The Influence of Drying Temperatures on the Metabolic Profiles and Antioxidant Activity of Manilkara zapota Leaves

 ,
,  and
and
Abstract
:1. Introduction
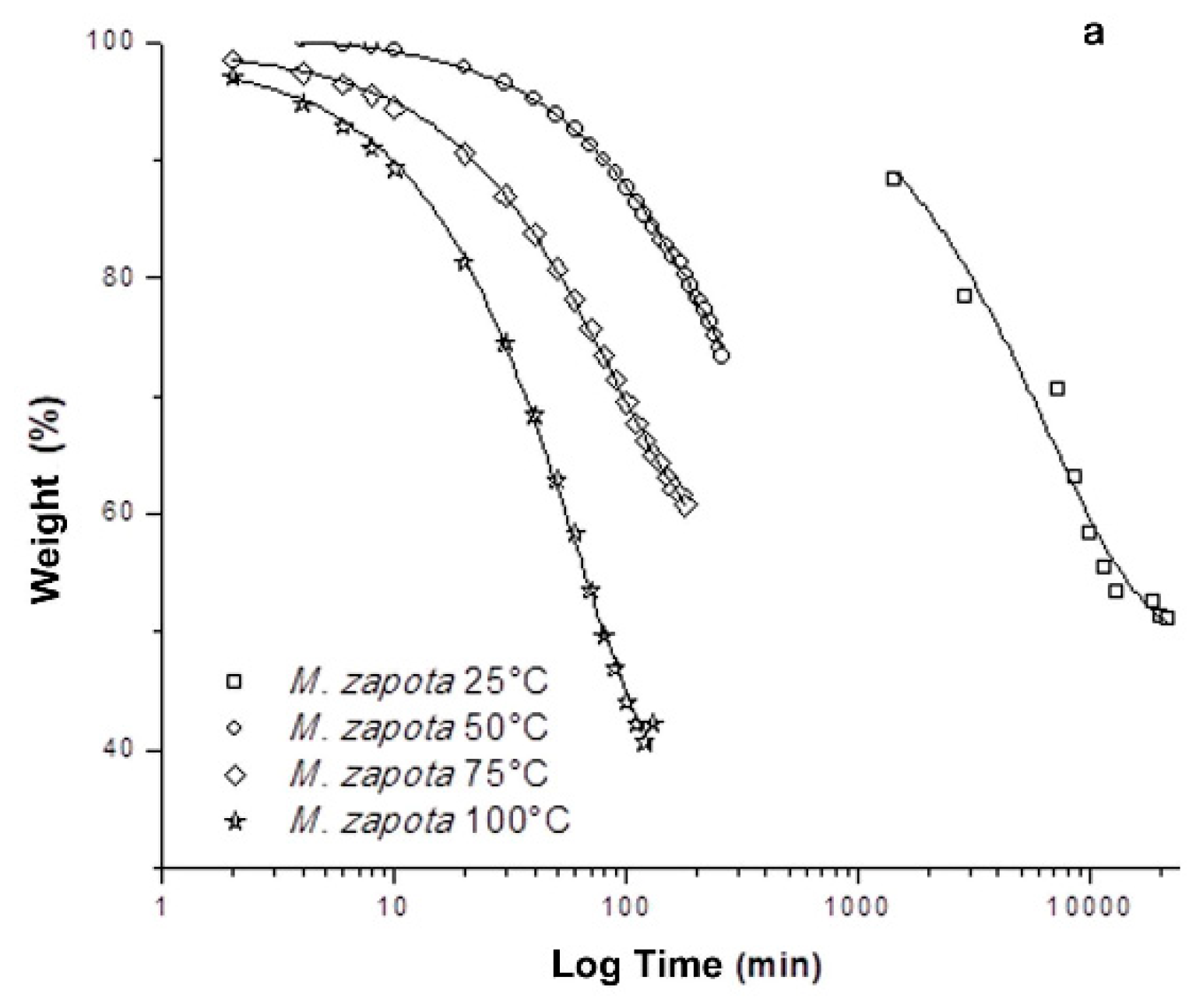
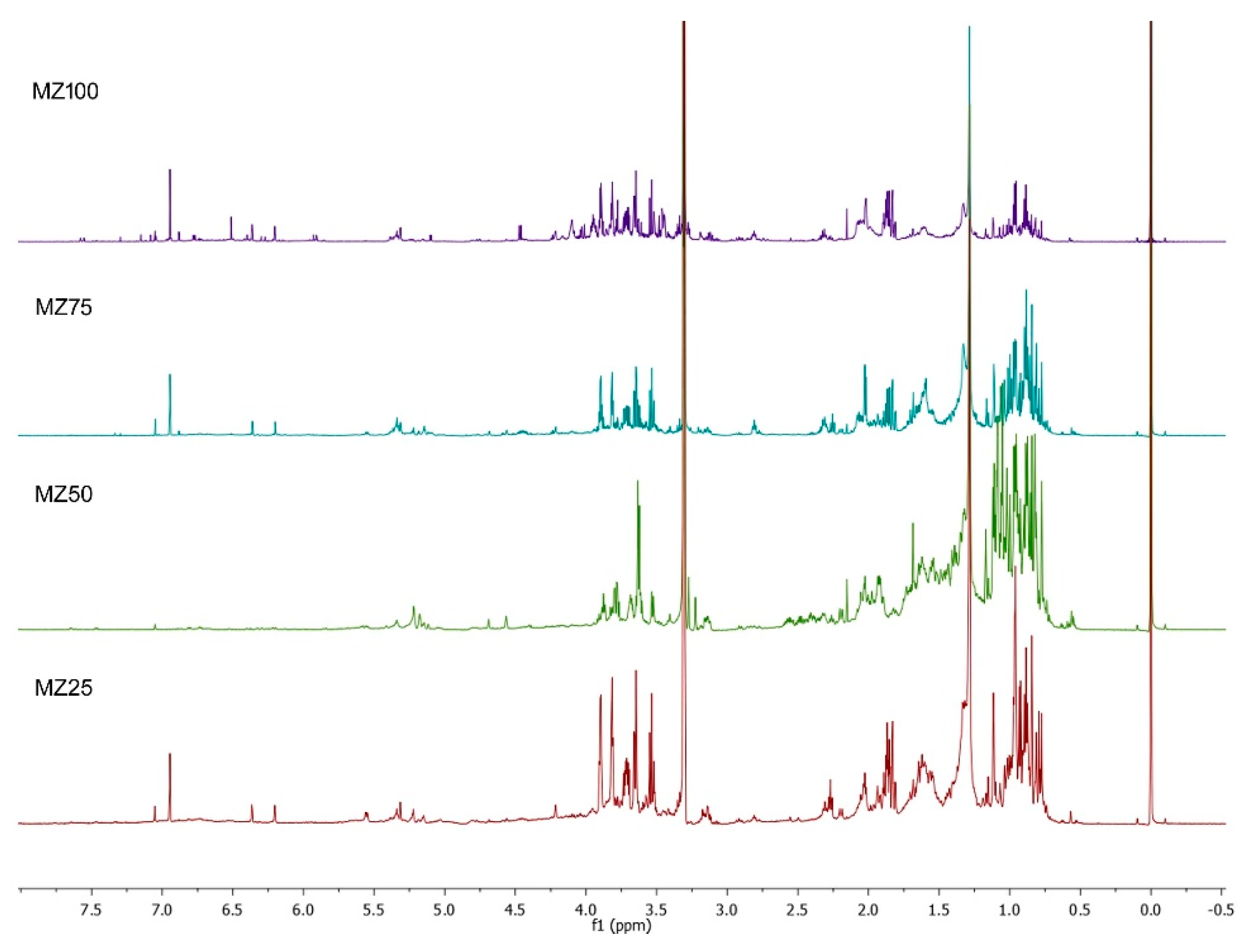
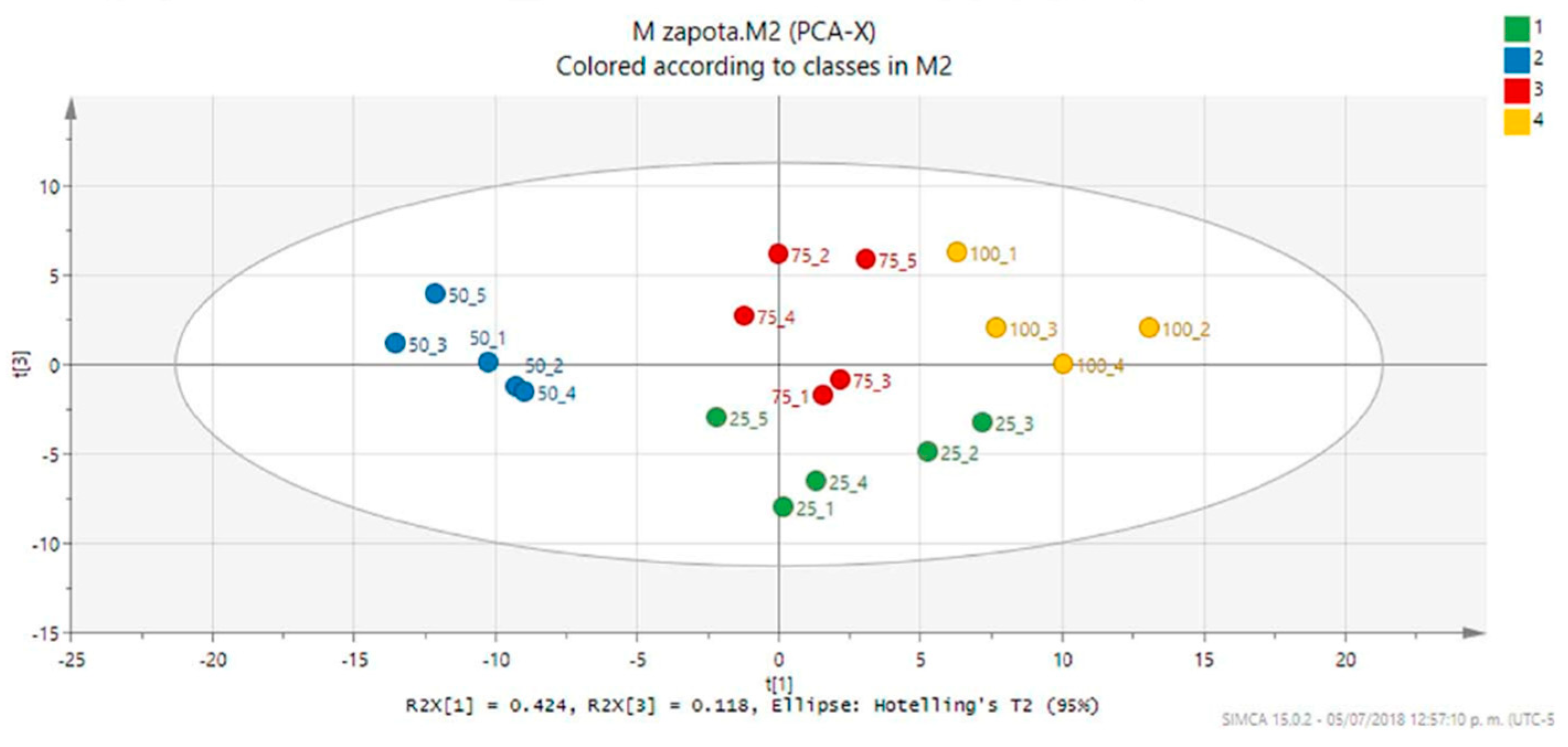
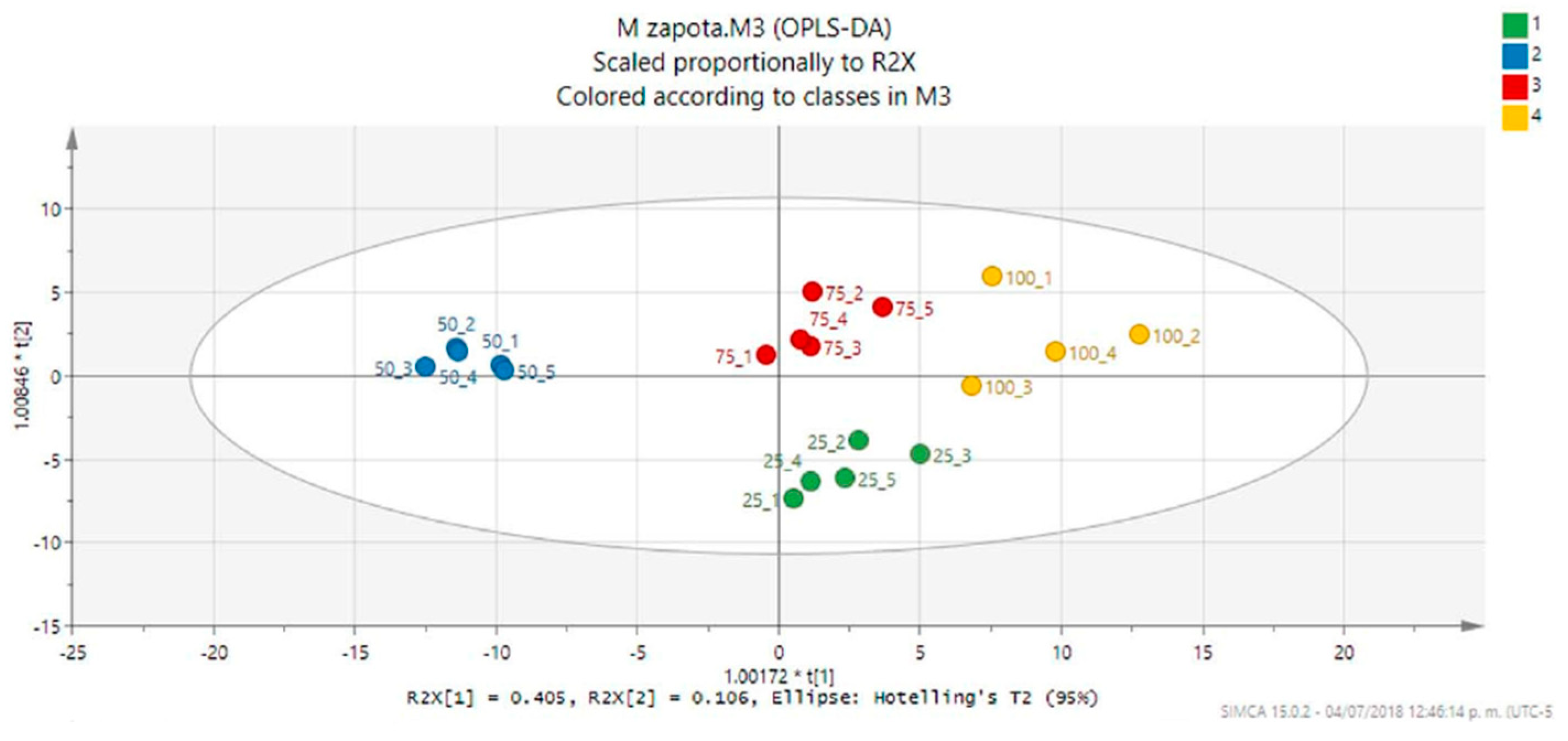
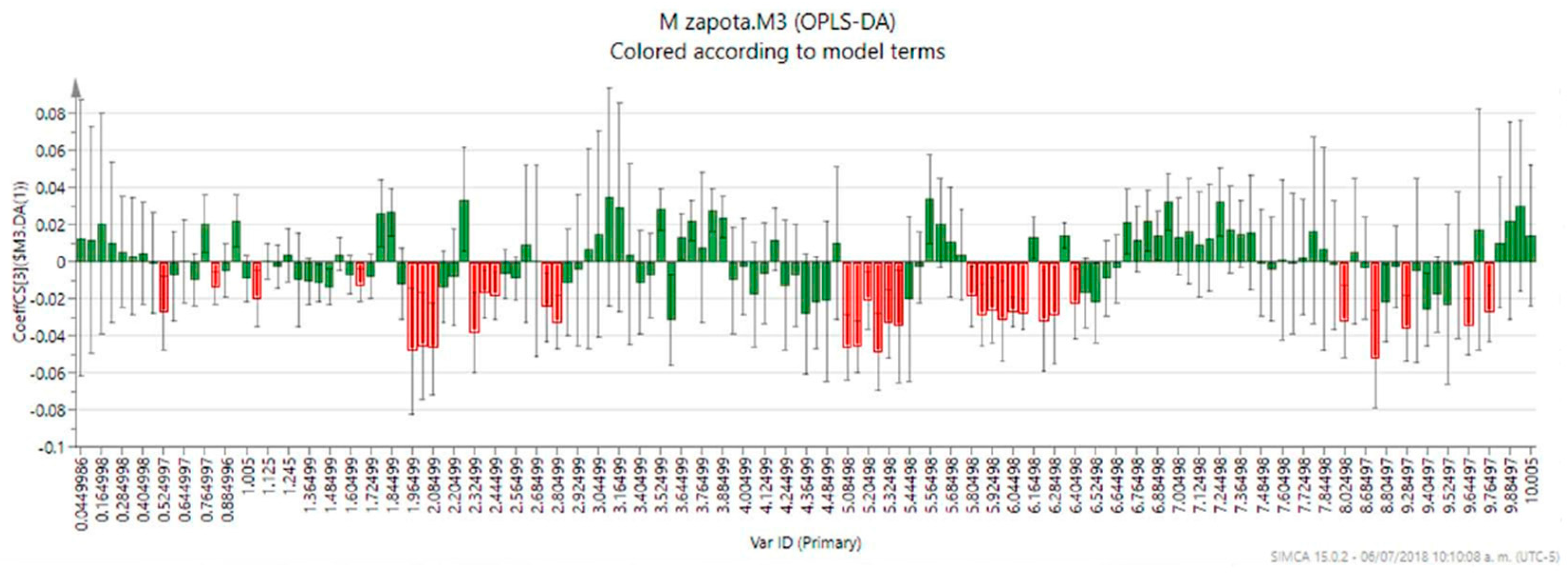
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material and Drying
