
In Vitro Animal Model for Estimating the Time since Death with Attention to Early Postmortem Stage
 , , , , , and
, , , , , and 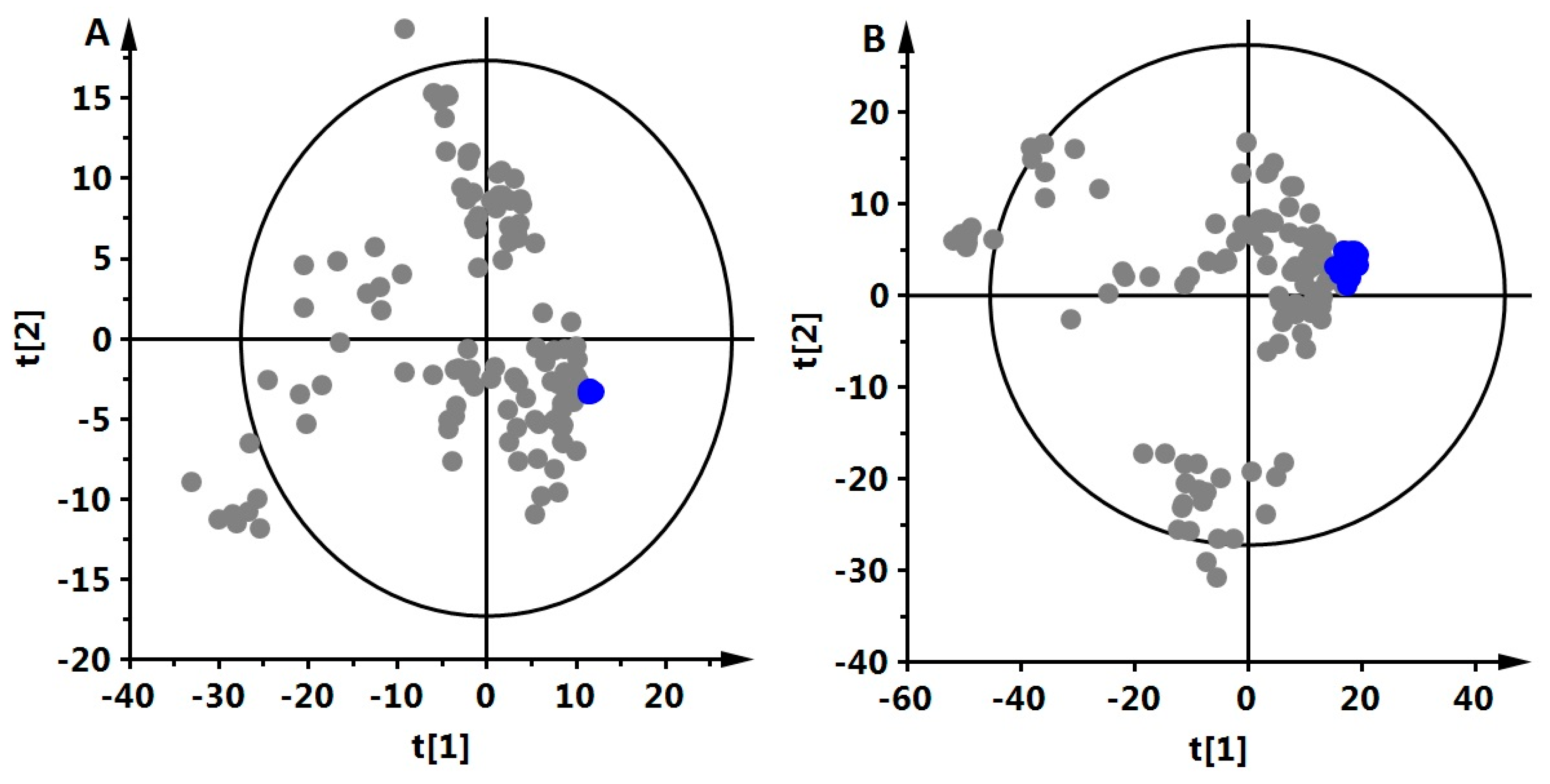{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Main Experiment Criteria
2.2. Ethics
2.3. Procedures
2.3.1. Chemicals and Reagents
2.3.2. Sample Processing
2.4. Assays
2.4.1. Plasma Metabolic Fingerprinting
2.4.2. LC-MS Data Processing
2.4.3. Statistics
2.4.4. Metabolite Identification
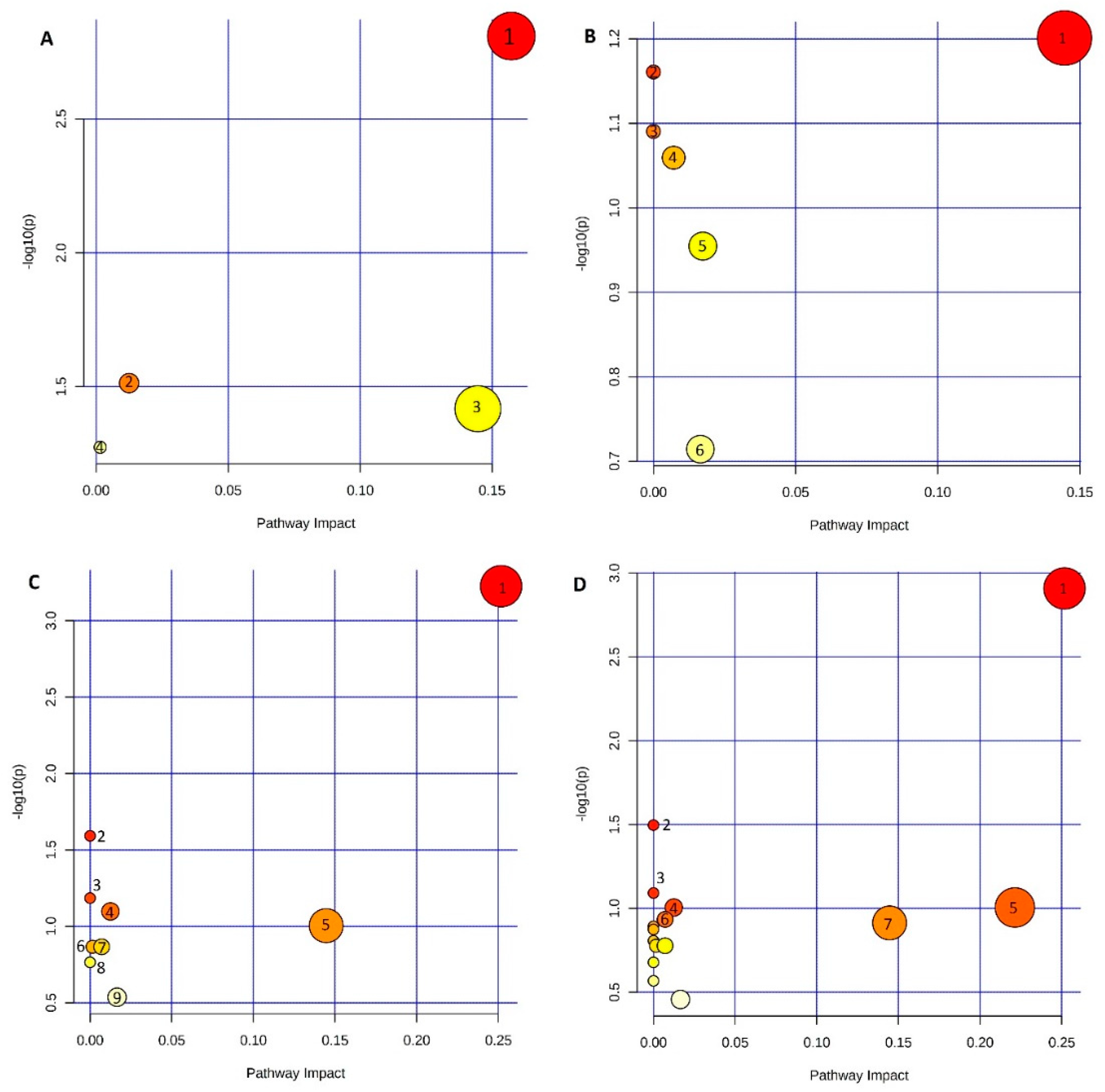
2.4.5. Metabolic Pathways Analysis
3. Results and Discussion
3.1. Hypoxanthine
3.2. Lactic Acid
3.3. Histidine
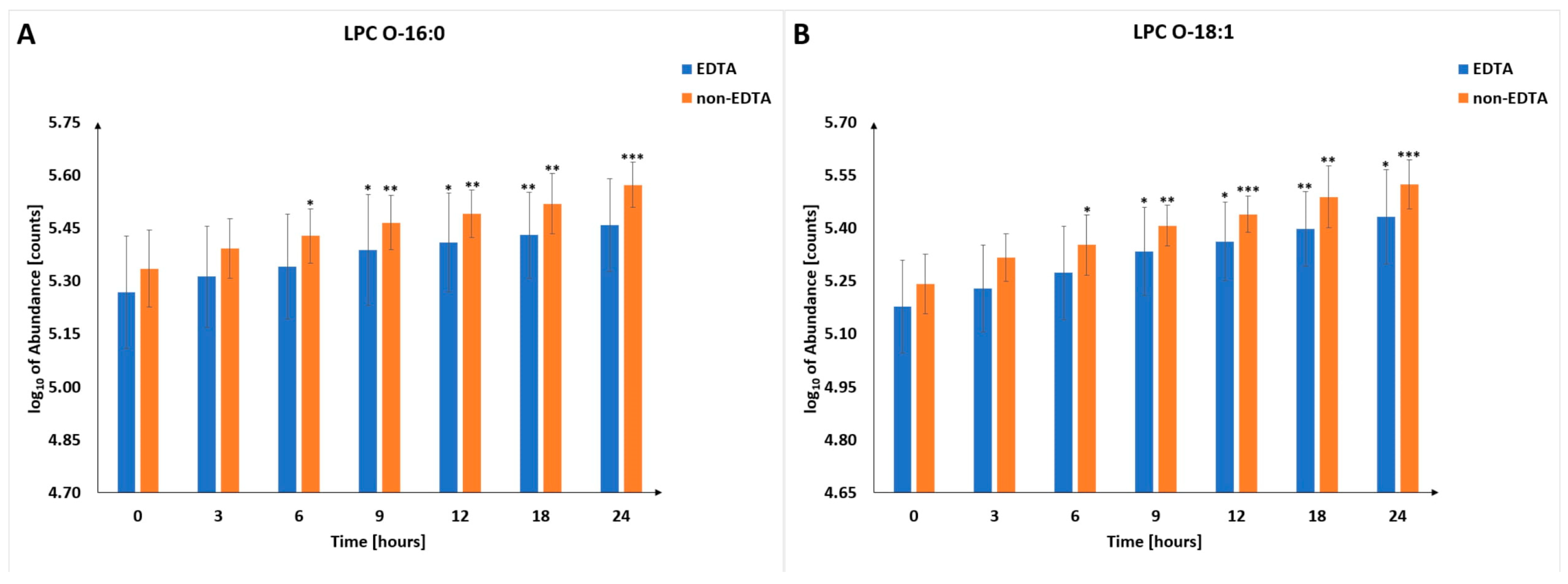
3.4. Lysophosphatidic Acids (LPAs)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mathur, A.; Agrawal, Y.K. An overview of methods used for estimation of time since death. Aust. J. Forensic Sci. 2011, 43, 275–285. [Google Scholar] [CrossRef]
- Ingham, A.I.; Byard, R.W. The potential significance of elevated vitreous sodium levels at autopsy. J. Forensic Leg. Med. 2009, 16, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Passos, M.L.C.; Santos, A.M.; Pereira, A.I.; Rodrigo Santos, J.; Santos, A.J.C.; Saraiva, M.L.M.F.S.; Lima, J.L.F.C. Estimation of postmortem interval by hypoxanthine and potassium evaluation in vitreous humor with a sequential injection system. Talanta 2009, 79, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Yahia, D.; El-Amir, Y.O.; Sadek, A.A.I. Early postmortem biochemical and histopathological changes in the kidney, liver, and muscles of dogs. Comp. Clin. Pathol. 2018, 27, 1447–1455. [Google Scholar] [CrossRef]
- Sampaio-Silva, F.; Magalhães, T.; Carvalho, F.; Dinis-Oliveira, R.; Silvestre, R. Profiling of RNA degradation for estimation of post mortem [corrected] interval. PLoS ONE 2013, 8, e56507. [Google Scholar] [CrossRef]
- Larkin, B.; Iaschi, S.; Dadour, I.; Tay, G.K. Using accumulated degree-days to estimate postmortem interval from the DNA yield of porcine skeletal muscle. Forensic Sci. Med. Pathol. 2010, 6, 83–92. [Google Scholar] [CrossRef]
- Matuszewski, S.; Bajerlein, D.; Konwerski, S.; Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 1: Pattern and rate of decomposition. Forensic Sci. Int. 2010, 1964, 85–93. [Google Scholar] [CrossRef]
- Matuszewski, S.; Bajerlein, D.; Konwerski, S.; Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 2: Composition and residency patterns of carrion fauna. Forensic Sci. Int. 2010, 195, 42–51. [Google Scholar] [CrossRef]
- Malewski, T.; Draber-Mońko, A.; Pomorski, J.; Łoś, M.; Bogdanowicz, W. Identification of forensically important blowfly species (Diptera: Calliphoridae) by high-resolution melting PCR analysis. Peer J. 2020, 8, e9680. [Google Scholar] [CrossRef]
