Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling


Abstract
:1. Introduction
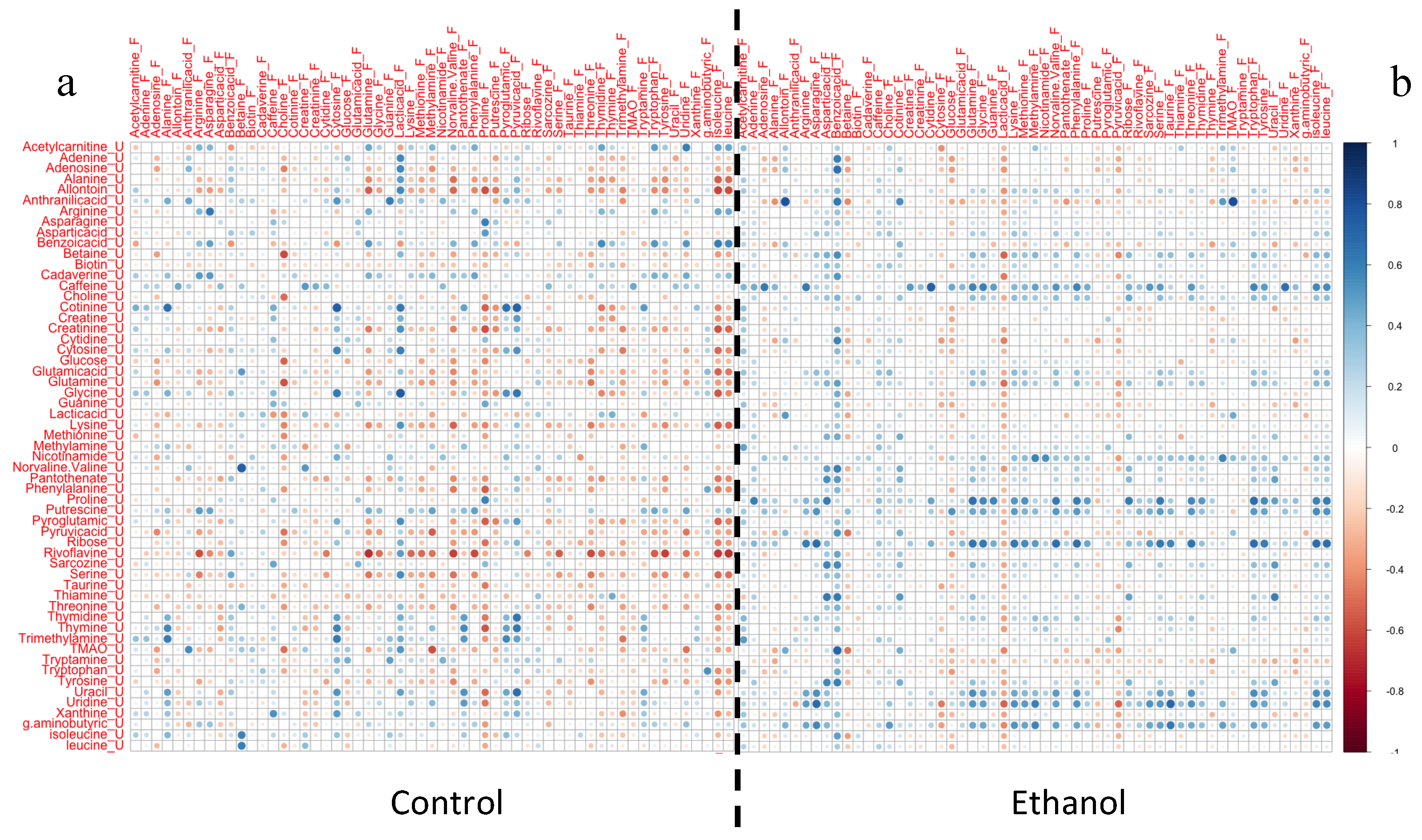
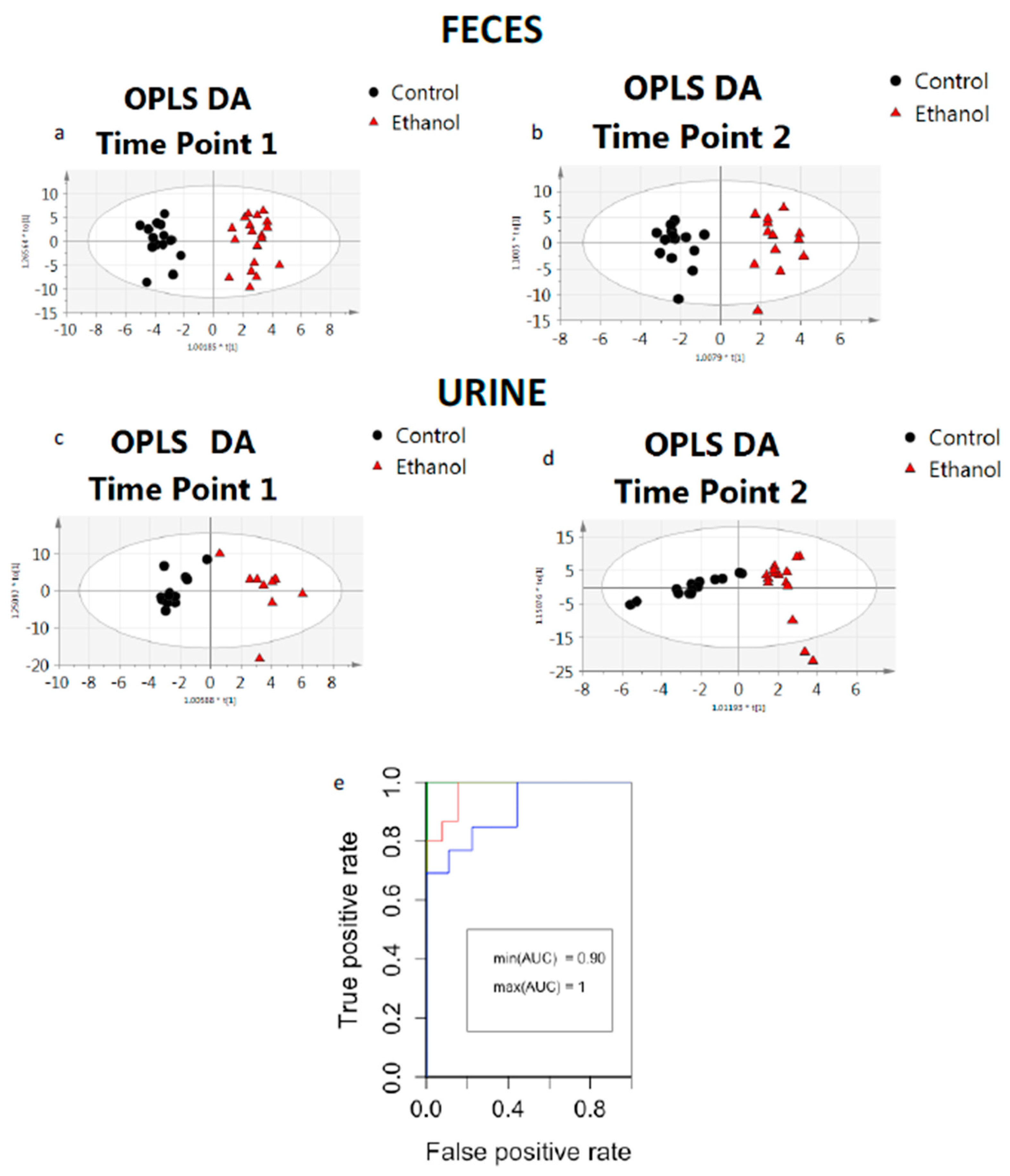
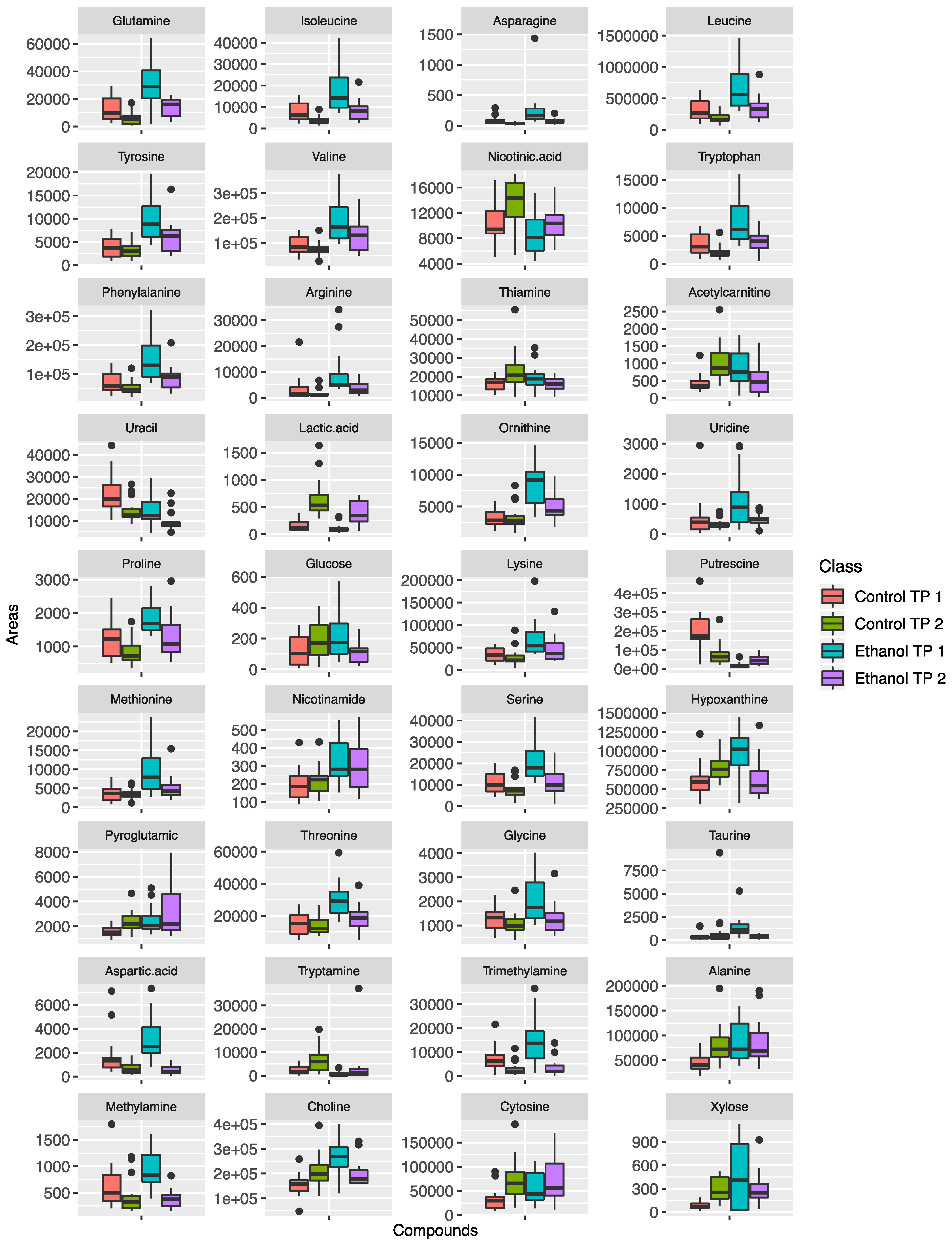
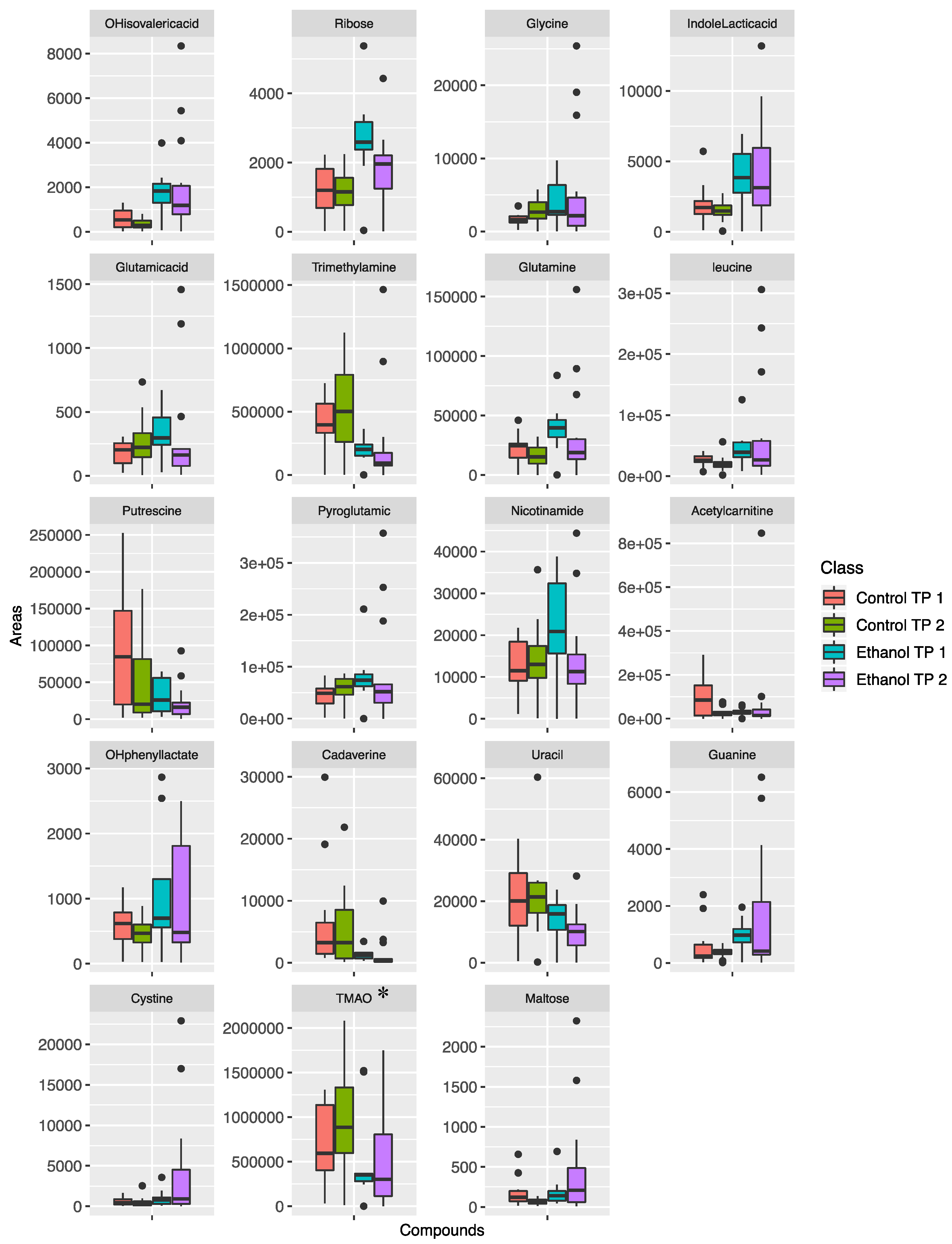
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. In vivo Study
4.3. Sample Preparation of Fecal and Urine Samples
4.4. LC-MS/MS analysis
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farooq, M.O.; Bataller, R. Pathogenesis and Management of Alcoholic Liver Disease. DDI 2016, 34, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.K.; Patterson, A.D.; Yang, Q.; Krausz, K.W.; Li, H.; Idle, J.R.; Fornace, A.J.; Gonzalez, F.J. Identification of noninvasive biomarkers for alcohol-induced liver disease using urinary metabolomics and the Ppara-null mouse. J. Proteome Res. 2010, 9, 4176–4188. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wei, X.; Yin, X.; Wang, Y.; Zhang, M.; Zhao, C.; Zhao, H.; McClain, C.J.; Feng, W.; Zhang, X. Hepatic and Fecal Metabolomic Analysis of the Effects of Lactobacillus rhamnosus GG on Alcoholic Fatty Liver Disease in Mice. J. Proteome Res. 2015, 14, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Theodoridis, G.; Gika, H.G.; Wilson, I.D. Mass spectrometry-based holistic analytical approaches for metabolite profiling in systems biology studies. Mass Spectrom. Rev. 2011, 30, 884–906. [Google Scholar] [CrossRef]
- Xie, G.; Zhong, W.; Zheng, X.; Li, Q.; Qiu, Y.; Li, H.; Chen, H.; Zhou, Z.; Jia, W. Chronic Ethanol Consumption Alters Mammalian Gastrointestinal Content Metabolites. J. Proteome Res. 2013, 12, 3297–3306. [Google Scholar] [CrossRef] [Green Version]
- Gika, H.G.; Wilson, I.D. Global metabolic profiling for the study of alcohol-related disorders. Bioanalysis 2014, 6, 59–77. [Google Scholar] [CrossRef]