3.3. Preparation of Plant Extracts
3.4. Total Phenol Content
3.5. DPPH Assay
3.6. 1H-NMR Metabolic Profiles and Multivariate Analysis
3.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bucar, F.; Wube, A.; Schmid, M. Natural product isolation - how to get from biological material to pure compounds. Nat. Prod. Rep. 2013, 30, 525–545. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Nybom, H.; Lindholm, C.; Rumpunen, K. Major polyphenols in aerial organs of greater plantain (Plantago major L.), and effects of drying temperature on polyphenol contents in the leaves. Sci. Hortic. 2011, 128, 523–529. [Google Scholar] [CrossRef]
- Katsube, T.; Tsurunaga, Y.; Sugiyama, M.; Furuno, T.; Yamasaki, Y. Effect of air-drying temperature on antioxidant capacity and stability of polyphenolic compounds in mulberry (Morus alba L.) leaves. Food Chem. 2009, 113, 964–969. [Google Scholar] [CrossRef]
- Darfour, B.; Asare, K.; Ofosu, O.; Daniel, G.A.; Elom, S.A.; Agbenyegah, S. The Effect of Different Drying Methods on the Phytochemicals and Radical Scavenging Activity of Ceylon Cinnamon (Cinnamomum zeylanicum) Plant Parts. Eur. J. Med. Plants 2014, 4, 1324–1335. [Google Scholar]
- Mediani, A.; Abas, F.; Tan, C.; Khatib, A. Effects of Different Drying Methods and Storage Time on Free Radical Scavenging Activity and Total Phenolic Content of Cosmos Caudatus. Antioxidants 2014, 3, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.O.; Myers, J.E. Transferencia de Cantidad de Movimiento, Calor y Materia; Editorial Reverté: Bogota, Colombia, 1979; ISBN 9788429170528. [Google Scholar]
- Tropicos.org Missouri Botanical Garden. Available online: http://www.tropicos.org/Name/28700370 (accessed on 17 December 2018).