- Chandrakanth, H.V.; Kanchan, T.; Balaraj, B.M.; Virupaksha, H.S.; Chandrashekar, T.N. Postmortem vitreous chemistry--an evaluation of sodium, potassium and chloride levels in estimation of time since death (during the first 36 h after death). J. Forensic Leg. Med. 2013, 20, 211–216. [Google Scholar] [CrossRef]
- Bardale, R.V.; Tumram, N.K.; Dixit, P.G.; Deshmukh, A.Y. Evaluation of histologic changes of the skin in postmortem period. Am. J. Forensic Med. Pathol. 2012, 33, 357–361. [Google Scholar] [CrossRef]
- Abo El-Noor, M.M.; Elhosary, N.M.; Khedr, N.F.; El-Desouky, K.I. Estimation of Early Postmortem Interval Through Biochemical and Pathological Changes in Rat Heart and Kidney. Am. J. Forensic Med. Pathol 2016, 37, 40–46. [Google Scholar] [CrossRef]
- Sener, M.T.; Suleyman, H.; Hacimuftuoglu, A.; Polat, B.; Cetin, N.; Suleyman, B.; Akcay, F. Estimating the postmortem interval by the difference between oxidant/antioxidant parameters in liver tissue. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2012, 21, 727–733. [Google Scholar]
- Tumram, N.K.; Bardale, R.V.; Dongre, A.P. Postmortem analysis of synovial fluid and vitreous humour for determination of death interval: A comparative study. Forensic Sci. Int. 2011, 204, 186–190. [Google Scholar] [CrossRef]
- Swain, R.; Kumar, A.; Sahoo, J.; Lakshmy, R.; Gupta, S.K.; Bhardwaj, D.N.; Pandey, R.M. Estimation of post-mortem interval: A comparison between cerebrospinal fluid and vitreous humour chemistry. J. Forensic Leg. Med. 2015, 36, 144–148. [Google Scholar] [CrossRef]
- Metcalf, J.L. Estimating the postmortem interval using microbes: Knowledge gaps and a path to technology adoption. Forensic Sci. Int. Genet. 2019, 38, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.L.; Xu, Z.Z.; Weiss, S.; Lax, S.; Van Treuren, W.; Hyde, E.R.; Song, S.J.; Amir, A.; Larsen, P.; Sangwan, N.; et al. Microbial community assembly and metabolic function during mammalian corpse decomposition. Science 2016, 351, 158. [Google Scholar] [CrossRef] [Green Version]
- Szeremeta, M.; Pietrowska, K.; Niemcunowicz-Janica, A.; Kretowski, A.; Ciborowski, M. Applications of Metabolomics in Forensic Toxicology and Forensic Medicine. Int. J. Mol. Sci. 2021, 22, 3010. [Google Scholar] [CrossRef]
- Harada, H.; Maeiwa, M.; Yoshikawa, K.; Ohsaka, A. Identification and quantitation by 1H-NMR of metabolites in animal organs and tissues. An application of NMR spectroscopy in forensic science. Forensic Sci. Int. 1984, 24, 1–7. [Google Scholar] [CrossRef]
