliferation in normal and chronic renal failure rats: The effects of calcium, phosphate and vitamin d. J. Clin. Investig. 1995, 96, 1786–1793. [Google Scholar] [CrossRef]
- Levi, R.; Ben Dov, I.Z.; Lavi-Moshayoff, V.; Dinur, M.; Martin, D.; Naveh-Many, T.; Silve, J. Increased parathyroid hormone gene expression in secondary hyperparathyroidism of experimental uremia is reversed by calcimimetics: Correlation with posttranslational modification of the trans acting factor auf1. J. Am. Soc. Nephrol. 2006, 17, 107–112. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/16291838 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Goodman, W.G.; Quarles, L.D. Development and progression of secondary hyperparathyroidism in chronic kidney disease: Lessons from molecular genetics. Kidney Int. 2008, 74, 276–288. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17568787 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Lewin, E.; Huan, J.; Olgaard, K. Parathyroid growth and suppression in renal failure. Semin. Dial. 2006, 19, 238–245. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/16689976 (accessed on 20 December 2021). [CrossRef]
- Brown, A.J.; Dusso, A.; Lopez-Hilker, S.; Lewis-Finch, J.; Grooms, P.; Slatopolsky, E. 1,25-(oh)2d receptors are decreased in parathyroid glands from chronically uremic dogs. Kidney Int. 1989, 35, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.; Kilav, R.; Naveh-Many, T. Mechanisms of secondary hyperparathyroidism. Am. J. Physiol. Ren. Physiol. 2002, 283, F367–F376. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12167586 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Moallem, E.; Silver, J.; Kilav, R.; Naveh-Many, T. Rna protein binding and post-transcriptional regulation of pth gene expression by calcium and phosphate. J. Biol. Chem. 1998, 273, 5253–5259. [Google Scholar] [CrossRef] [Green Version]
- Sela-Brown, A.; Silver, J.; Brewer, G.; Naveh-Many, T. Identification of auf1 as a parathyroid hormone mrna 3′-untranslated region binding protein that determines parathyroid hormone mrna stability. J. Biol. Chem. 2000, 275, 7424–7429. [Google Scholar] [CrossRef] [Green Version]
- Nechama, M.; Ben Dov, I.Z.; Briata, P.; Gherzi, R.; Naveh-Many, T. The mrna decay promoting factor k-homology splicing regulator protein post-transcriptionally determines parathyroid hormone mrna levels. FASEB J. 2008, 22, 3458–3468. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/18583400 (accessed on 20 December 2021). [CrossRef]
- Kilav-Levin, R.; Hassan, A.; Nechama, M.; Shilo, V.; Silver, J.; Ben-Dov, I.Z.; Naveh-Many, T. Post-transcriptional mechanisms regulating parathyroid hormone gene expression in secondary hyperparathyroidism. FEBS J. 2020, 287, 2903–2913. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.M.; Brewer, G. The search for trans-acting factors controlling messenger rna decay. Prog. Nucleic Acid Res. Mol. Biol. 1999, 62, 257–291. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/0009932457 (accessed on 20 December 2021).
- Briata, P.; Chen, C.Y.; Giovarelli, M.; Pasero, M.; Trabucchi, M.; Ramos, A.; Gherzi, R. Ksrp, many functions for a single protein. Front. Biosci. (Landmark Ed.) 2011, 16, 1787–1796. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/21196264 (accessed on 20 December 2021). [CrossRef]
- Barreau, C.; Paillard, L.; Osborne, H.B. Au-rich elements and associated factors: Are there unifying principles? Nucleic Acids Res. 2006, 33, 7138–7150. [Google Scholar] [CrossRef]
- Garneau, N.L.; Wilusz, J.; Wilusz, C.J. The highways and byways of mrna decay. Nat. Rev. Mol. Cell Biol. 2007, 8, 113–126. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17245413 (accessed on 20 December 2021). [CrossRef]
- Peng, S.S.; Chen, C.Y.; Xu, N.; Shyu, A.B. Rna stabilization by the au-rich element binding protein, hur, an elav protein. EMBO J. 1998, 17, 3461–3470. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/9628881 (accessed on 20 December 2021). [CrossRef]
- Wickens, M.; Anderson, P.; Jakson, R.J. Life and death in the cytoplasm: Messages from the 3’ end. Curr. Opin. Genet. Dev. 1996, 7, 220–232. [Google Scholar] [CrossRef]
- Chen, C.Y.; Shyu, A.B. Au-rich elements: Characterization and importance in mrna degradation. Trends Biochem. Sci. 1995, 20, 465–470. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8578590 (accessed on 20 December 2021). [CrossRef]
- Zhang, W.; Wagner, B.J.; Ehrenman, K.; Schaefer, A.W.; DeMaria, C.T.; Crater, D.; DeHaven, K.; Long, L.; Brewer, G. Purification, characterization, and cdna cloning of an au-rich element rna-binding protein, auf1. Mol. Cell Biol. 1993, 13, 7652–7665. [Google Scholar]