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestree Database: A Tree Reference and Selection Guide Version 4.0. Available online: http://www.worldagroforestry.org/treedb/AFTPDFS/Manilkara_zapota.PDF (accessed on 11 February 2019).
- Biblioteca Digital de la Medicina Tradicional Mexicana Manilkara zapota. Available online: http://www.medicinatradicionalmexicana.unam.mx/monografia.php?l=3&t=Chicozapote&id=7519 (accessed on 20 January 2019).
- Chanda, S.V.; Nagani, K.V. Antioxidant Capacity of Manilkara zapota L. Leaves Extracts Evaluated by Four in vitro Methods. J. Biol. Sci. 2010, 8, 260–266. [Google Scholar]
- Kaneria, M.; Chanda, S. Evaluation of antioxidant and antimicrobial properties of Manilkara zapota L.(chiku) leaves by sequential soxhlet extraction method. Asian Pac. J. Trop. Biomed. 2012, 2, S1526–S1533. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Bueno, P.C.D.S.; Delazari, D.S.; Guiguer, E.L.; Coqueiro, D.P.; Araújo, A.C.; de Souza, M.D.S.S.; Farinazzi-Machado, F.M.V.; Mendes, C.G.; Groppo, M. Antidiabetic and Antilipidemic Effects of Manilkara zapota. J. Med. Food 2015, 18, 385–391. [Google Scholar] [CrossRef]
- Paul, S.R.; Hakim, M.L. In vivo hypoglycemic study of Manilkara zapota leaf and seed extracts. Bangladesh J. Pharmacol. 2015, 10, 246–250. [Google Scholar] [CrossRef]
- Ma, J.; Luo, X.D.; Protiva, P.; Yang, H.; Ma, C.; Basile, M.J.; Weinstein, I.B.; Kennelly, E.J. Bioactive novel polyphenols from the fruit of Manilkara zapota (Sapodilla). J. Nat. Prod. 2003, 66, 983–986. [Google Scholar] [CrossRef]
- Fayek, N.M.; Monem, A.R.A.; Mossa, M.Y.; Meselhy, M.R.; Shazly, A.H. Chemical and biological study of Manilkara zapota (L.) Van Royen leaves (Sapotaceae) cultivated in Egypt. Pharmacogn. Res. 2012, 4, 85–91. [Google Scholar]
- Tanko, H.; Carrier, D.J.; Duan, L.; Clausen, E. Pre- and post-harvest processing of medicinal plants. Plant Genet. Resour. 2005, 3, 304–313. [Google Scholar] [CrossRef]
- Schulze, E.D.; Lange, O.L.; Kappen, L.; Buschbom, U.; Evenari, M. Stomatal responses to changes in temperature at increasing water stress. Planta 1973, 110, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrauri, J.A.; Rupérez, P.; Saura-Calixto, F. Effect of Drying Temperature on the Stability of Polyphenols and Antioxidant Activity of Red Grape Pomace Peels. J. Agric. Food Chem. 1997, 45, 1390–1393. [Google Scholar] [CrossRef]
- Yu, J. Thermal stability of major classes of polyphenols in skins, seeds and stems of grape pomace. In Grapes: Production, Phenolic Composition and Potential Biomedical Effects; de Sousa Câmara, J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2014; pp. 273–285. ISBN 9781633214101. [Google Scholar]
- Volf, I.; Ignat, I.; Neamtu, M.; Popa, V.I. Thermal stability, antioxidant activity, and photo-oxidation of natural polyphenols. Chem. Pap. 2014, 68, 121–129. [Google Scholar] [CrossRef]
- Terán-Hilares, R.; Chirinos, R.; Pedreschi, R.; Campos, D. Enhanced antioxidant properties of tara (Caesalpinia spinosa) gallotannins by thermal hydrolysis and its synergistic effects with α-tocopherol, ascorbyl palmitate, and citric acid on sacha inchi (Plukenetia volubilis) oil. J. Food Process Eng. 2018, 41, e12613. [Google Scholar] [CrossRef]