- Mao, S.; Fu, G.; Seese, R.R.; Wang, Z.-Y. Estimation of PMI depends on the changes in ATP and its degradation products. Leg. Med. (Tokyo). 2013, 15, 235–238. [Google Scholar] [CrossRef]
- Donaldson, A.E.; Lamont, I.L. Biochemistry changes that occur after death: Potential markers for determining post-mortem interval. PLOS ONE 2013, 8, e82011. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, A.E.; Lamont, I.L. Metabolomics of post-mortem blood: Identifying potential markers of post-mortem interval. Metabolomics 2015, 11, 237–245. [Google Scholar] [CrossRef]
- Dai, X.; Fan, F.; Ye, Y.; Lu, X.; Chen, F.; Wu, Z.; Liao, L. An experimental study on investigating the postmortem interval in dichlorvos poisoned rats by GC/MS-based metabolomics. Leg. Med. 2019, 36, 28–36. [Google Scholar] [CrossRef]
- Fang, S.Y.; Dai, X.H.; Xiao, L.; Zou, J.; Yang, L.; Ye, Y.; Liao, L.C. Estimation of Early Postmortem Interval of Asphyxial Death Rats at Different Ambient Temperatures by GC-MS-Based Metabolomics. Fa Yi Xue Za Zhi. 2020, 36, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Takeichi, S.; Wakasugi, C.; Shikata, I. Fluidity of cadaveric blood after sudden death: Part I. Postmortem fibrinolysis and plasma catecholamine level. Am. J. Forensic Med. Pathol. 1984, 5, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Daniluk, U.; Daniluk, J.; Kucharski, R.; Kowalczyk, T.; Pietrowska, K.; Samczuk, P.; Filimoniuk, A.; Kretowski, A.; Lebensztejn, D.; Ciborowski, M. Untargeted Metabolomics and Inflammatory Markers Profiling in Children with Crohn’s Disease and Ulcerative Colitis-A Preliminary Study. Inflamm. Bowel Dis. 2019, 25, 1120–1128. Inflamm. Bowel Dis. 2019, 25, 1120–1128. [Google Scholar] [CrossRef] [Green Version]
- Armitage, E.G.; Godzien, J.; Alonso-Herranz, V.; López-Gonzálvez, Á.; Barbas, C. Missing value imputation strategies for metabolomics data. Electrophoresis 2015, 36, 3050–3060. [Google Scholar] [CrossRef]
- Pietrowska, K.; Dmuchowska, D.A.; Samczuk, P.; Kowalczyk, T.; Krasnicki, P.; Wojnar, M.; Skowronska, A.; Mariak, Z.; Kretowski, A.; Ciborowski, M. LC-MS-Based Metabolic Fingerprinting of Aqueous Humor. J. Anal. Methods Chem. 2017, 2017, 6745932. [Google Scholar] [CrossRef] [Green Version]
- Krupska, O.; Kowalczyk, T.; Beręsewicz-Haller, M.; Samczuk, P.; Pietrowska, K.; Zabłocki, K.; Kretowski, A.; Ciborowski, M.; Zabłocka, B. Hippocampal Sector–Specific Metabolic Profiles Reflect Endogenous Strategy for Ischemia-Reperfusion Insult Resistance. Mol. Neurobiol. 2021, 58, 1621–1633. [Google Scholar] [CrossRef]