- Bradford, B.U.; O’Connell, T.M.; Han, J.; Kosyk, O.; Shymonyak, S.; Ross, P.K.; Winnike, J.; Kono, H.; Rusyn, I. Metabolomic profiling of a modified alcohol liquid diet model for liver injury in the mouse uncovers new markers of disease. Toxicol. Appl. Pharmacol. 2008, 232, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, P.C.; Kim, D.; Crews, F.T.; Macdonald, J.M. 1H NMR-Based Metabolomic Analysis of Liver, Serum, and Brain Following Ethanol Administration in Rats. Chem. Res. Toxicol. 2008, 21, 408–420. [Google Scholar] [CrossRef]
- Rachakonda, V.; Gabbert, C.; Raina, A.; Bell, L.N.; Cooper, S.; Malik, S.; Behari, J. Serum Metabolomic Profiling in Acute Alcoholic Hepatitis Identifies Multiple Dysregulated Pathways. PLoS ONE 2014, 9, e113860. [Google Scholar] [CrossRef]
- Irwin, C.; van Reenen, M.; Mason, S.; Mienie, L.J.; Wevers, R.A.; Westerhuis, J.A.; Reinecke, C.J. The 1H-NMR-based metabolite profile of acute alcohol consumption: A metabolomics intervention study. PLoS ONE 2018, 13, e0196850. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Wang, C.; Li, B.; Zhang, A. Metabolomics of alcoholic liver disease: A clinical discovery study. RSC Adv. 2015, 5, 80381–80387. [Google Scholar] [CrossRef]
- Jain, A.; Li, X.H.; Chen, W.N. An untargeted fecal and urine metabolomics analysis of the interplay between the gut microbiome, diet and human metabolism in Indian and Chinese adults. Sci. Rep. 2019, 9, 9191. [Google Scholar] [CrossRef] [PubMed]
- Couch, R.D.; Dailey, A.; Zaidi, F.; Navarro, K.; Forsyth, C.B.; Mutlu, E.; Engen, P.A.; Keshavarzian, A. Alcohol Induced Alterations to the Human Fecal VOC Metabolome. PLoS ONE 2015, 10, e0119362. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, P.; Seebauer, C.T.; Schnabl, B. Alcoholic Liver Disease: The Gut Microbiome and Liver Cross Talk. Alcohol. Clin. Exp. Res. 2015, 39, 763–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutlu, E.; Keshavarzian, A.; Engen, P.; Forsyth, C.B.; Sikaroodi, M.; Gillevet, P. Intestinal