Optimization of Polyhydroxybutyrate Production by Amazonian Microalga Stigeoclonium sp. B23

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization and Identification of B23 Strain
2.2. 18S rRNA Gene Sequence Analysis
2.3. Media and Growth Conditions
2.4. Chlorophyll a Content
2.5. Fluorescent Microscopy
2.6. Quantification of PHB Fluorescence
2.7. PHB Extraction
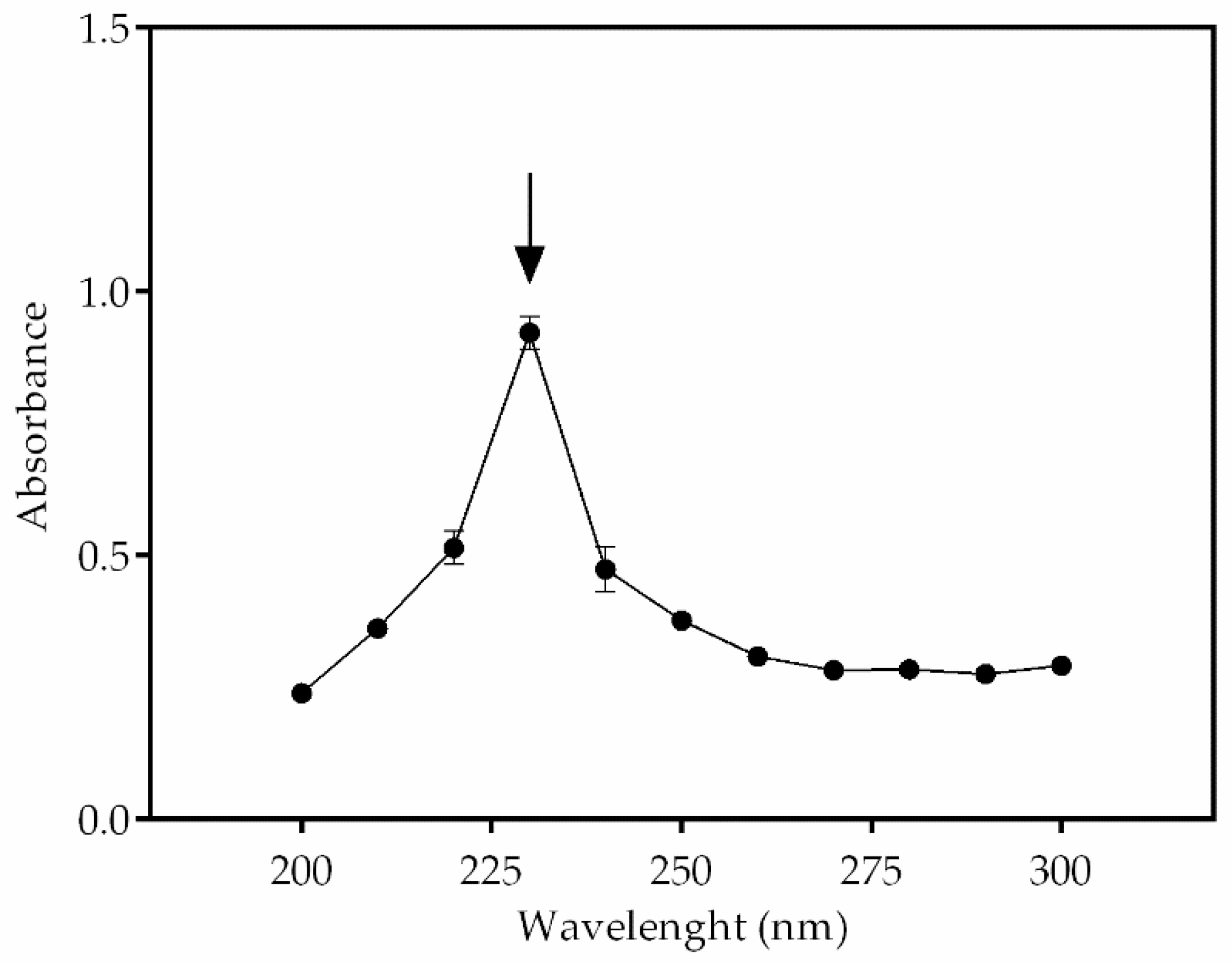
2.8. Quantification of PHB Content
2.9. Statistical Analysis
3. Results
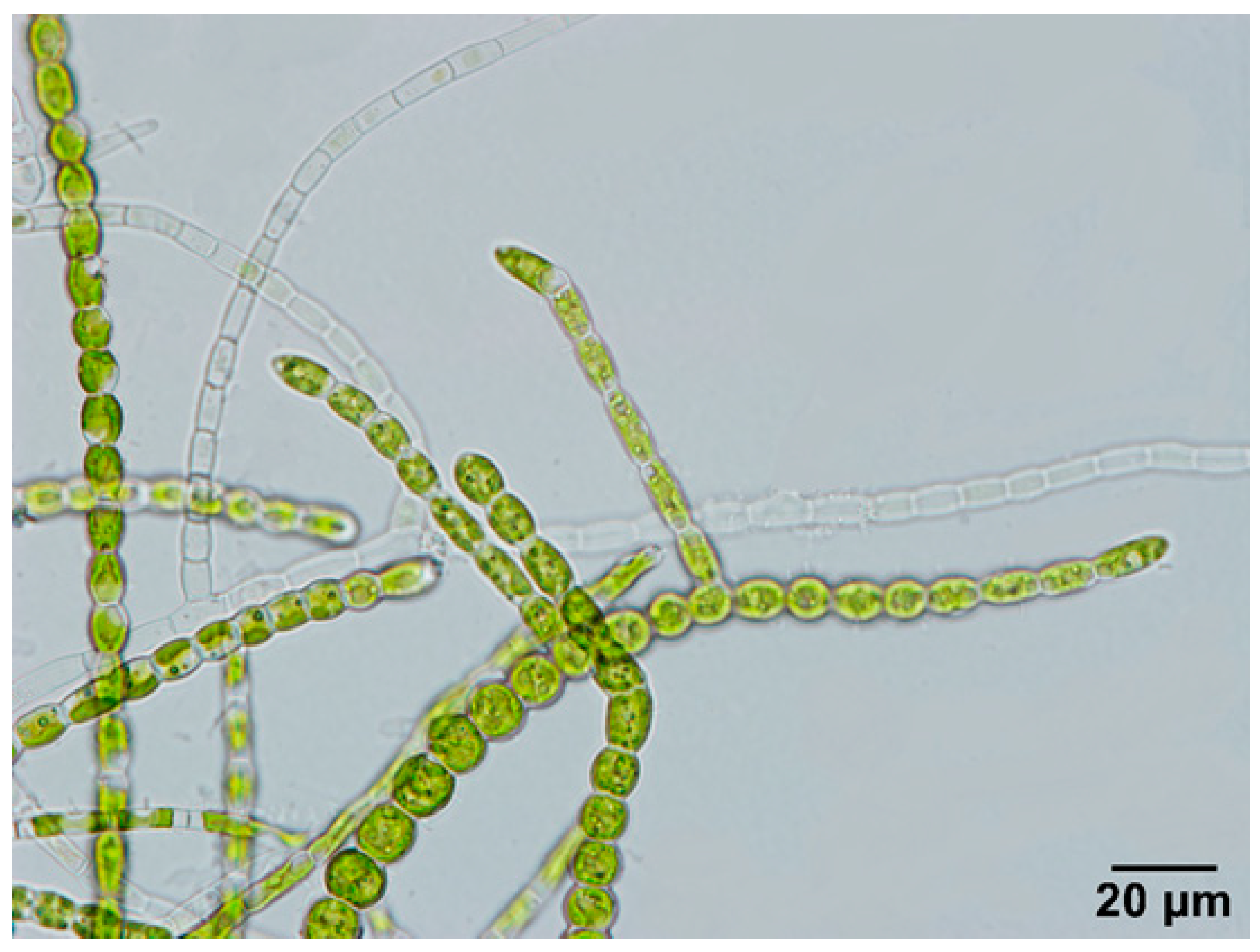
3.1. Morphological and Molecular Identification of Strain B23
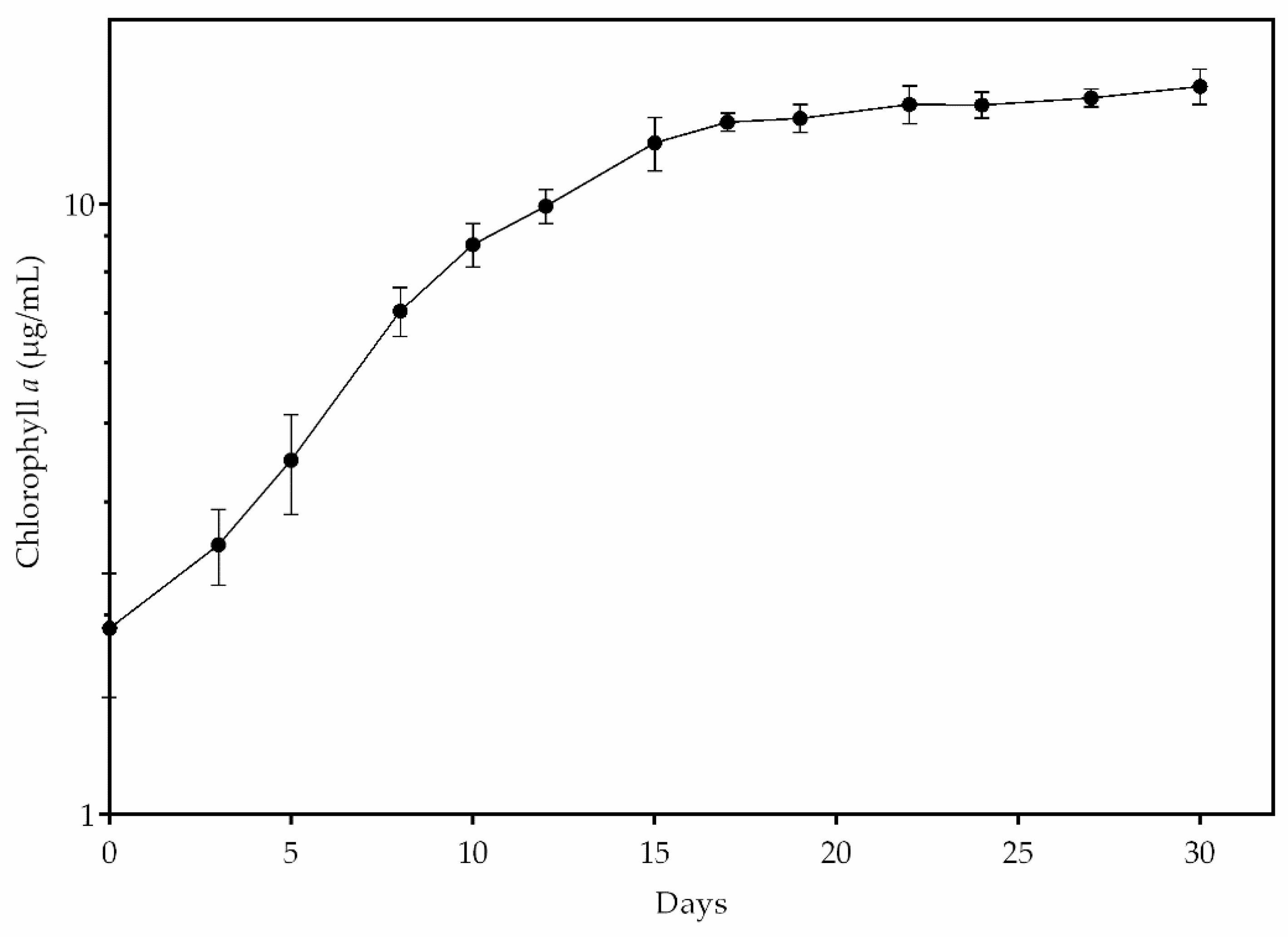
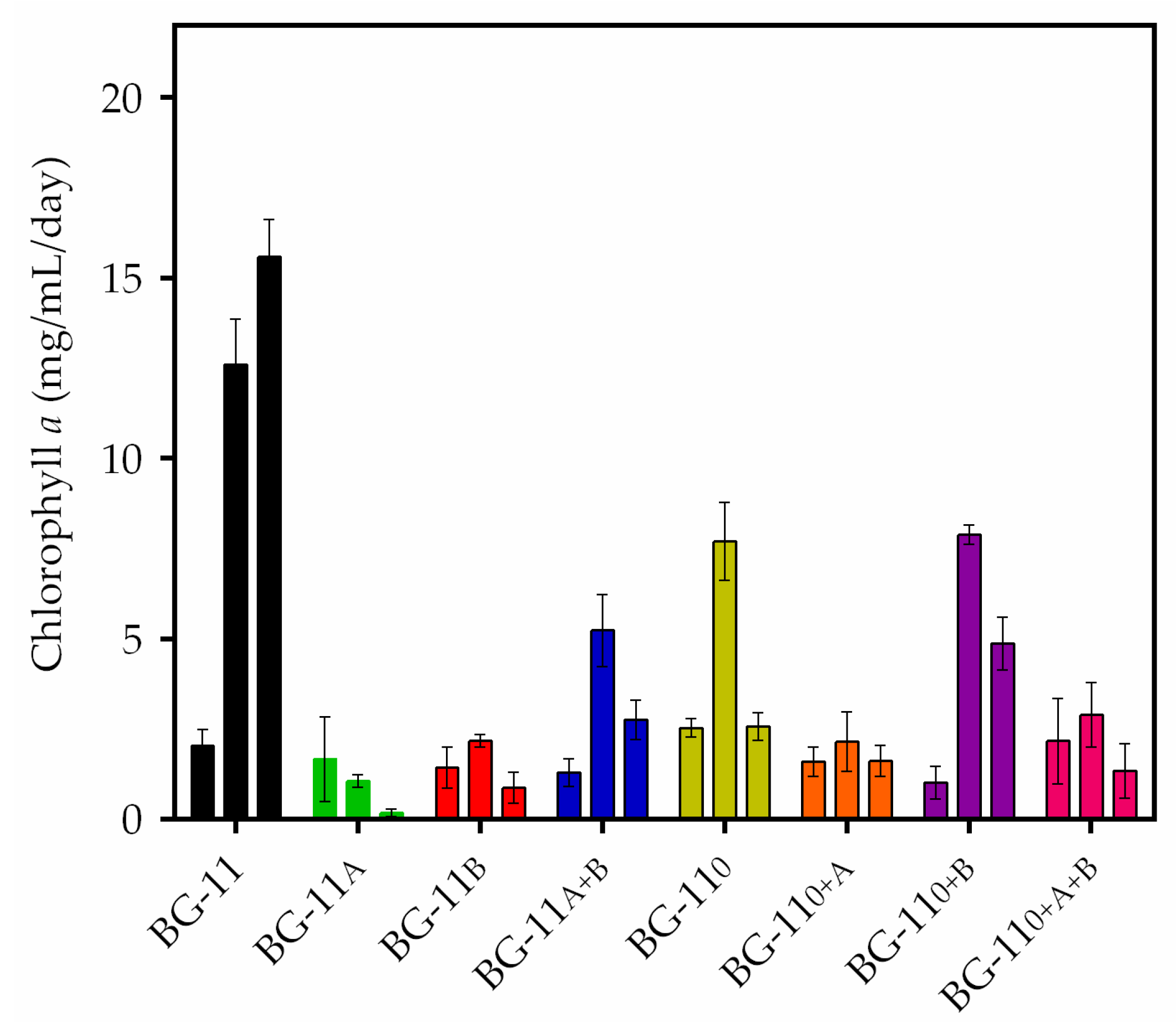
3.2. Growth of Stigeoclonium sp. B23
3.3. Growth of Stigeoclonium sp. B23
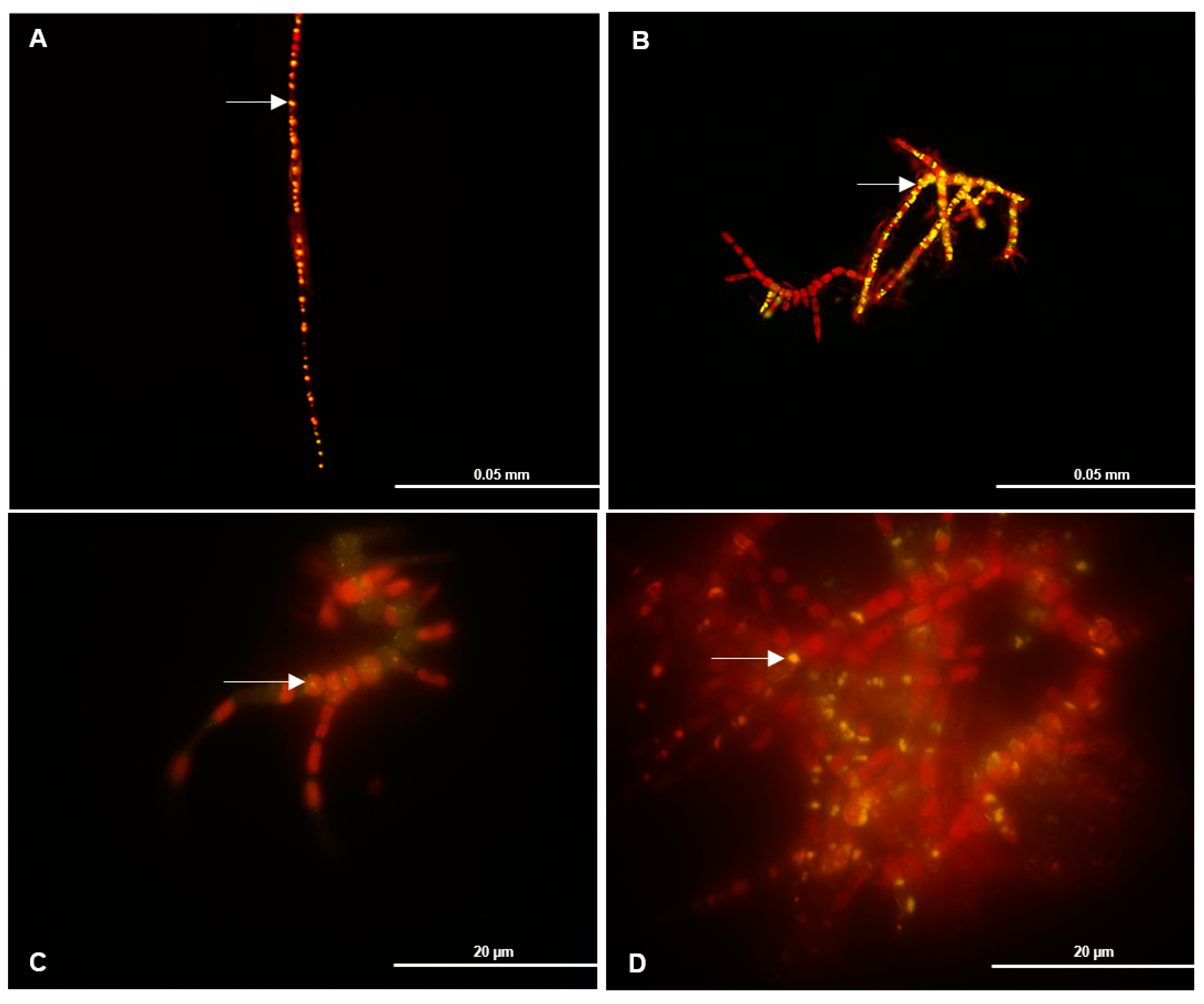
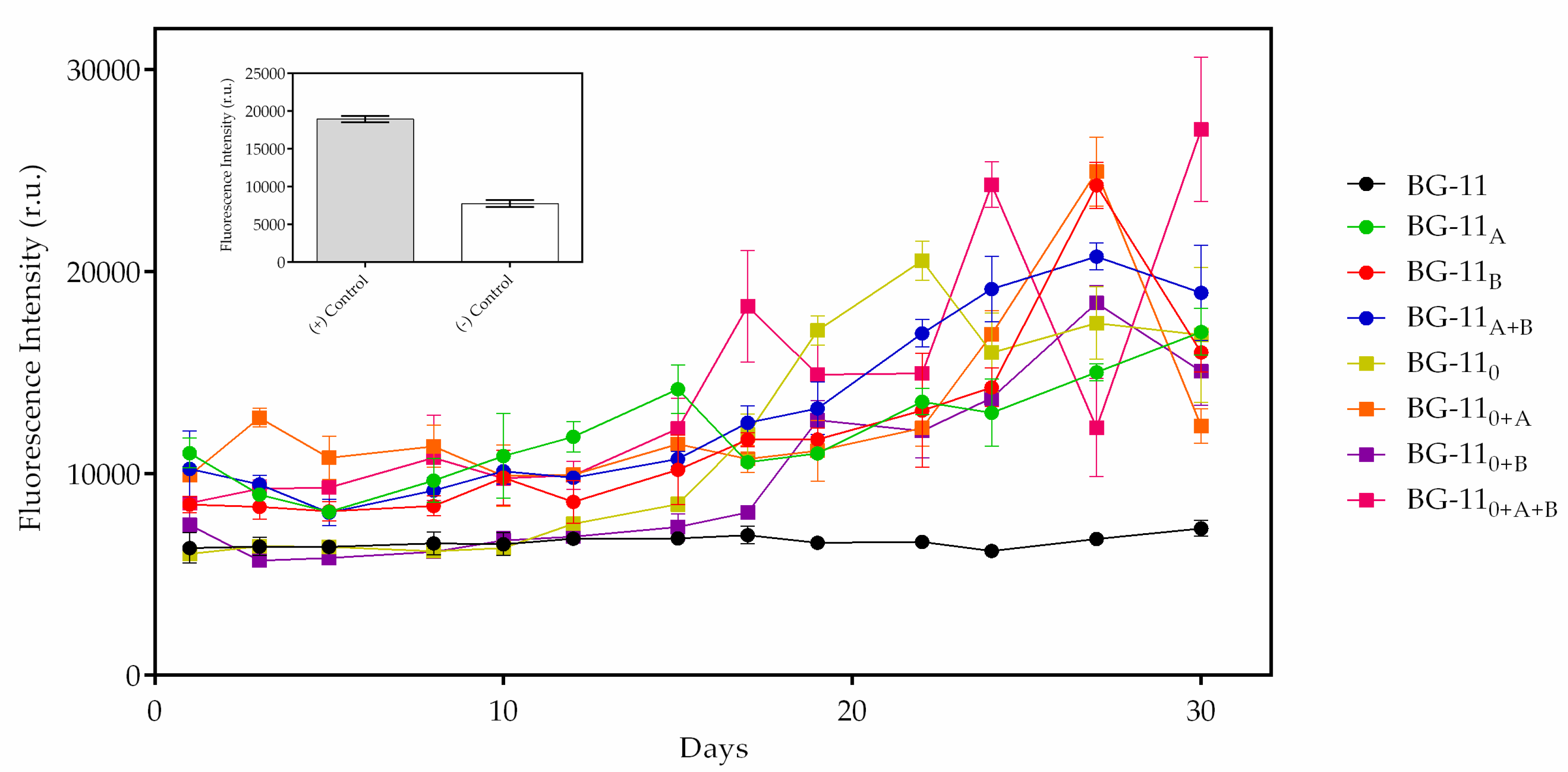
3.4. PHB Detection by Fluorescence
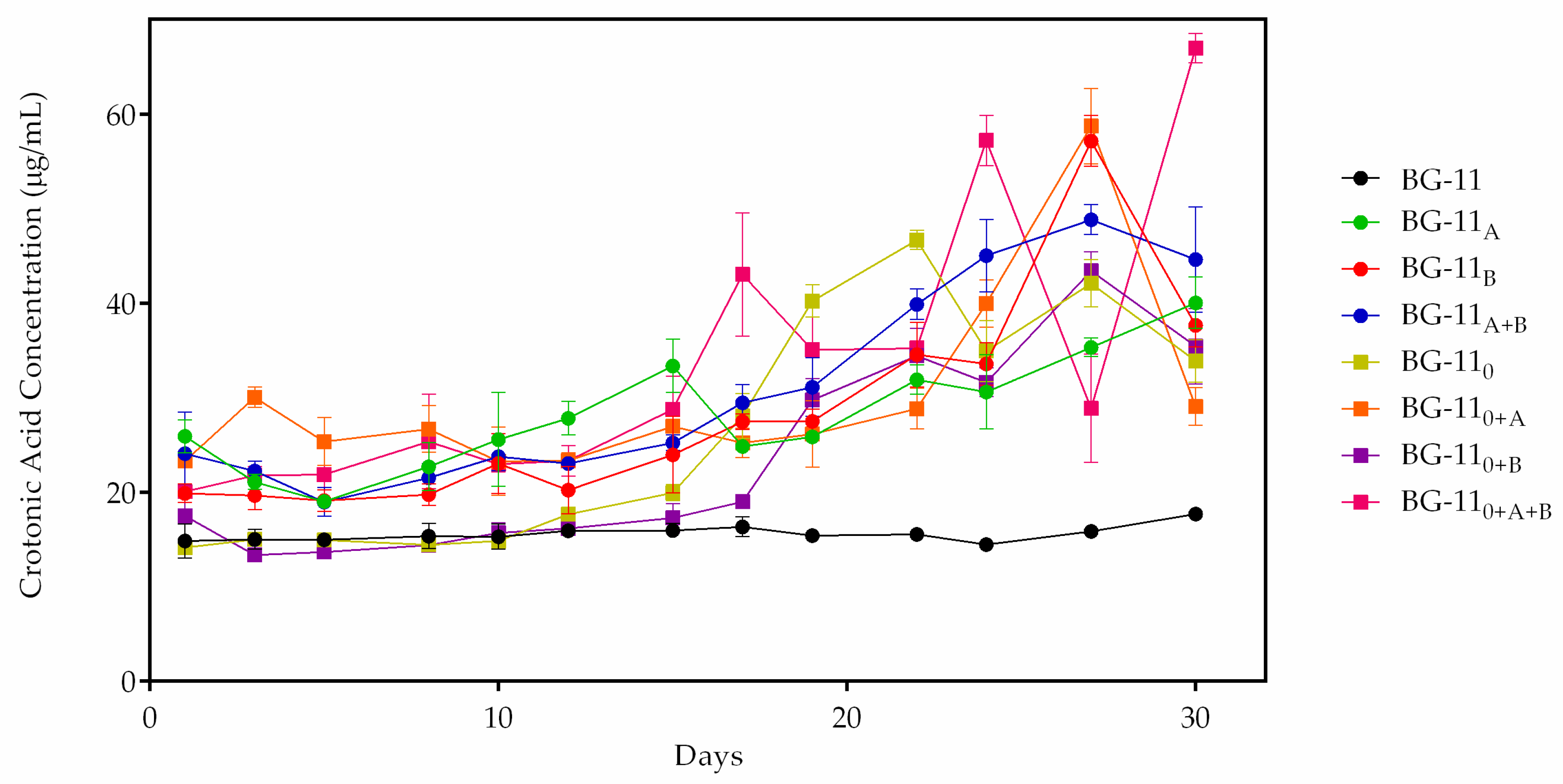
3.5. PHB and Fluorescence Correlation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doi, Y.; Kanesawa, Y.; Tanahashi, N.; Kumagai, Y. Biodegradation of microbial polyesters in the marine environment. Polym. Degrad. Stab. 1992, 36, 173–177. [Google Scholar] [CrossRef]
- Lee, S.Y.; Chang, H.N. High cell density cultivation of Escherichia coli W using sucrose as a carbon source. Biotechnol. Lett. 1993, 15, 971–974. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Plastics from bacteria and for bacteria: Poly(beta-hydroxyalkanoates) as natural, biocompatible, and biodegradable polyesters. Adv. Biochem. Eng. Biotechnol. 1990, 41, 77–93. [Google Scholar] [CrossRef]