- Godzien, J.; Ciborowski, M.; Martínez-Alcázar, M.P.; Samczuk, P.; Kretowski, A.; Barbas, C. Rapid and Reliable Identification of Phospholipids for Untargeted Metabolomics with LC-ESI-QTOF-MS/MS. J. Proteome Res. 2015, 14, 3204–3216. [Google Scholar] [CrossRef]
- Ciborowski, M.; Teul, J.; Martin-Ventura, J.L.; Egido, J.; Barbas, C. Metabolomics with LC-QTOF-MS permits the prediction of disease stage in aortic abdominal aneurysm based on plasma metabolic fingerprint. PLoS ONE 2012, 7, e31982. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Jawor, P.; Ząbek, A.; Wojtowicz, W.; Król, D.; Stefaniak, T.; Młynarz, P.A.-O. Metabolomic studies as a tool for determining the post-mortem interval (PMI) in stillborn calves. BMC Vet. Res. 2019, 15, 189. [Google Scholar] [CrossRef] [Green Version]
- Zelentsova, E.A.; Yanshole, L.V.; Snytnikova, O.A.; Yanshole, V.V.; Tsentalovich, Y.P.; Sagdeev, R.Z. Post-mortem changes in the metabolomic compositions of rabbit blood, aqueous and vitreous humors. Metabolomics 2016, 12, 172. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Juan, G.; Li, X.; Gorczyca, W.; Murakami, T.; Traganos, F. Cytometry in cell necrobiology: Analysis of apoptosis and accidental cell death (necrosis). Cytometry 1997, 27, 1–20. [Google Scholar] [CrossRef]
- Aoki, J.; Inoue, A.; Okudaira, S. Two pathways for lysophosphatidic acid production. Biophys. Acta. 2008, 1781, 513–518. [Google Scholar] [CrossRef]
- Pagès, C.; Simon, M.F.; Valet, P.; Saulnier-Blache, J.S. Lysophosphatidic acid synthesis and release. Prostaglandins Other Lipid Mediat. 2001, 64, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.E.; Herr, D.R.; Chun, J. Lysophosphatidic acid (LPA) receptors: Signaling properties and disease relevance. Prostaglandins Other Lipid Mediat. 2010, 91, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Karliner, J.S.; Honbo, N.; Summers, K.; Gray, M.O.; Goetzl, E.J. The lysophospholipids sphingosine-1-phosphate and lysophosphatidic acid enhance survival during hypoxia in neonatal rat cardiac myocytes. J. Mol. Cell Cardiol. 2001, 33, 1713–1717. [Google Scholar] [CrossRef]
- Chen, J.; Baydoun, A.R.; Xu, R.; Deng, L.; Liu, X.; Zhu, W.; Shi, L.; Cong, X.; Hu, S.; Chen, X. Lysophosphatidic acid protects mesenchymal stem cells against hypoxia and serum deprivation-induced apoptosis. Stem Cells 2008, 26, 135–145. [Google Scholar] [CrossRef]
- Okusa, M.D.; Ye, H.; Huang, L.; Sigismund, L.; Macdonald, T.; Lynch, K.R. Selective blockade of lysophosphatidic acid LPA3 receptors reduces murine renal ischemia-reperfusion injury. Am. J. Physiol. Renal. Physiol. 2003, 285, F565–F574. [Google Scholar] [CrossRef] [PubMed]