dysbiosis: A possible mechanism of alcohol-induced endotoxemia and alcoholic steatohepatitis in rats. Alcohol. Clin. Exp. Res. 2009, 33, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhao, A.; Zhou, M.; Lin, J.; Qiu, Y.; Su, M.; Jia, W. GC/MS-based urinary metabolomics reveals systematic differences in metabolism and ethanol response between Sprague–Dawley and Wistar rats. Metabolomics 2011, 7, 363–374. [Google Scholar] [CrossRef]
- Shi, X.; Yao, D.; Chen, C. Identification of N-Acetyltaurine as a Novel Metabolite of Ethanol through Metabolomics-guided Biochemical Analysis. J. Biol. Chem. 2012, 287, 6336–6349. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, S.; Matamoros, S.; Cani, P.D.; Neyrinck, A.M.; Jamar, F.; Stärkel, P.; Windey, K.; Tremaroli, V.; Bäckhed, F.; Verbeke, K.; et al. Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity. Proc. Natl. Acad. Sci. USA 2014, 111, E4485–E4493. [Google Scholar] [CrossRef] [Green Version]
- Engen, P.A.; Green, S.J.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. The Gastrointestinal Microbiome. Alcohol Res. 2015, 37, 223–236. [Google Scholar]
- Cassard, A.-M.; Ciocan, D. Microbiota, a key player in alcoholic liver disease. Clin. Mol. Hepatol. 2018, 24, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirpich, I.A.; Petrosino, J.; Ajami, N.; Feng, W.; Wang, Y.; Liu, Y.; Beier, J.I.; Barve, S.S.; Yin, X.; Wei, X.; et al. Saturated and Unsaturated Dietary Fats Differentially Modulate Ethanol-Induced Changes in Gut Microbiome and Metabolome in a Mouse Model of Alcoholic Liver Disease. Am. J. Pathol. 2016, 186, 765–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Zhou, Z. Alterations of the gut microbiome and metabolome in alcoholic liver disease. World J. Gastrointest. Pathophysiol. 