- Gomes Gradíssimo, D.; Pereira Xavier, L.; Valadares Santos, A. Cyanobacterial Polyhydroxyalkanoates: A Sustainable Alternative in Circular Economy. Molecules 2020, 25, 4331. [Google Scholar] [CrossRef]
- Holmes, P.A. Applications of PHB—A microbially produced biodegradable thermoplastic. Phys. Technol. 1985, 16, 32–36. [Google Scholar] [CrossRef]
- Lee, S.Y. Plastic bacteria? Progress and prospects for polyhydroxyalkanoate production in bacteria. Trends Biotechnol. 1996, 14, 431–438. [Google Scholar] [CrossRef]
- Kavitha, G.; Kurinjimalar, C.; Sivakumar, K.; Kaarthik, M.; Aravind, R.; Palani, P.; Rengasamy, R. Optimization of polyhydroxybutyrate production utilizing waste water as nutrient source by Botryococcus braunii Kütz using response surface methodology. Int. J. Biol. Macromol. 2016, 93, 534–542. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol. Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef]
- Cassuriaga, A.P.A.; Freitas, B.C.B.; Morais, M.G.; Costa, J.A.V. Innovative polyhydroxybutyrate production by Chlorella fusca grown with pentoses. Bioresour. Technol. 2018, 265, 456–463. [Google Scholar] [CrossRef]
- Chaogang, W.; Zhangli, H.; Anping, L.; Baohui, J. Biosynthesis of Poly-3-hydroxybutyrate (PHB) in the transgenic green alga Chlamydomonas reinhardtii. J. Phycol. 2010, 46, 396–402. [Google Scholar] [CrossRef]
- Berlanga, M.; Montero, M.T.; Fernández-Borrell, J.; Guerrero, R. Rapid spectrofluorometric screening of poly-hydroxyalkanoate-producing bacteria from microbial mats. Int. Microbiol. 2006, 9, 95–102. [Google Scholar] [CrossRef]
- Nishioka, M.; Nakai, K.; Miyake, M.; Asada, Y.; Taya, M. Production of poly-β-hydroxybutyrate by thermophilic cyanobacterium, Synechococcus sp. MA19, under phosphate-limited conditions. Biotechnol. Lett. 2001, 23, 1095–1099. [Google Scholar] [CrossRef]