- Gherzi, R.; Trabucchi, M.; Ponassi, M.; Ruggiero, T.; Corte, G.; Moroni, C.; Chen, C.Y.; Khabar, K.S.; Andersen, J.S.; Briata, P. The rna-binding protein ksrp promotes decay of beta-catenin mrna and is inactivated by pi3k-akt signaling. PLoS Biol. 2006, 5, e5. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17177604 (accessed on 20 December 2021). [CrossRef]
- DeMaria, C.T.; Brewer, G. Auf1 binding affinity to a+u-rich elements correlates with rapid mrna degradation. J. Biol. Chem. 1996, 271, 12179–12184. [Google Scholar] [CrossRef] [Green Version]
- Kiledjian, M.; DeMaria, C.T.; Brewer, G.; Novick, K. Identification of auf1 (heterogeneous nclear ribonucleoprotein d) as a component of the a-globin mrna stability complex. Mol. Cell Biol. 1997, 17, 4870–4876. [Google Scholar] [CrossRef] [Green Version]
- Gherzi, R.; Lee, K.Y.; Briata, P.; Wegmuller, D.; Moroni, C.; Karin, M.; Chen, C.Y. A kh domain rna binding protein, ksrp, promotes are-directed mrna turnover by recruiting the degradation machinery. Mol. Cell 2004, 14, 571–583. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/15175153 (accessed on 20 December 2021). [CrossRef]
- Esnault, S.; Shen, Z.J.; Whitesel, E.; Malter, J.S. The peptidyl-prolyl isomerase pin1 regulates granulocyte-macrophage colony-stimulating factor mrna stability in t lymphocytes. J. Immunol. 2006, 177, 6999–7006. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17082615 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Bell, O.; Gaberman, E.; Kilav, R.; Levi, R.; Cox, K.B.; Molkentin, J.D.; Silver, J.; Naveh-Many, T. The protein phosphatase calcineurin determines basal parathyroid hormone gene expression. Mol. Endocrinol. 2005, 19, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di¡az-Moreno, I.; Hollingworth, D.; Frenkiel, T.A.; Kelly, G.; Martin, S.; Howell, S.; García-Mayoral, M.A.; Gherzi, R.; Briata, P.; Ramos, A. Phosphorylation-mediated unfolding of a kh domain regulates ksrp localization via 14-3-3 binding. Nat. Struct. Mol. Biol. 2009, 16, 238–246. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC2858377/ (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Briata, P.; Forcales, S.V.; Ponassi, M.; Corte, G.; Chen, C.Y.; Karin, M.; Puri, P.L.; Gherzi, R. P38-dependent phosphorylation of the mrna decay-promoting factor ksrp controls the stability of select myogenic transcripts. Mol. Cell. 2005, 20, 891–903. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/16364914 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Wilson, G.M.; Lu, J.; Sutphen, K.; Suarez, Y.; Sinha, S.; Brewer, B.; Villanueva-Feliciano, E.C.; Ysla, R.M.; Charles, S.; Brewe, G. Phosphorylation of p40auf1 regulates binding to a + u-rich mrna-destabilizing elements and protein-induced changes in ribonucleoprotein structure. J. Biol. Chem. 2003, 278, 33039–33048. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12819194 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Lee, E.K.; Gorospe, M. Minireview: Posttranscriptional regulation of the insulin and insulin-like growth factor systems. Endocrinology 2010, 151, 1403–1408. Available online: https://pubmed.ncbi.nlm.nih.gov/20032049 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Ing, N.H. Steroid hormones regulate gene expression posttranscriptionally by altering the stabilities of messenger rnas. Biol. Reprod. 2005, 72, 1290–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.K.; Kash, J.C.; Peegel, H.; Meno, K.M.J. Post-transcriptional regulation of luteinizing hormone receptor mrna in the ovary by a novel mrna-binding protein. J. Biol. Chem. 2002, 277, 21468–21473. Available online: http://www.jbc.org/content/277/24/21468.abstract (accessed on 20 December 2021). [CrossRef] [Green Version]
- Menon, K.M.J.; Menon, B.; Gulappa, T. Regulation of luteinizing hormone receptor mrna expression in the ovary: The role of mir-122. Vitam. Horm. 2018, 107, 67–87. Available online: https://pubmed.ncbi.nlm.nih.gov/29544643 (accessed on 20 December 2021). [CrossRef]
- Shilo, V.; Silver, J.; Naveh-Many, T. Micro-rnas in the parathyroid: A new portal in understanding secondary hyperparathyroidism. Curr. Opin. Nephrol. Hypertens. 2016, 25, 271–277. [Google Scholar] [CrossRef]
- Kilav, R.; Silver, J.; Naveh-Man, T. A conserved cis-acting element in the parathyroid hormone 3′-untranslated region is sufficient for regulation of rna stability by calcium and phosphate. J. Biol. Chem. 2001, 276, 8727–8733. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.P.; Zhou, X.Z. The prolyl isomerase pin1: A pivotal new twist in phosphorylation signalling and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 904–916. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17878917 (accessed on 20 December 2021). [CrossRef]
- Lu, K.P.; Hanes, S.D.; Hunter, T. A human peptidyl-prolyl isomerase essential for regulation of mitosis. Nature 1996, 380, 544–547. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/8606777 (accessed on 20 December 2021).