2014, 5, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, Q.; Guo, L.; Zeng, H.; Ding, C.; Zhang, W.; Xu, D.; Wang, X.; Qiu, J.; Dong, Q.; et al. Gut Microbiota and Relevant Metabolites Analysis in Alcohol Dependent Mice. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Rachamin, G.; Macdonald, J.A.; Wahid, S.; Clapp, J.J.; Khanna, J.M.; Israel, Y. Modulation of alcohol dehydrogenase and ethanol metabolism by sex hormones in the spontaneously hypertensive rat. Effect of chronic ethanol administration. Biochem. J. 1980, 186, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, K.; Malmström, K.K. Sex differencds in consumption and elimination of alcohol in albino rats. Ann. Med. Exp. Biol. Fenn. 1967, 45, 389–392. [Google Scholar]
- Duan, J.; Esberg, L.B.; Ye, G.; Borgerding, A.J.; Ren, B.H.; Aberle, N.S.; Epstein, P.N.; Ren, J. Influence of gender on ethanol-induced ventricular myocyte contractile depression in transgenic mice with cardiac overexpression of alcohol dehydrogenase. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 134, 607–614. [Google Scholar] [CrossRef]
- Middaugh, L.D.; Frackelton, W.F.; Boggan, W.O.; Onofrio, A.; Shepherd, C.L. Gender differences in the effects of ethanol on C57BL/6 mice. Alcohol 1992, 9, 257–260. [Google Scholar] [CrossRef]
- Thomasson, H.R. Gender Differences in Alcohol Metabolism. In Recent Developments in Alcoholism: Alcoholism and Women; Galanter, M., Begleiter, H., Deitrich, R., Gallant, D., Goodwin, D., Gottheil, E., Paredes, A., Rothschild, M., Van Thiel, D., Edwards, H., Eds.; Recent Developments in Alcoholism; Springer: Boston, MA, USA, 1995; pp. 163–179. [Google Scholar]
- Kono, H.; Wheeler, M.D.; Rusyn, I.; Lin, M.; Seabra, V.; Rivera, C.A.; Bradford, B.U.; Forman, D.T.; Thurman, R.G. Gender differences in early alcohol-induced liver injury: Role of CD14, NF-kappaB, and TNF-alpha. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G652–G661. [Google Scholar] [CrossRef]