- Albuquerque, M.G.E.; Torres, C.A.V.; Reis, M.A.M. Polyhydroxyalkanoate (PHA) production by a mixed microbial culture using sugar molasses: Effect of the influent substrate concentration on culture selection. Water Res. 2010, 44, 3419–3433. [Google Scholar] [CrossRef] [PubMed]
- Fradinho, J.C.; Domingos, J.M.B.; Carvalho, G.; Oehmen, A.; Reis, M.A.M. Polyhydroxyalkanoates production by a mixed photosynthetic consortium of bacteria and algae. Bioresour. Technol. 2013, 132, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Salgaonkar, B.B.; Bragança, J.M. Utilization of sugarcane bagasse by Halogeometricum borinquense strain e3 for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate). Bioengineering 2017, 4, 50. [Google Scholar] [CrossRef] [Green Version]
- Schlebusch, M.; Forchhammer, K. Requirement of the nitrogen starvation-induced protein s110783 for polyhydroxybutyrate accumulation in Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2010, 76, 6101–6107. [Google Scholar] [CrossRef] [Green Version]
- Sudesh, K.; Taguchi, K.; Doi, Y. Effect of increased PHA synthase activity on polyhydroxyalkanoates biosynthesis in Synechocystis sp. PCC6803. Int. J. Biol. Macromol. 2002, 30, 97–104. [Google Scholar] [CrossRef]
- Vincenzini, M.; Sili, C.; De Philippis, R.; Ena, A.; Materassi, R. Occurrence of poly-β-hydroxybutyrate in Spirulina species. J. Bacteriol. 1990, 172, 2791–2792. [Google Scholar] [CrossRef] [Green Version]