- Cotran, R.; Kumar, V.; Robbins, S. Cellular Injury and Cellular Death. In Robbins Pathologic Basis of disease, 5th ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1995; pp. 4–11. [Google Scholar]
- Sawyer, W.R.; Steup, D.r.; Martin, B.S.; Forney, R.B. Cardiac blood pH as a possible indicator of postmortem interval. J. Forensic Sci. 1988, 33, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K. Histidine and lysine as targets of oxidative modification. Amino Acids 2003, 25, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Remko, M.; Fitz, D.; Rode, B.M. Effect of metal ions (Li+, Na+, K+, Mg2+, Ca2+, Ni2+, Cu2+ and Zn2+) and water coordination on the structure and properties of L-histidine and zwitterionic L-histidine. Amino Acids 2010, 39, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fitz, D.; Fraser, D.G.; Rode, B.M. Catalytic effects of histidine enantiomers and glycine on the formation of dileucine and dimethionine in the salt-induced peptide formation reaction. Amino Acids 2010, 38, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Norton, D.; Crow, B.; Bishop, M.; Kovalcik, K.; George, J.; Bralley, J.A. High performance liquid chromatography-tandem mass spectrometry (HPLC/MS/MS) assay for chiral separation of lactic acid enantiomers in urine using a teicoplanin based stationary phase. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2007, 850, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Cevasco, G.; Piątek, A.M.; Scapolla, C.; Thea, S. A simple, sensitive and efficient assay for the determination of D- and L-lactic acid enantiomers in human plasma by high-performance liquid chromatography. J. Chromatogr. A 2011, 1218, 787–792. [Google Scholar] [CrossRef]
- Madea, B. Methods for determining time of death. Forensic Sci. Med. Pathol. 2016, 12, 451–485. [Google Scholar] [CrossRef]
- Rognum, T.O.; Holmen, S.; Musse, M.A.; Dahlberg, P.S.; Stray-Pedersen, A.; Saugstad, O.D.; Opdal, S.H. Estimation of time since death by vitreous humor hypoxanthine, potassium, and ambient temperature. Forensic Sci. Int. 2016, 262, 160–165. [Google Scholar] [CrossRef]
- Gurler, M.; Ozturk, G.; Kir, M.Z.; Ginis, Z.; Erden, G.; Akyol, S.; Kaya, M.; Karapirli, M.; Akyol, O. Simultaneous analysis of biochemical markers in vitreous humour and serum: A preliminary study on the effect of storage time at −20 °C. Aust. J. Forensic Sci. 2016, 48, 150–158. [Google Scholar] [CrossRef]
- Zelentsova, E.A.; Yanshole, L.V.; Melnikov, A.D.; Kudryavtsev, I.S.; Novoselov, V.P.; Tsentalovich, Y.P. Post-mortem changes in metabolomic profiles of human serum, aqueous humor and vitreous humor. Metabolomics 2020, 16, 80. [Google Scholar] [CrossRef]