- Thapar, R. Roles of prolyl isomerases in rna-mediated gene expression. Biomolecules 2015, 5, 974–999. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/25992900 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Shen, Z.J.; Malter, J.S. Regulation of au-rich element rna binding proteins by phosphorylation and the prolyl isomerase pin1. Biomolecules 2015, 5, 412–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nechama, M.; Uchida, T.; Yosef-Levi, I.M.; Silver, J.; Naveh-Many, T. The peptidyl-prolyl isomerase pin1 determines parathyroid hormone mrna levels and stability in rat models of secondary hyperparathyroidism. J. Clin. Investig. 2009, 119, 3102–3114. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.J.; Esnault, S.; Rosenthal, L.A.; Szakaly, R.J.; Sorkness, R.L.; Westmark, P.R.; Sandor, M.; Malter, J.S. Pin1 regulates tgf-beta1 production by activated human and murine eosinophils and contributes to allergic lung fibrosis. J. Clin. Investig. 2008, 118, 479–490. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/18188456 (accessed on 20 December 2021).
- Shen, Z.J.; Esnault, S.; Malter, J.S. The peptidyl-prolyl isomerase pin1 regulates the stability of granulocyte-macrophage colony-stimulating factor mrna in activated eosinophils. Nat. Immunol. 2005, 6, 1280–1287. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/16273101 (accessed on 20 December 2021). [CrossRef]
- Nechama, M.; Lin, C.L.; Richter, J.D. An unusual two-step control of cpeb destruction by pin1. Mol. Cell Biol. 2013, 33, 48–58. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/23090969 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Schelhorn, C.; Martín-Malpartida, P.; Suñol, D.; Macias, M.J. Structural analysis of the pin1-cpeb1 interaction and its potential role in cpeb1 degradation. Sci. Rep. 2015, 5, 14990. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/srep14990#supplementary-information (accessed on 20 December 2021). [CrossRef] [Green Version]
- Kumar, R. Pin1 regulates parathyroid hormone mrna stability. J. Clin. Investig. 2009, 119, 2887–2891. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/19770518 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Oster, M.; Reyer, H.; Gerlinger, C.; Trakooljul, N.; Siengdee, P.; Keiler, J.; Ponsuksili, S.; Wolf, P.; Wimme, K. Mrna profiles of porcine parathyroid glands following variable phosphorus supplies throughout fetal and postnatal life. Biomedicines 2021, 9, 454. Available online: https://pubmed.ncbi.nlm.nih.gov/33922173 (accessed on 20 December 2021). [CrossRef]
- Santamaria, I.; Alvarez-Hernandez, D.; Jofre, R.; Polo, J.R.; Menarguez, J.; Cannata-Andi, J.B. Progression of secondary hyperparathyroidism involves deregulation of genes related to DNA and rna stability. Kidney Int. 2005, 67, 2267–2279. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/15882268 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Balenga, N.; Azimzadeh, P.; Hogue, J.A.; Staats, P.N.; Shi, Y.; Koh, J.; Dressman, H.; Olson, J.A. Orphan adhesion gpcr gpr64/adgrg2 is overexpressed in parathyroid tumors and attenuates calcium-sensing receptor-mediated signaling. J. Bone Miner. Res. 2017, 32, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, L.-L.; Ding, F.-X.; Cao, P.; Qi, Y.-Y.; Wan, J. Pin1 and secondary hyperparathyroidism of chronic kidney disease: Gene polymorphisms and protein levels. Ren. Fail. 2017, 39, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Micrornas: Target recognition and regulatory functions. Cell 2009, 136, 215–233. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/19167326 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Hafner, M.; Landgraf, P.; Ludwig, J.; Rice, A.; Ojo, T.; Lin, C.; Holoch, D.; Lim, C.; Tuschl, T. Identification of micrornas and other small regulatory rnas using cdna library sequencing. Methods 2008, 44, 3–12. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/18158127 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Ahn, C.; Han, J.J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; et al. The nuclear rnase iii drosha initiates microrna processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef]
- Chen, J.F.; Murchison, E.P.; Tang, R.; Callis, T.E.; Tatsuguchi, M.; Deng, Z.; Rojas, M.; Hammond, S.M.; Schneider, M.D.; Selzman, C.H.; et al. Targeted deletion of dicer in the heart leads to dilated cardiomyopathy and heart failure. Proc. Natl. Acad. Sci. USA 2008, 105, 2111–2116. [Google Scholar] [CrossRef] [Green Version]
- Melkman-Zehavi, T.; Oren, R.; Kredo-Russo, S.; Shapira, T.; Mandelbaum, A.D.; Rivkin, N.; Nir, T.; Lennox, K.A.; Behlke, M.A.; Dor, Y.; et al. Mirnas control insulin content in pancreatic beta-cells via downregulation of transcriptional repressors. EMBO J. 2011, 30, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Harfe, B.D.; McManus, M.T.; Mansfield, J.H.; Hornstein, E.; Tabin, C.J. The rnaseiii enzyme dicer is required for morphogenesis but not patterning of the vertebrate limb. Proc. Natl. Acad. Sci. USA 2005, 102, 10898–10903. [Google Scholar] [CrossRef] [Green Version]
- Shilo, V.; Dov, I.Z.B.; Nechama, M.; Silver, J.; Naveh-Many, T. Parathyroid-specific deletion of dicer-dependent micrornas abrogates the response of the parathyroid to acute and chronic hypocalcemia and uremia. FASEB J. 2015, 29, 3964–3976. [Google Scholar] [CrossRef] [Green Version]