- Nanji, A.A.; Jokelainen, K.; Fotouhinia, M.; Rahemtulla, A.; Thomas, P.; Tipoe, G.L.; Su, G.L.; Dannenberg, A.J. Increased severity of alcoholic liver injury in female rats: Role of oxidative stress, endotoxin, and chemokines. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1348–G1356. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Mathews, S.; Ki, S.H.; Wang, H.; Gao, B. Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nat. Protoc. 2013, 8, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazier, T.H.; Stocker, A.M.; Kershner, N.A.; Marsano, L.S.; McClain, C.J. Treatment of alcoholic liver disease. Ther. Adv. Gastroenterol. 2011, 4, 63–81. [Google Scholar] [CrossRef]
- Becker, H.C.; Ron, D. Animal models of excessive alcohol consumption: Recent advances and future challenges. Alcohol 2014, 48, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Gika, H.G.; Ji, C.; Theodoridis, G.A.; Michopoulos, F.; Kaplowitz, N.; Wilson, I.D. Investigation of chronic alcohol consumption in rodents via ultra-high-performance liquid chromatography-mass spectrometry based metabolite profiling. J. Chromatogr. A 2012, 1259, 128–137. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; Liang, X.; Roberts, M.S. Chapter 30—Hepatic Metabolism in Liver Health and Disease. In Liver Pathophysiology; Muriel, P., Ed.; Academic Press: Boston, MA, USA, 2017; pp. 391–400. [Google Scholar]
- Yamamoto, T.; Moriwaki, Y.; Takahashi, S. Effect of ethanol on metabolism of purine bases (hypoxanthine, xanthine, and uric acid). Clin. Chim. Acta 2005, 356, 35–57. [Google Scholar] [CrossRef]
- Afzali, A.; Weiss, N.S.; Boyko, E.J.; Ioannou, G.N. Association between serum uric acid level and chronic liver disease in the United States. Hepatology 2010, 52, 578–589. [Google Scholar] [CrossRef]
- Loguercio, C.; Blanco, F.D.; De Girolamo, V.; Disalvo, D.; Nardi, G.; Parente, A.; Blanco, C.D. Ethanol consumption, amino acid and glutathione blood levels in patients with and without chronic liver disease. Alcohol. Clin. Exp. Res. 1999, 23, 1780–1784. [Google Scholar] [CrossRef]