- Boulieu, R.; Bory, C.; Baltassat, P.; Gonnet, C. Hypoxanthine and xanthine levels determined by high-performance liquid chromatography in plasma, erythrocyte, and urine samples from healthy subjects: The problem of hypoxanthine level evolution as a function of time. Anal. Biochem. 1983, 129, 398–404. [Google Scholar] [CrossRef]
- Poulsen, J.P.; Oyasaeter, S.; Sanderud, J.; Rognum, T.O.; Saugstad, O.D. Hypoxanthine, xanthine, and uric acid concentrations in the cerebrospinal fluid, plasma, and urine of hypoxemic pigs. Pediatr. Res. 1990, 28, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Rognum, T.O.; Hauge, S.; Oyasaeter, S.; Saugstad, O.D. A new biochemical method for estimation of postmortem time. Forensic Sci. Int. 1991, 51, 139–146. [Google Scholar] [CrossRef]
- Zhu, B.-L.; Ishikawa, T.; Michiue, T.; Tanaka, S.; Zhao, D.; Li, D.-R.; Quan, L.; Oritani, S.; Maeda, H. Differences in postmortem urea nitrogen, creatinine and uric acid levels between blood and pericardial fluid in acute death. Leg. Med. 2007, 9, 115–122. [Google Scholar] [CrossRef]
- Keltanen, T.; Nenonen, T.; Ketola, R.A.; Ojanperä, I.; Sajantila, A.; Lindroos, K. Post-mortem analysis of lactate concentration in diabetics and metformin poisonings. Int. J. Legal. Med. 2015, 129, 1225–1231. [Google Scholar] [CrossRef]
- Boulagnon, C.; Garnotel, R.; Fornes, P.; Gillery, P. Post-mortem biochemistry of vitreous humor and glucose metabolism: An update. Clin. Chem. Lab. Med. 2011, 49, 1265–1270. [Google Scholar] [CrossRef]
- Jetter, W.W.; McLean, R. Biochemical Changes in Body Fluids After Death. Am. J. Clin. Pathol. 1943, 13, 178–185. [Google Scholar] [CrossRef]
- Schleyer, F. Determination of the time of death in the early postmortem interval. In Methods of Forensic Science; Lundquist, F., Ed.; Interscience: New York, NY, USA, 1963; Volume 2, pp. 253–293. [Google Scholar]
- Sato, T.; Zaitsu, K.; Tsuboi, K.; Nomura, M.; Kusano, M.; Shima, N.; Abe, S.; Ishii, A.; Tsuchihashi, H.; Suzuki, K. A preliminary study on postmortem interval estimation of suffocated rats by GC-MS/MS-based plasma metabolic profiling. Anal. Bioanal. Chem. 2015, 407, 3659–3665. [Google Scholar] [CrossRef]
- Katsumata, Y.; Sato, K.; Yada, S.; Uematsu, T.; Oya, M.; Yoshino, M. Anaerobic metabolism in dogs after organismal death. Nihon hōigaku zasshi Jpn. J. Leg. Med. 1983, 37, 75–78. [Google Scholar] [PubMed]
- Uemura, K.; Shintani-Ishida, K.; Saka, K.; Nakajima, M.; Ikegaya, H.; Kikuchi, Y.; Yoshida, K. Biochemical blood markers and sampling sites in forensic autopsy. Biochemical blood markers and sampling sites in forensic autopsy. J. Forensic Leg. Med. 2008, 15, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Fekete, J.F.; Brunsdon, D.F.V. The Use of Routine Laboratory Tests in Postmortem Examinations. Can. Soc. Forensic Sci. J. 1974, 7, 238–254. [Google Scholar] [CrossRef]