- Aurilia, C.; Donati, S.; Palmini, G.; Miglietta, F.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. Are non-coding rnas useful biomarkers in parathyroid tumorigenesis? Int. J. Mol. Sci. 2021, 22, 10465. [Google Scholar] [CrossRef] [PubMed]
- Verdelli, C.; Forno, I.; Vaira, V.; Corbetta, S. Epigenetic alterations in human parathyroid tumors. Endocrine 2015, 49, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, V.; Muscarella, L.A.; Verdelli, C.; Corbetta, S. Alterations of DNA methylation in parathyroid tumors. Mol. Cell Endocrinol. 2018, 469, 60–69. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/28501573 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Verdelli, C.; Corbetta, S. Epigenetic alterations in parathyroid cancers. Int. J. Mol. Sci. 2017, 18, 310. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/28157158 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Silva-Figueroa, A.M.; Perrier, N.D. Epigenetic processes in sporadic parathyroid neoplasms. Mol. Cell Endocrinol. 2018, 469, 54–59. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/28400272 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Mizamtsidi, M.; Nastos, K.; Palazzo, F.; Constantinides, V.; Dina, R.; Farenden, M.; Mastorakos, G.; Vassiliou, I.; Gazouli, M. Association between hsa-mir-30e polymorphisms and sporadic primary hyperparathyroidism risk. In Vivo 2019, 33, 1263–1269. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/31280217 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Mizamtsidi, M.; Nastos, C.; Mastorakos, G.; Dina, R.; Vassiliou, I.; Gazouli, M.; Palazzo, F. Diagnosis, management, histology and genetics of sporadic primary hyperparathyroidism: Old knowledge with new tricks. Endocr. Connect. 2018, 7, R56–R68. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/29330338 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Vaira, V.; Elli, F.; Forno, I.; Guarnieri, V.; Verdelli, C.; Ferrero, S.; Scillitani, A.; Vicentini, L.; Cetani, F.; Mantovani, G.; et al. The microrna cluster c19mc is deregulated in parathyroid tumours. J. Mol. Endocrinol. 2012, 49, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Corbetta, S.; Vaira, V.; Guarnieri, V.; Scillitani, A.; Eller-Vainicher, C.; Ferrero, S.; Vicentini, L.; Chiodini, I.; Bisceglia, M.; Beck-Peccoz, P.; et al. Differential expression of micrornas in human parathyroid carcinomas compared with normal parathyroid tissue. Endocr. Relat. Cancer 2010, 17, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Shilo, V.; Levi, I.M.; Abel, R.; Mihailović, A.; Wasserman, G.; Naveh-Many, T.; Ben-Dov, I.Z. Let-7 and microrna-148 regulate parathyroid hormone levels in secondary hyperparathyroidism. J. Am. Soc. Nephrol. 2017, 28, 2353–2363. Available online: http://jasn.asnjournals.org/content/early/2017/03/14/ASN.2016050585.abstract (accessed on 20 December 2021). [CrossRef] [Green Version]
- Esau, C.C. Inhibition of microrna with antisense oligonucleotides. Methods 2008, 44, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Palnitkar, S.; Parfitt, A.M. Parathyroid cell proliferation in the rat: Effect of age and of phosphate administration and recovery. Endocrinology 1996, 137, 4558–4562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parfitt, A.M. Parathyroid growth, normal and abnormal. In The Parathyroids; Bilezikian, J.P., Levine, M.A., Marcus, R., Eds.; Raven Press, Ltd.: New York, NY, USA, 1994; pp. 373–405. [Google Scholar]
- Tominaga, Y.; Kohara, S.; Namii, Y.; Nagasaka, T.; Haba, T.; Uchida, K.; Numano, M.; Tanaka, Y.; Takagi, H. Clonal analysis of nodular parathyroid hyperplasia in renal hyperparathyroidism. World J. Surg. 1996, 20, 744–750, discussion 50–52. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.; Brown, M.F.; Urena, P.; Gaz, R.D.; Sarfati, E.; Drueke, T.B. Monoclonality of parathyroid tumors in chronic renal failure and in primary parathyroid hyperplasia. J. Clin. Investig. 1995, 95, 2047–2053. [Google Scholar] [CrossRef]
- Drueke, T.B. Cell biology of parathyroid gland hyperplasia in chronic renal failure. J. Am. Soc. Nephrol. 2000, 11, 1141–1152. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/0010820180 (accessed on 20 December 2021). [CrossRef]
- Denda, M.; Finch, J.; Slatopolsky, E. Phosphorus accelerates the development of parathyroid hyperplasia and secondary hyperparathyroidism in rats with renal failure. Am. J. Kidney Dis. 1996, 28, 596–602. [Google Scholar] [CrossRef]
- Yokozawa, T.; Zheng, P.D.; Oura, H.; Koizumi, F. Animal model of adenine-induced chronic renal failure in rats. Nephron 1986, 44, 230–234. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/3785486 (accessed on 20 December 2021). [CrossRef]
- Ceballos-Picot, I.; Saha, A.; Arora, N.; Kapoor, K.; Kaur, M.; Dhull, R.S.; Goyal, S. Adenine phosphoribosyltransferase deficiency due to novel mutation. Kidney Int. Rep. 2018, 4, 624–628. Available online: https://pubmed.ncbi.nlm.nih.gov/30993240 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Jia, T.; Olauson, H.; Lindberg, K.; Amin, R.; Edvardsson, K.; Lindholm, B.; Andersson, G.; Wernerson, A.; Sabbagh, Y.; Schiavi, S.; et al. A novel model of adenine-induced tubulointerstitial nephropathy in mice. BMC Nephrol. 2013, 14, 116. [Google Scholar] [CrossRef] [Green Version]
- Ritter, C.S.; Finch, J.L.; Slatopolsky, E.A.; Brown, A.J. Parathyroid hyperplasia in uremic rats precedes down-regulation of the calcium receptor. Kidney Int. 2001, 60, 1737–1744. [Google Scholar] [CrossRef] [Green Version]