- Taroncher-Oldenburg, G.; Nishina, K.; Stephanopoulos, G. Identification and analysis of the polyhydroxyalkanoate-specific β-ketothiolase and acetoacetyl coenzyme a reductase genes in the cyanobacterium Synechocystis sp. strain PCC6803. Appl. Environ. Microbiol. 2000, 66, 4440–4448. [Google Scholar] [CrossRef] [Green Version]
- Abed, R.M.M.; Garcia-Pichel, F. Long-term compositional changes after transplant in a microbial mat cyanobacterial community revealed using a polyphasic approach. Environ. Microbiol. 2001, 3, 53–62. [Google Scholar] [CrossRef]
- Stubbe, J.A.; Tian, J.; He, A.; Sinskey, A.J.; Lawrence, A.G.; Liu, P. Nontemplate-dependent polymerization processes: Polyhydroxyalkanoate synthases as a paradigm. Annu. Rev. Biochem. 2005, 74, 433–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, B.H.A. Polyester synthases: Natural catalysts for plastics. Biochem. J. 2003, 376, 15–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastav, A.; Mishra, S.K.; Mishra, S. Polyhydroxyalkanoate (PHA) synthesis by Spirulina subsalsa from Gujarat coast of India. Int. J. Biol. Macromol. 2010, 46, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, P.; Rehm, B.H.A.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef]
- Bertozzini, E.; Galluzzi, L.; Penna, A.; Magnani, M. Application of the standard addition method for the absolute quantification of neutral lipids in microalgae using Nile red. J. Microbiol. Methods 2011, 87, 17–23. [Google Scholar] [CrossRef]
- Zuriani, R.; Vigneswari, S.; Azizan, M.N.M.; Majid, M.I.A.; Amirul, A.A. A high throughput Nile red fluorescence method for rapid quantification of intracellular bacterial polyhydroxyalkanoates. Biotechnol. Bioprocess. Eng. 2013, 18, 472–478. [Google Scholar] [CrossRef]