- Frezza, C.; Zheng, L.; Tennant, D.A.; Papkovsky, D.B.; Hedley, B.A.; Kalna, G.; Watson, D.G.; Gottlieb, E. Metabolic Profiling of Hypoxic Cells Revealed a Catabolic Signature Required for Cell Survival. PLoS ONE 2011, 6, e24411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solberg, R.; Enot, D.; Deigner, H.-P.; Koal, T.; Scholl-Bürgi, S.; Saugstad, O.D.; Keller, M. Metabolomic Analyses of Plasma Reveals New Insights into Asphyxia and Resuscitation in Pigs. PLoS ONE 2010, 5, e9606. [Google Scholar] [CrossRef]
- Lametsch, R.; Roepstorff, P.; Bendixen, E. Identification of Protein Degradation during Post-mortem Storage of Pig Meat. J. Agric. Food Chem. 2002, 50, 5508–5512. [Google Scholar] [CrossRef]
- Baker, D.L.; Umstot, E.S.; Desiderio, D.M.; Tigyi, G.J. Quantitative analysis of lysophosphatidic acid in human blood fractions. Ann. N. Y. Acad. Sci. 2000, 905, 267–269. [Google Scholar] [CrossRef]
- Gerrard, J.M.; Robinson, P. Identification of the molecular species of lysophosphatidic acid produced when platelets are stimulated by thrombin. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1989, 1001, 282–285. [Google Scholar] [CrossRef]
- Hayashi, K.; Takahashi, M.; Nishida, W.; Yoshida, K.; Ohkawa, Y.; Kitabatake, A.; Aoki, J.; Arai, H.; Sobue, K. Phenotypic modulation of vascular smooth muscle cells induced by unsaturated lysophosphatidic acids. Circ. Res. 2001, 89, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Eichholtz, T.; Jalink, K.; Fahrenfort, I.; Moolenaar, W.H. The bioactive phospholipid lysophosphatidic acid is released from activated platelets. Biochem. J. 1993, 291, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Nishida, W.; Hayashi, K.i.; Ohkawa, Y.; Ogawa, A.; Aoki, J.; Arai, H.; Sobue, K. Vascular Remodeling Induced by Naturally Occurring Unsaturated Lysophosphatidic Acid In Vivo. Circulation 2003, 108, 1746–1752. [Google Scholar] [CrossRef] [Green Version]
- Aikawa, S.; Hashimoto, T.; Kano, K.; Aoki, J. Lysophosphatidic acid as a lipid mediator with multiple biological actions. J. Biochem. 2015, 157, 81–89. [Google Scholar] [CrossRef]