- Galitzer, H.; Dov, I.Z.B.; Silver, J.; Naveh-Many, T. Parathyroid cell resistance to fibroblast growth factor 23 in secondary hyperparathyroidism of chronic kidney disease. Kidney Int. 2010, 77, 211–218. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/20016468 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Komaba, H.; Goto, S.; Fujii, H.; Hamada, Y.; Kobayashi, A.; Shibuya, K.; Tominaga, Y.; Otsuki, N.; Nibu, K.I.; Nakagawa, K.; et al. Depressed expression of klotho and fgf receptor 1 in hyperplastic parathyroid glands from uremic patients. Kidney Int. 2010, 77, 232–238. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/19890272 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Canalejo, R.; Canalejo, A.; Martinez-Moreno, J.M.; Rodriguez-Ortiz, M.E.; Estepa, J.C.; Mendoza, F.J.; Munoz-Castaneda, J.R.; Shalhoub, V.; Almaden, Y.; Rodriguez, M. Fgf23 fails to inhibit uremic parathyroid glands. J. Am. Soc. Nephrol. 2010, 21, 1125–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, E.; Garfia, B.; Recio, F.L.; Rodriguez, M.; Olgaard, K. Persistent downregulation of calcium-sensing receptor mrna in rat parathyroids when severe secondary hyperparathyroidism is reversed by an isogenic kidney transplantation. J. Am. Soc. Nephrol. 2002, 13, 2110–2116. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12138143 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Garfia, B.; Canadillas, S.; Canalejo, A.; Luque, F.; Siendones, E.; Quesada, M.; Almaden, Y.; Aguilera-Tejero, E.; Rodriguez, M. Regulation of parathyroid vitamin d receptor expression by extracellular calcium. J. Am. Soc. Nephrol. 2002, 13, 2945–2952. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12444213 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Healy, K.D.; Vanhooke, J.L.; Prahl, J.M.; DeLuca, H.F. Parathyroid hormone decreases renal vitamin d receptor expression in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 4724–4728. Available online: https://pubmed.ncbi.nlm.nih.gov/15769857 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Naveh-Many, T.; Volovelsky, O. Parathyroid cell proliferation in secondary hyperparathyroidism of chronic kidney disease. Int. J. Mol. Sci. 2020, 21, 4332. Available online: https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/21/12/4332 (accessed on 20 December 2021). [CrossRef]
- Rodríguez, M.; Goodman, W.G.; Liakopoulos, V.; Messa, P.; Wiecek, A.; Cunningham, J. The use of calcimimetics for the treatment of secondary hyperparathyroidism: A 10 year evidence review. Semin. Dial. 2015, 28, 497–507. [Google Scholar] [CrossRef]
- Canaff, L.; Hendy, G.N. Human calcium-sensing receptor gene. Vitamin d response elements in promoters p1 and p2 confer transcriptional responsiveness to 1,25-dihydroxyvitamin d. J. Biol. Chem. 2002, 277, 30337–30350. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12036954 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Denda, M.; Finch, J.; Brown, A.J.; Nishii, Y.; Kubodera, N.; Slatopolsky, E. 1,25-dihydroxyvitamin d3 and 22-oxacalcitriol prevent the decrease in vitamin d receptor content in the parathyroid glands of uremic rats. Kidney Int. 1996, 50, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Sakai, M.; Tokunaga, S.; Kawai, M.; Murai, M.; Kobayashi, M.; Kitayama, T.; Saeki, S.; Kawata, T. Evocalcet prevents ectopic calcification and parathyroid hyperplasia in rats with secondary hyperparathyroidism. PLoS ONE 2020, 15, e0232428. Available online: https://pubmed.ncbi.nlm.nih.gov/32343734 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Meola, M.; Petrucci, I.; Barsotti, G. Long-term treatment with cinacalcet and conventional therapy reduces parathyroid hyperplasia in severe secondary hyperparathyroidism. Nephrol. Dial. Transplant. 2009, 24, 982–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, F.J.; Lopez, I.; Canalejo, R.; Almaden, Y.; Martin, D.; Aguilera-Tejero, E.; Rodriguez, M. Direct upregulation of parathyroid calcium-sensing receptor and vitamin d receptor by calcimimetics in uremic rats. Am. J. Physiol. Ren. Physiol. 2009, 296, F605–F613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nechama, M.; Dov, I.Z.B.; Silver, J.; Naveh-Many, T. Regulation of pth mrna stability by the calcimimetic r568 and the phosphorus binder lanthanum carbonate in ckd. Am. J. Physiol. Ren. Physiol. 2009, 296, F795–F800. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/19129257 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Haven, C.J.; Howell, V.M.; Eilers, P.H.; Dunne, R.; Takahashi, M.; van Puijenbroek, M.; Furge, K.; Kievit, J.; Tan, M.H.; Fleuren, G.J.; et al. Gene expression of parathyroid tumors: Molecular subclassification and identification of the potential malignant phenotype. Cancer Res. 2004, 64, 7405–7411. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/15492263 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Libutti, S.K.; Crabtree, J.S.; Lorang, D.; Burns, A.L.; Mazzanti, C.; Hewitt, S.M.; O’Connor, S.; Ward, J.M.; Emmert-Buck, M.R.; Remaley, A.; et al. Parathyroid gland-specific deletion of the mouse men1 gene results in parathyroid neoplasia and hypercalcemic hyperparathyroidism. Cancer Res. 2003, 63, 8022–8028. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/14633735 (accessed on 20 December 2021).