- Elsey, D.; Jameson, D.; Raleigh, B.; Cooney, M.J. Fluorescent measurement of microalgal neutral lipids. J. Microbiol. Methods 2007, 68, 639–642. [Google Scholar] [CrossRef]
- Praveenkumar, R.; Johncy, K.; MubarakAli, D.; Vijayan, D.; Thajuddin, N.; Gunasekaran, M. Demonstration of increased lipid accumulation potential of Stigeoclonium sp., Kütz. BUM11007 under nitrogen starved regime: A new source of lipids for biodiesel production. J. Biobased Mater. Bioenergy 2012, 6, 209–213. [Google Scholar] [CrossRef]
- Vischer, W. Über einige kritische gattungen und die systematik der chaetophorales. Beih. Bot. Centralbl. 1933, 51, 1–101. [Google Scholar]
- Hoshina, R.; Kato, Y.; Kamako, S.; Imamura, N. Genetic evidence of “American” and “European” type symbiotic algae of Poramecium bursaria Ehrenberg. Plant. Biol. 2005, 7, 526–532. [Google Scholar] [CrossRef]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94. [Google Scholar] [CrossRef]
- Allen, M.M. Simple Conditions for growth of unicellular blue-green algae on plates. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Meeks, J.C.; Castenholz, R.W. Growth and photosynthesis in an extreme thermophile, Synechococcus lividus (Cyanophyta). Arch. Mikrobiol. 1971. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.D.F.; Neilan, B.A.; Copp, J.N.; Rodrigues, J.L.M.; Tsai, S.M.; Lee, H.; Trevors, J.T. Characterization of nitrogen-fixing cyanobacteria in the Brazilian Amazon floodplain. Water Res. 2005, 39, 5017–5026. [Google Scholar] [CrossRef] [PubMed]