- Cederbaum, A.I. ALCOHOL METABOLISM. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef]
- Malaguarnera, G.; Giordano, M.; Nunnari, G.; Bertino, G.; Malaguarnera, M. Gut microbiota in alcoholic liver disease: Pathogenetic role and therapeutic perspectives. World J. Gastroenterol. 2014, 20, 16639–16648. [Google Scholar] [CrossRef] [PubMed]
- Thurman, R.G. Hepatic alcohol oxidation and its metabolic liability. Fed. Proc. 1977, 36, 1640–1646. [Google Scholar] [PubMed]
- Fernando, H.; Bhopale, K.K.; Kondraganti, S.; Kaphalia, B.S.; Ansari, G.A.S. Lipidomic Changes in Rat Liver after Long-Term Exposure to Ethanol. Toxicol Appl. Pharmacol. 2011, 255, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hoek, J.B.; Cahill, A.; Pastorino, J.G. Alcohol and Mitochondria: A Dysfunctional Relationship. Gastroenterology 2002, 122, 2049–2063. [Google Scholar] [CrossRef]
- Lieberman, M.; Peet, A. Mark’s Basic Medical Biochemistry, 4th ed.; North American edition; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2012. [Google Scholar]
- Seitz, H.K.; Gärtner, U.; Egerer, G.; Simanowski, U.A. Ethanol metabolism in the gastrointestinal tract and its possible consequences. Alcohol Alcohol. Suppl. 1994, 2, 157–162. [Google Scholar]
- Loftus, N.; Barnes, A.; Ashton, S.; Michopoulos, F.; Theodoridis, G.; Wilson, I.; Ji, C.; Kaplowitz, N. Metabonomic Investigation of Liver Profiles of Nonpolar Metabolites Obtained from Alcohol-Dosed Rats and Mice Using High Mass Accuracy MSn Analysis. J. Proteome Res. 2011, 10, 705–713. [Google Scholar] [CrossRef]
- Manna, S.K.; Patterson, A.D.; Yang, Q.; Krausz, K.W.; Idle, J.R.; Fornace, A.J.; Gonzalez, F.J. UPLC–MS-based Urine Metabolomics Reveals Indole-3-lactic Acid and Phenyllactic Acid as Conserved Biomarkers for Alcohol-induced Liver Disease in the Ppara-null Mouse Model. J. Proteome Res. 2011, 10, 4120–4133. [Google Scholar] [CrossRef]