- Mulligan, L.M.; Marsh, D.J.; Robinson, B.G.; Schuffenecker, I.; Zedenius, J.; Lips, C.J.; Gagel, R.F.; Takai, S.I.; Noll, W.W.; Fink, M.; et al. Genotype-phenotype correlation in multiple endocrine neoplasia type 2: Report of the international ret mutation consortium. J. Intern. Med. 1995, 238, 343–346. [Google Scholar] [CrossRef]
- Hsi, E.D.; Zukerberg, L.R.; Yang, W.I.; Arnold, A. Cyclin d1/prad1 expression in parathyroid adenomas: An immunohistochemical study. J. Clin. Endocrinol. Metab. 1996, 81, 1736–1739. [Google Scholar]
- Motokura, T.; Arnold, A. Prad1/cyclin d1 proto-oncogene: Genomic organization, 5′ DNA sequence, and sequence of a tumor-specific rearrangement breakpoint. Genes Chromosom.Cancer 1993, 7, 89–95. [Google Scholar] [CrossRef]
- Arnold, A. Molecular basis of primary hyperparathyroidism. In The Parathyroids; Bilezikian, J.P., Levine, M.A., Marcus, R., Eds.; Raven Press, Ltd.: New York, NY, USA, 1994; pp. 407–421. [Google Scholar]
- Brewer, K.; Costa-Guda, J.; Arnold, A. Molecular genetic insights into sporadic primary hyperparathyroidism. Endocr. Relat. Cancer 2019, 26, R53–R72. [Google Scholar] [CrossRef]
- Imanishi, Y.; Hosokawa, Y.; Yoshimoto, K.; Schipani, E.; Mallya, S.; Papanikolaou, A.; Kifor, O.; Tokura, T.; Sablosky, M.; Ledgard, F.; et al. Primary hyperparathyroidism caused by parathyroid-targeted overexpression of cyclin d1 in transgenic mice. J. Clin. Investig. 2001, 107, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Hirama, T.; Koeffler, H.P. Role of the cyclin-dependent kinase inhibitors in the development of cancer. Blood 1995, 86, 841–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tominaga, Y.; Tsuzuki, T.; Uchida, K.; Haba, T.; Otsuka, S.; Ichimori, T.; Yamada, K.; Numano, M.; Tanaka, Y.; Takagi, H. Expression of prad1/cyclin d1, retinoblastoma gene products, and ki67 in parathyroid hyperplasia caused by chronic renal failure versus primary adenoma. Kidney Int. 1999, 55, 1375–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graña, X.; Reddy, E.P. Cell cycle control in mammalian cells: Role of cyclins, cyclin dependent kinases (cdks), growth suppressor genes and cyclin-dependent kinase inhibitors (ckis). Oncogene 1995, 11, 211–219. [Google Scholar]
- Dusso, A.; Pavlopoulos, T.; Naumovich, L.; Lu, Y.; Finch, J.; Brown, A.J.; Morrissey, J.; Slatopolsky, E. P21waf1 and transforming growth factor-alpha mediate dietary posphate regulation of parathyroid cell growth. Kidney Int. 2001, 59, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Tokumoto, M.; Tsuruya, K.; Fukuda, K.; Kanai, H.; Kuroki, S.; Hirakata, H. Reduced p21, p27 and vitamin d receptor in the nodular hyperplasia in patients with advanced secondary hyperparathyroidism. Kidney Int. 2002, 62, 1196–1207. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/12234290 (accessed on 20 December 2021). [CrossRef]
- Cozzolino, M.; Lu, Y.; Finch, J.; Slatopolsky, E.; Dusso, A.S. P21waf1 and tgf-alpha mediate parathyroid growth arrest by vitamin d and high calcium. Kidney Int. 2001, 60, 2109–2117. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/11737585 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Gogusev, J.; Duchambon, P.; Stoermann-Chopard, C.; Giovannini, M.; Sarfati, E.; Drueke, T.B. De novo expression of transforming growth factor-alpha in parathyroid gland tissue of patients with primary or secondary uraemic hyperparathyroidism. Nephrol. Dial. Transplant. 1996, 11, 2155–2162. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/8941573 (accessed on 20 December 2021). [CrossRef]
- Arcidiacono, M.V.; Sato, T.; Alvarez-Hernandez, D.; Yang, J.; Tokurl, M.; Gonzalez-Suarez, I.; Lu, Y.; Tominaga, Y.; Cannata-Andia, J.; Slatopolsky, E.; et al. Egfr activation increases parathyroid hyperplasia and calcitriol resistance in kidney disease. J. Am. Soc. Nephrol. 2008, 19, 310–320. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, M.; Lu, Y.; Sato, T.; Yang, J.; Suarez, I.G.; Brancaccio, D.; Slatopolsky, E.; Dusso, A.S. A critical role for enhanced tgf-alpha and egfr expression in the initiation of parathyroid hyperplasia in experimental kidney disease. Am. J. Physiol. Ren. Physiol. 2005, 289, F1096-102. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/15998841 (accessed on 20 December 2021). [CrossRef] [Green Version]