- Elhamirad, A.H.; Zamanipoor, M.H. Thermal stability of some flavonoids and phenolic acids in sheep tallow olein. Eur. J. Lipid Sci. Technol. 2012, 114, 602–606. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Howard, L.R. Effects of solvent and temperature on pressurized liquid extraction of anthocyanins and total phenolics from dried red grape skin. J. Agric. Food Chem. 2003, 51, 5207–5213. [Google Scholar] [CrossRef]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Trygg, J.; Wikström, C.; Wold, S. Multi-and Megavariate Data Analysis; Umetrics AB: Umeá, Sweden, 2006. [Google Scholar]
- Zafrilla, P.; Ferreres, F.; Tomás-Barberán, F.A. Effect of processing and storage on the antioxidant ellagic acid derivatives and flavonoids of red raspberry (Rubus idaeus) jams. J. Agric. Food Chem. 2001, 49, 3651–3655. [Google Scholar] [CrossRef]
- Khatib, A.; Wilson, E.G.; Kim, H.K.; Lefeber, A.W.M.; Erkelens, C.; Choi, Y.H.; Verpoorte, R. Application of two-dimensional J-resolved nuclear magnetic resonance spectroscopy to differentiation of beer. Anal. Chim. Acta 2006, 559, 264–270. [Google Scholar] [CrossRef]
- Mahrous, E.A.; Farag, M.A. Two dimensional NMR spectroscopic approaches for exploring plant metabolome: A review. J. Adv. Res. 2015, 6, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drying Temperature (°C) | Water Loss (%) a | Extraction Yield (%) b | Total Phenol Content (mg/g) b | DPPH Antioxidant Activity EC50 (µg/mL) b |
|---|---|---|---|---|
| 25 | 48.94 ± 9.37 | 3.67 ± 0.03 | 0.57 ± 0.003 | 26.41 ± 0.180 |
| 50 | 26.71 ± 3.61 | 2.88 ± 0.02 | 0.24 ± 0.004 | 22.73 ± 0.050 |
| 75 | 39.25 ± 2.00 | 1.90 ± 0.02 | 1.21 ± 0.004 | 17.31 ± 0.010 |
| 100 | 57.81 ± 1.48 | 1.41 ± 0.03 | 2.59 ± 0.004 | 11.04 ± 0.010 |
| C. sinensis | - | - | 2.96 ± 0.020 | 9.22 ± 0.060 |
| Gallic acid | - | - | - | 2.18 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Bolio, G.I.; Dzul-Romero, R.E.; Maldonado Velázquez, M.G.; Zamora Cresencio, P.; Hernández-Núñez, E.; Aguirre-Crespo, F.J. The Influence of Drying Temperatures on the Metabolic Profiles and Antioxidant Activity of Manilkara zapota Leaves. Metabolites 2019, 9, 217. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100217
Hernández-Bolio GI, Dzul-Romero RE, Maldonado Velázquez MG, Zamora Cresencio P, Hernández-Núñez E, Aguirre-Crespo FJ. The Influence of Drying Temperatures on the Metabolic Profiles and Antioxidant Activity of Manilkara zapota Leaves. Metabolites. 2019; 9(10):217. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100217
Chicago/Turabian StyleHernández-Bolio, Gloria I., Rubí E. Dzul-Romero, María G. Maldonado Velázquez, Pedro Zamora Cresencio, Emanuel Hernández-Núñez, and Francisco J. Aguirre-Crespo. 2019. "The Influence of Drying Temperatures on the Metabolic Profiles and Antioxidant Activity of Manilkara zapota Leaves" Metabolites 9, no. 10: 217. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100217