- Jiménez-Girón, A.; Ibáñez, C.; Cifuentes, A.; Simó, C.; Muñoz-González, I.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Faecal metabolomic fingerprint after moderate consumption of red wine by healthy subjects. J. Proteome Res. 2015, 14, 897–905. [Google Scholar] [CrossRef]
- Virgiliou, C.; Sampsonidis, I.; Gika, H.G.; Raikos, N.; Theodoridis, G.A. Development and validation of a HILIC-MS/MS multitargeted method for metabolomics applications. Electrophoresis 2015, 36, 2215–2225. [Google Scholar] [CrossRef]
- Deda, O.; Gika, H.; Panagoulis, T.; Taitzoglou, I.; Raikos, N.; Theodoridis, G. Impact of exercise on fecal and cecal metabolome over aging: A longitudinal study in rats. Bioanalysis 2017, 9, 21–36. [Google Scholar] [CrossRef]
- Begou, O.; Deda, O.; Agapiou, A.; Taitzoglou, I.; Gika, H.; Theodoridis, G. Urine and fecal samples targeted metabolomics of carobs treated rats. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1114, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Spyrelli, E.D.; Kyriazou, A.V.; Virgiliou, C.; Nakas, A.; Deda, O.; Papageorgiou, V.P.; Assimopoulou, A.N.; Gika, H.G. Metabolic profiling study of shikonin’s cytotoxic activity in the Huh7 human hepatoma cell line. Mol. Biosyst. 2017, 13, 841–851. [Google Scholar] [CrossRef]
- Gika, H.G.; Zisi, C.; Theodoridis, G.; Wilson, I.D. Protocol for quality control in metabolic profiling of biological fluids by U(H)PLC-MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1008, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Statistics of the Model | Predictive Ability | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Apred | Aorth | R2Y | R2X | Q2YCV | p (CV ANOVA) | Sensitivity | Specificity | Accuracy | AUC | |
| Fecal samples Control vs. Ethanol TP 1 | 1 | 2 | 0.942 | 0.567 | 0.942 | 2.84 × 10−8 | 1 | 1 | 100% | 1 |