- Terada, Y.; Inoshita, S.; Nakashima, O.; Kuwahara, M.; Sasaki, S.; Marumo, F. Regulation of cyclin d1 expression and cell cycle progression by mitogen-activated protein kinase cascade. Kidney Int. 1999, 56, 1258–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcidiacono, M.V.; Yang, J.; Fernandez, E.; Dusso, A. Parathyroid-specific epidermal growth factor-receptor inactivation prevents uremia-induced parathyroid hyperplasia in mice. Nephrol. Dial. Transplant. 2015, 30, 434–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yang, Z.; Passaniti, A.; Lapidus, R.G.; Liu, X.; Cullen, K.J.; Dan, H.C. A positive feedback loop involving egfr/akt/mtorc1 and ikk/nf-kb regulates head and neck squamous cell carcinoma proliferation. Oncotarget 2016, 7, 31892–31906. Available online: https://pubmed.ncbi.nlm.nih.gov/26895469 (accessed on 20 December 2021). [CrossRef] [PubMed]
- Huse, J.T.; Holland, E.C. Targeting brain cancer: Advances in the molecular pathology of malignant glioma and medulloblastoma. Nat. Rev. Cancer 2010, 10, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. Tor signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Shimobayashi, M.; Hall, M.N. Making new contacts: The mtor network in metabolism and signalling crosstalk. Nat. Rev. Mol. Cell Biol. 2014, 15, 155–162. [Google Scholar] [CrossRef]
- Shahbazian, D.; Roux, P.P.; Mieulet, V.; Cohen, M.S.; Raught, B.; Taunton, J.; Hershey, J.W.B.; Blenis, J.; Pende, M.; Sonenberg, N. The mtor/pi3k and mapk pathways converge on eif4b to control its phosphorylation and activity. EMBO J. 2006, 25, 2781–2791. [Google Scholar] [CrossRef]
- Meyuhas, O. Physiological roles of ribosomal protein s6: One of its kind. Int. Rev. Cell Mol. Biol. 2008, 268, 1–37. [Google Scholar]
- Ruvinsky, I.; Katz, M.; Dreazen, A.; Gielchinsky, Y.; Saada, A.; Freedman, N.; Mishani, E.; Zimmerman, G.; Kasir, J.; Meyuhas, O. Mice deficient in ribosomal protein s6 phosphorylation suffer from muscle weakness that reflects a growth defect and energy deficit. PLoS ONE 2009, 4, e5618. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Chen, J.; Dong, Z.; Meyuhas, O.; Chen, J.K. Phosphorylation of ribosomal protein s6 mediates compensatory renal hypertrophy. Kidney Int. 2015, 87, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Volovelsky, O.; Cohen, G.; Kenig, A.; Wasserman, G.; Dreazen, A.; Meyuhas, O.; Silver, J.; Naveh-Many, T. Phosphorylation of ribosomal protein s6 mediates mammalian target of rapamycin complex 1-induced parathyroid cell proliferation in secondary hyperparathyroidism. J. Am. Soc. Nephrol. 2016, 27, 1091–1101. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/26283674 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Tan, X.; Wen, X.; Liu, Y. Paricalcitol inhibits renal inflammation by promoting vitamin d receptor-mediated sequestration of nf-kappab signaling. J. Am. Soc. Nephrol. 2008, 19, 1741–1752. Available online: https://pubmed.ncbi.nlm.nih.gov/18525004 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Delfino, F.; Walker, W.H. Hormonal regulation of the nf-kappab signaling pathway. Mol. Cell Endocrinol. 1999, 157, 1–9. [Google Scholar] [CrossRef]
- Nakanishi, C.; Toi, M. Nuclear factor-kappab inhibitors as sensitizers to anticancer drugs. Nat. Rev. Cancer 2005, 5, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Guttridge, D.C.; Albanese, C.; Reuther, J.Y.; Pestell, R.G.; Baldwin, A.S.J. Nf-kappab controls cell growth and differentiation through transcriptional regulation of cyclin d1. Mol. Cell Biol. 1999, 19, 5785–5799. [Google Scholar] [CrossRef] [Green Version]
- Heppner, C.; Bilimoria, K.Y.; Agarwal, S.K.; Kester, M.; Whitty, L.J.; Guru, S.C.; Chandrasekharappa, S.C.; Collins, F.S.; Spiegel, A.M.; Marx, S.J.; et al. The tumor suppressor protein menin interacts with nf-κb proteins and inhibits nf-κb-mediated transactivation. Oncogene 2001, 20, 4917–4925. [Google Scholar] [CrossRef] [Green Version]