- Law, J.H.; Slepecky, R.A. Assay of poly-beta-hydroxybutyric acid. J. Bacteriol. 1961, 82, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, J.H.; Slepecky, R.A. A Rapid Spectrophotometric assay of alpha, beta-unsaturated acids and beta-hydroxy acids. Anal. Chem. 1960, 32, 1697–1699. [Google Scholar] [CrossRef]
- Getachew, A.; Woldesenbet, F. Production of biodegradable plastic by polyhydroxybutyrate (PHB) accumulating bacteria using low cost agricultural waste material. BMC Res. Notes 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Vanormelingen, P.; Vyverman, W. Fatty acid profiles of four filamentous green algae under varying culture conditions. Bioresour. Technol. 2016, 200, 1080–1084. [Google Scholar] [CrossRef]
- Marks, J.C.; Lowe, R.L. The independent and interactive effects of snail grazing and nutrient enrichment on structuring periphyton communities. Hydrobiologia 1989, 185, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Luttenton, M.R.; Lowe, R.L. Response of a lentic periphyton community to nutrient enrichment at low N:P ratios. J. Phycol. 2006, 42, 1007–1015. [Google Scholar] [CrossRef]
- Lee, S.Y.; Park, S.J. Biosynthesis and fermentative production of SCL-MCL-PHAs. In Biopolymers Online, 3rd ed.; Hofrichter, M., Steinbüchel, A., Eds.; Wiley-VCH: Weinheim, Germany, 2005; Volume 1, pp. 263–290. [Google Scholar] [CrossRef]
- Brigham, C.J.; Kurosawa, K. Bacterial carbon storage to value added products. J. Microb. Biochem. Technol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Higuchi-Takeuchi, M.; Morisaki, K.; Toyooka, K.; Numata, K. Synthesis of high-molecular-weight polyhydroxyalkanoates by marine photosynthetic purple bacteria. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariffin, H.; Nishida, H.; Shirai, Y.; Hassan, M.A. Determination of multiple thermal degradation mechanisms of poly(3-hydroxybutyrate). Polym. Degrad. Stab. 2008, 93, 1433–1439. [Google Scholar] [CrossRef]