| Fecal samples Control vs. Ethanol TP 1 (Male) | 1 | 2 | 0.998 | 0.768 | 0.967 | 7.60 × 10−5 | 1 | 1 | 100% | 1 |
| Fecal samples Control vs. Ethanol TP 1 (Female) | 1 | 2 | 0.955 | 0.583 | 0.836 | 9.92 × 10−5 | 1 | 1 | 100% | 1 |
| Fecal samples Control vs. Ethanol TP 2 | 1 | 2 | 0.925 | 0.481 | 0.925 | 3.00 × 10−4 | 1 | 0.92 | 96% | 0.99 |
| Fecal samples Control vs. Ethanol TP 2 (Male) | 1 | 4 | 0.982 | 0.57 | 0.8 | 1.00 × 10−2 | 1 | 1 | 100% | 1 |
| Fecal samples Control vs. Ethanol TP 2 (Female) | - | - | - | - | - | >1 | - | - | - | - |
| Urine samples Control vs. Ethanol TP 1 | 1 | 2 | 0.791 | 0.671 | 0.791 | 9.70 × 10−2 | 0.76 | 0.88 | 81% | 0.9 |
| Urine samples Control vs. Ethanol TP 1 (Male) | 1 | 2 | 0.998 | 0.657 | 0.796 | 1.00 × 10−2 | 1 | 1 | 100% | 1 |
| Urine samples Control vs. Ethanol TP 1 (Female) | - | - | - | - | - | >1 | - | - | - | - |
| Urine samples Control vs. Ethanol TP 2 | 1 | 1 | 0.877 | 0.625 | 0.877 | 4.93 × 10−6 | 1 | 0.93 | 96% | 0.99 |
| Urine samples Control vs. Ethanol TP 2 (Male) | - | - | - | - | - | >1 | - | - | - | - |
| Urine samples Control vs. Ethanol TP 2 (Female) | 1 | 1 | 0.822 | 0.633 | 0.708 | 9.00 × 10−3 | 1 | 1 | 100% | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deda, O.; Virgiliou, C.; Orfanidis, A.; Gika, H.G. Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling. Metabolites 2019, 9, 232. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100232
Deda O, Virgiliou C, Orfanidis A, Gika HG. Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling. Metabolites. 2019; 9(10):232. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100232
Chicago/Turabian StyleDeda, Olga, Christina Virgiliou, Amvrosios Orfanidis, and Helen G. Gika. 2019. "Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling" Metabolites 9, no. 10: 232. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo9100232