- Corbetta, S.; Vicentini, L.; Ferrero, S.; Lania, A.; Mantovani, G.; Cordella, D.; Beck-Peccoz, P.; Spada, A. Activity and function of the nuclear factor kappab pathway in human parathyroid tumors. Endocr. Relat. Cancer 2005, 12, 929–937. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/16322332 (accessed on 20 December 2021). [CrossRef]
- Kan, S.; Zhang, W.; Mao, J.; Wang, M.; Ni, L.; Zhang, M.; Zhang, Q.; Chen, J. Nf-κb activation contributes to parathyroid cell proliferation in chronic kidney disease. J. Nephrol. 2018, 31, 941–951. [Google Scholar] [CrossRef]
- Mann, J.R.; Backlund, M.G.; DuBois, R.N. Mechanisms of disease: Inflammatory mediators and cancer prevention. Nat. Clin. Pract. Oncol. 2005, 2, 202–210. [Google Scholar] [CrossRef]
- Bell, C.D.; Vidal, S.; Kovacs, K.; Anderson, J.; Rotondo, F. Cox-2 expression in the oxyphilic cells of the normal, hyperplastic, and adenomatous parathyroid gland. Endocr. Pathol. 2004, 15, 29–38. [Google Scholar] [CrossRef]
- Zhang, Q.; Qiu, J.; Li, H.; Lu, Y.; Wang, X.; Yang, J.; Wang, S.; Zhang, L.; Gu, Y.; Hao, C.-M.; et al. Cyclooxygenase 2 promotes parathyroid hyperplasia in esrd. J. Am. Soc. Nephrol. 2011, 22, 664–672. Available online: https://pubmed.ncbi.nlm.nih.gov/21335517 (accessed on 20 December 2021). [CrossRef] [PubMed] [Green Version]
- Gardner, D.G.; Brown, E.M.; Windeck, R.; Aurbach, G.D. Prostaglandin e2 inhibits 3′,5′-adenosine monophosphate accumulation and parathyroid hormone release from dispersed bovine parathyroid cells. Endocrinology 1979, 104, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.G.; Brown, E.M.; Attie, M.F.; Aurbach, G.D. Prostaglandin-mediated stimulation of adenosine 3′,5′-monophosphate accumulation and parathyroid hormone release in dispersed human parathyroid cells. J. Clin. Endocrinol. Metab. 1980, 51, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, S.; Ye, G.; Wang, M.; Ni, L.; Kan, S.; Zhang, M.; Chen, J. Prostaglandin e2 receptor ep2 mediates the effect of cyclooxygenase 2 on secondary parathyroid hyperplasia in end-stage renal disease. Nephrol. Dial. Transplant. 2019, 34, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the clock protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Fujiki, R.; Murayama, A.; Kitagawa, H.; Yamaoka, K.; Yamamoto, Y.; Mihara, M.; Takeyama, K.; Kato, S. 1alpha,25(oh)2d3-induced transrepression by vitamin d receptor through e-box-type elements in the human parathyroid hormone gene promoter. Mol. Endocrinol. 2007, 21, 334–342. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17095575 (accessed on 20 December 2021). [CrossRef]
- Egstrand, S.; Nordholm, A.; Morevati, M.; Mace, M.L.; Hassan, A.; Naveh-Many, T.; Rukov, J.L.; Gravesen, E.; Olgaard, K.; Lewin, E. A molecular circadian clock operates in the parathyroid gland and is disturbed in chronic kidney disease associated bone and mineral disorder. Kidney Int. 2020, 98, 1461–1475. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0085253820308280 (accessed on 20 December 2021). [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, A.; Khalaily, N.; Kilav-Levin, R.; Nechama, M.; Volovelsky, O.; Silver, J.; Naveh-Many, T. Molecular Mechanisms of Parathyroid Disorders in Chronic Kidney Disease. Metabolites 2022, 12, 111. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12020111
Hassan A, Khalaily N, Kilav-Levin R, Nechama M, Volovelsky O, Silver J, Naveh-Many T. Molecular Mechanisms of Parathyroid Disorders in Chronic Kidney Disease. Metabolites. 2022; 12(2):111. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12020111
Chicago/Turabian StyleHassan, Alia, Nareman Khalaily, Rachel Kilav-Levin, Morris Nechama, Oded Volovelsky, Justin Silver, and Tally Naveh-Many. 2022. "Molecular Mechanisms of Parathyroid Disorders in Chronic Kidney Disease" Metabolites 12, no. 2: 111. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo12020111