- Rajankar, M.P.; Ravindranathan, S.; Rajamohanan, P.R.; Raghunathan, A. Absolute quantitation of poly(R)-3-hydroxybutyric acid using spectrofluorometry in recombinant Escherichia coli. Biol. Methods Protoc. 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Gorenflo, V.; Steinbüchel, A.; Marose, S.; Rieseberg, M.; Scheper, T. Quantification of bacterial polyhydroxyalkanoic acids by Nile red staining. Appl. Microbiol. Biotechnol. 1999, 51, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Staehelin, L.A. Chloroplast Structure and Supramolecular Organization of Photosynthetic Membranes. In Photosynthesis III, 1st ed.; Staehelin, L.A., Arntzen, C.J., Eds.; Encyclopedia of Plant Physiology (New Series); Springer: Berlin/Heidelberg, Germany, 1986; Volume 19, pp. 1–84. [Google Scholar] [CrossRef]
- Bhuwal, A.K.; Singh, G.; Aggarwal, N.K.; Goyal, V.; Yadav, A. Poly-β-hydroxybutyrate production and management of cardboard industry effluent by new Bacillus sp. NA10. Bioresour. Bioprocess. 2014, 1. [Google Scholar] [CrossRef] [Green Version]
- Kamravamanesh, D.; Pflügl, S.; Nischkauer, W.; Limbeck, A.; Lackner, M.; Herwig, C. Photosynthetic poly-β-hydroxybutyrate accumulation in unicellular cyanobacterium Synechocystis sp. PCC 6714. AMB Express 2017, 7. [Google Scholar] [CrossRef]