- Stoddard, N.C.; Chun, J. Promising pharmacological directions in the world of lysophosphatidic Acid signaling. Biomol. Ther. 2015, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yung, Y.C.; Stoddard, N.C.; Chun, J. LPA receptor signaling: Pharmacology, physiology, and pathophysiology. J. Lipid Res. 2014, 55, 1192–1214. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, L.H.M.; Spohr, T.C.L.d.S.; Amaral, R.F.d.; Fonseca, A.C.C.d.; Garcia, C.; Mendes, F.d.A.; Freitas, C.; dosSantos, M.F.; Lima, F.R.S. Role of lysophosphatidic acid and its receptors in health and disease: Novel therapeutic strategies. Signal Transduct. Target. Ther. 2021, 6, 45. [Google Scholar] [CrossRef]
- Brindley, D.N.; Pilquil, C. Lipid phosphate phosphatases and signaling. J. Lipid Res. 2009, 50, S225–S230. [Google Scholar] [CrossRef] [Green Version]
- Bandoh, K.; Aoki, J.; Hosono, H.; Kobayashi, S.; Kobayashi, T.; Murakami-Murofushi, K.; Tsujimoto, M.; Arai, H.; Inoue, K. Molecular cloning and characterization of a novel human G-protein-coupled receptor, EDG7, for lysophosphatidic acid. J. Biol. Chem. 1999, 274, 27776–27785. [Google Scholar] [CrossRef] [Green Version]
- Bandoh, K.; Aoki, J.; Taira, A.; Tsujimoto, M.; Arai, H.; Inoue, K. Lysophosphatidic acid (LPA) receptors of the EDG family are differentially activated by LPA species. Structure-activity relationship of cloned LPA receptors. FEBS Lett. 2000, 478, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Yanagida, K.; Masago, K.; Nakanishi, H.; Kihara, Y.; Hamano, F.; Tajima, Y.; Taguchi, R.; Shimizu, T.; Ishii, S. Identification and characterization of a novel lysophosphatidic acid receptor, p2y5/LPA6. J. Biol. Chem. 2009, 284, 17731–17741. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Yung, Y.C.; Chen, A.; Chun, J. Lysophosphatidic acid signalling in development. Development 2015, 142, 1390–1395. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szeremeta, M.; Samczuk, P.; Pietrowska, K.; Kowalczyk, T.; Przeslaw, K.; Sieminska, J.; Kretowski, A.; Niemcunowicz-Janica, A.; Ciborowski, M. In Vitro Animal Model for Estimating the Time since Death with Attention to Early Postmortem Stage. Metabolites 2023, 13, 26. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo13010026
Szeremeta M, Samczuk P, Pietrowska K, Kowalczyk T, Przeslaw K, Sieminska J, Kretowski A, Niemcunowicz-Janica A, Ciborowski M. In Vitro Animal Model for Estimating the Time since Death with Attention to Early Postmortem Stage. Metabolites. 2023; 13(1):26. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo13010026
Chicago/Turabian StyleSzeremeta, Michal, Paulina Samczuk, Karolina Pietrowska, Tomasz Kowalczyk, Katarzyna Przeslaw, Julia Sieminska, Adam Kretowski, Anna Niemcunowicz-Janica, and Michal Ciborowski. 2023. "In Vitro Animal Model for Estimating the Time since Death with Attention to Early Postmortem Stage" Metabolites 13, no. 1: 26. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo13010026