- Asada, Y.; Miyake, M.; Miyake, J.; Kurane, R.; Tokiwa, Y. Photosynthetic accumulation of poly-(hydroxybutyrate) by cyanobacteria—The metabolism and potential for CO2 recycling. Int. J. Biol. Macromol. 1999, 25, 37–42. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, T.; Cai, Z.; Li, Y. From cyanochemicals to cyanofactories: A review and perspective. Microb. Cell Fact. 2016, 15. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Carbon Source 1 | Carbon Source 2 | Nitrogen Source |
|---|---|---|---|
| BG-11 | * | * | Sodium nitrate |
| BG-11A | Sodium acetate | * | Sodium nitrate |
| BG-11B | * | Sodium bicarbonate | Sodium nitrate |
| BG-11A+B | Sodium acetate | Sodium bicarbonate | Sodium nitrate |
| BG-110 | * | * | * |
| BG-110+A | Sodium acetate | * | * |
| BG-110+B | * | Sodium bicarbonate | * |
| BG-110+A+B | Sodium acetate | Sodium bicarbonate | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourão, M.M.; Gradíssimo, D.G.; Santos, A.V.; Schneider, M.P.C.; Faustino, S.M.M.; Vasconcelos, V.; Xavier, L.P. Optimization of Polyhydroxybutyrate Production by Amazonian Microalga Stigeoclonium sp. B23. Biomolecules 2020, 10, 1628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10121628
Mourão MM, Gradíssimo DG, Santos AV, Schneider MPC, Faustino SMM, Vasconcelos V, Xavier LP. Optimization of Polyhydroxybutyrate Production by Amazonian Microalga Stigeoclonium sp. B23. Biomolecules. 2020; 10(12):1628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10121628
Chicago/Turabian StyleMourão, Murilo Moraes, Diana Gomes Gradíssimo, Agenor Valadares Santos, Maria Paula Cruz Schneider, Silvia Maria Mathes Faustino, Vitor Vasconcelos, and Luciana Pereira Xavier. 2020. "Optimization of Polyhydroxybutyrate Production by Amazonian Microalga Stigeoclonium sp. B23" Biomolecules 10, no. 12: 1628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10121628