A Novel Serine Protease Inhibitor PE-BBI Ameliorates Cockroach Extract-Mediated Airway Epithelial Barrier Dysfunction

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract and Reagents
2.2. Peptide Inhibitors
2.3. Determination of Putative Protease Inhibitor Potency versus Recombinant Trypsin and CRE TLP Activity
2.4. Cell Culture
2.5. The 16HBE Cell Surface TLP Activity Assay
2.6. Cell Viability Assay
2.7. Airway Epithelial Cell Barrier Measurement
2.8. Statistical Analysis
3. Results
3.1. Evaluation of Putative Protease Inhibitors versus Trypsin Activities
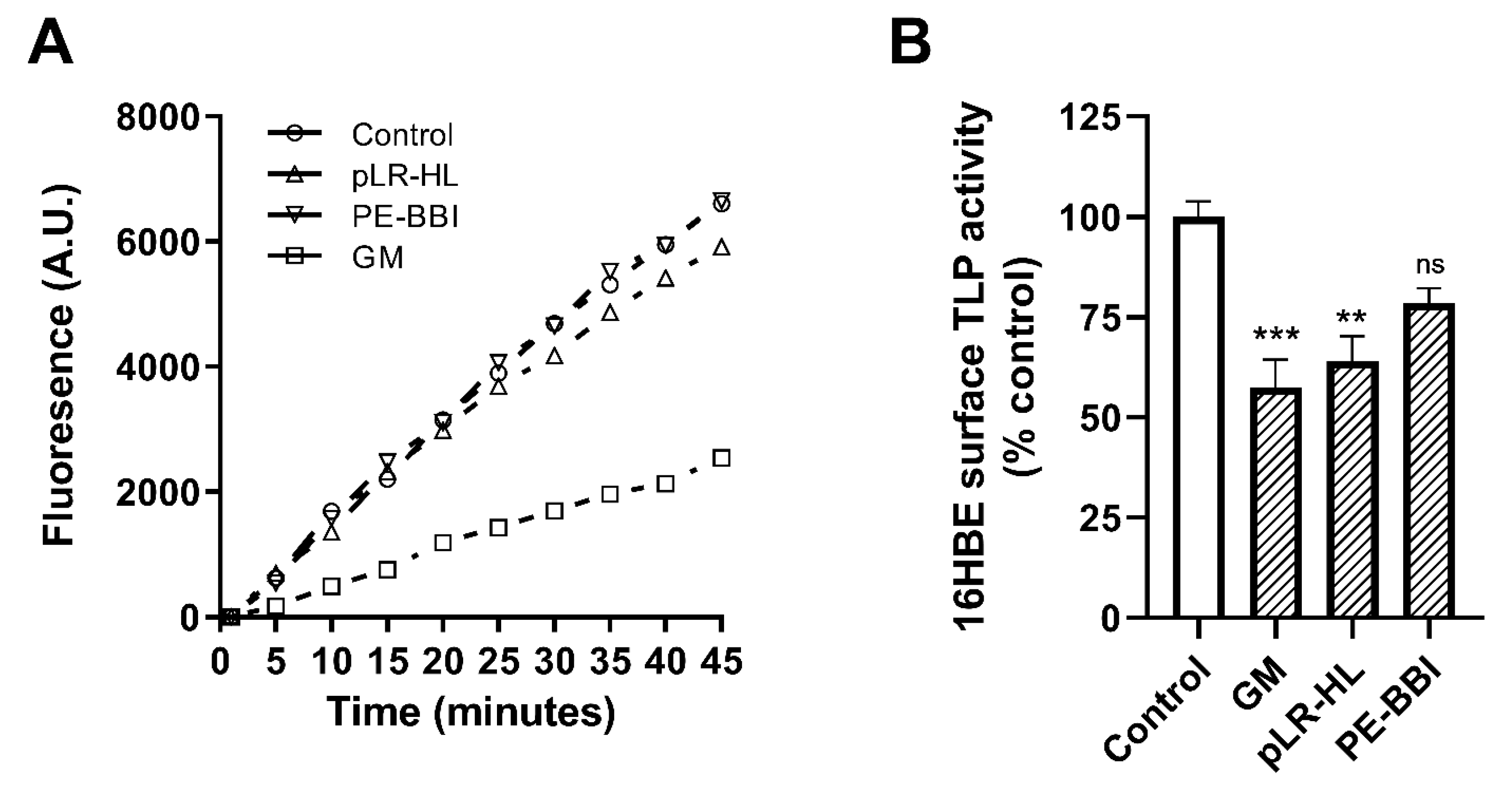
3.2. Evaluation of Putative Protease Inhibitors versus 16HBE Cell Surface Activity
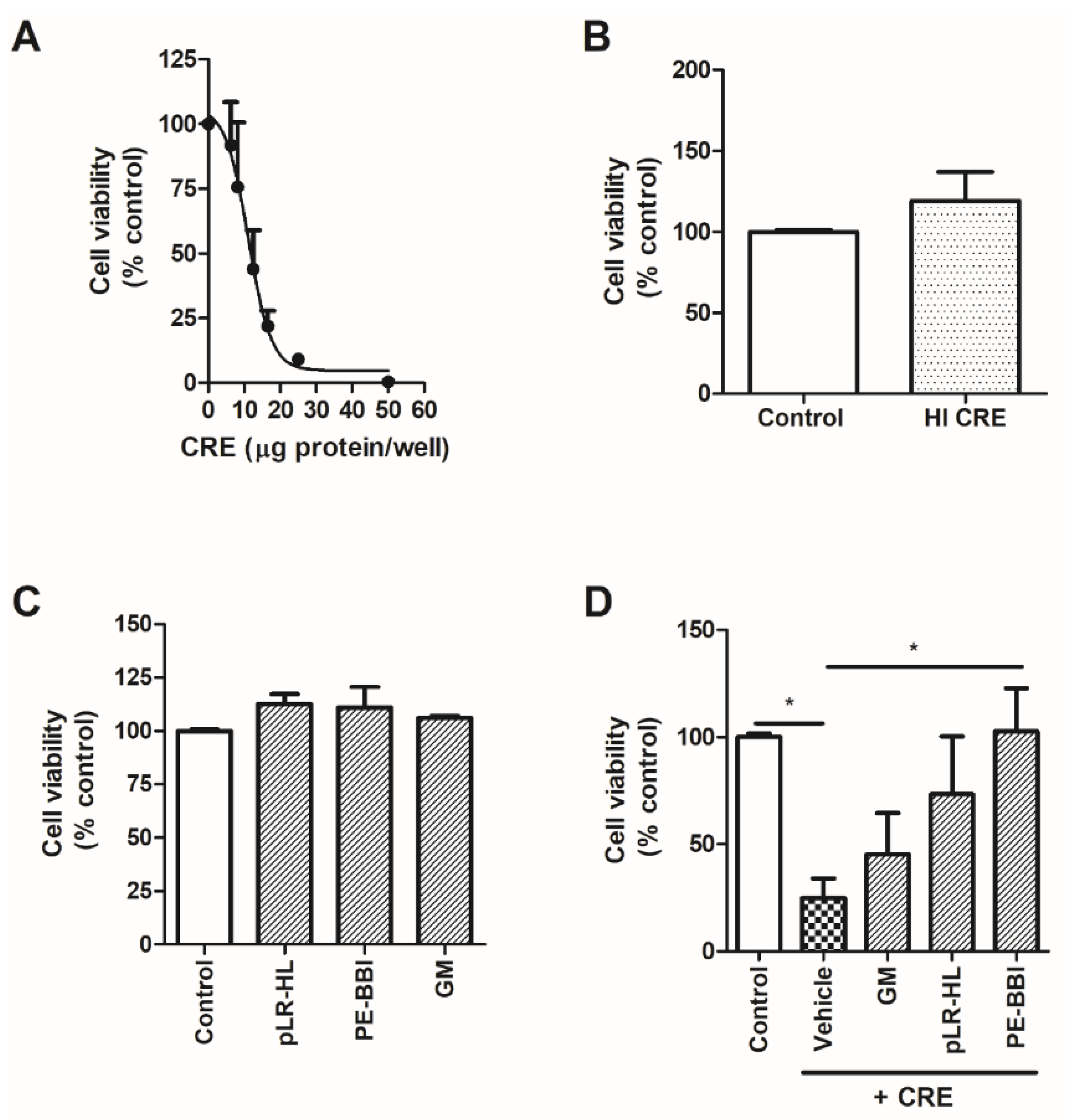
3.3. PE-BBI Protects 16HBE Cells from CRE-Induced Damage
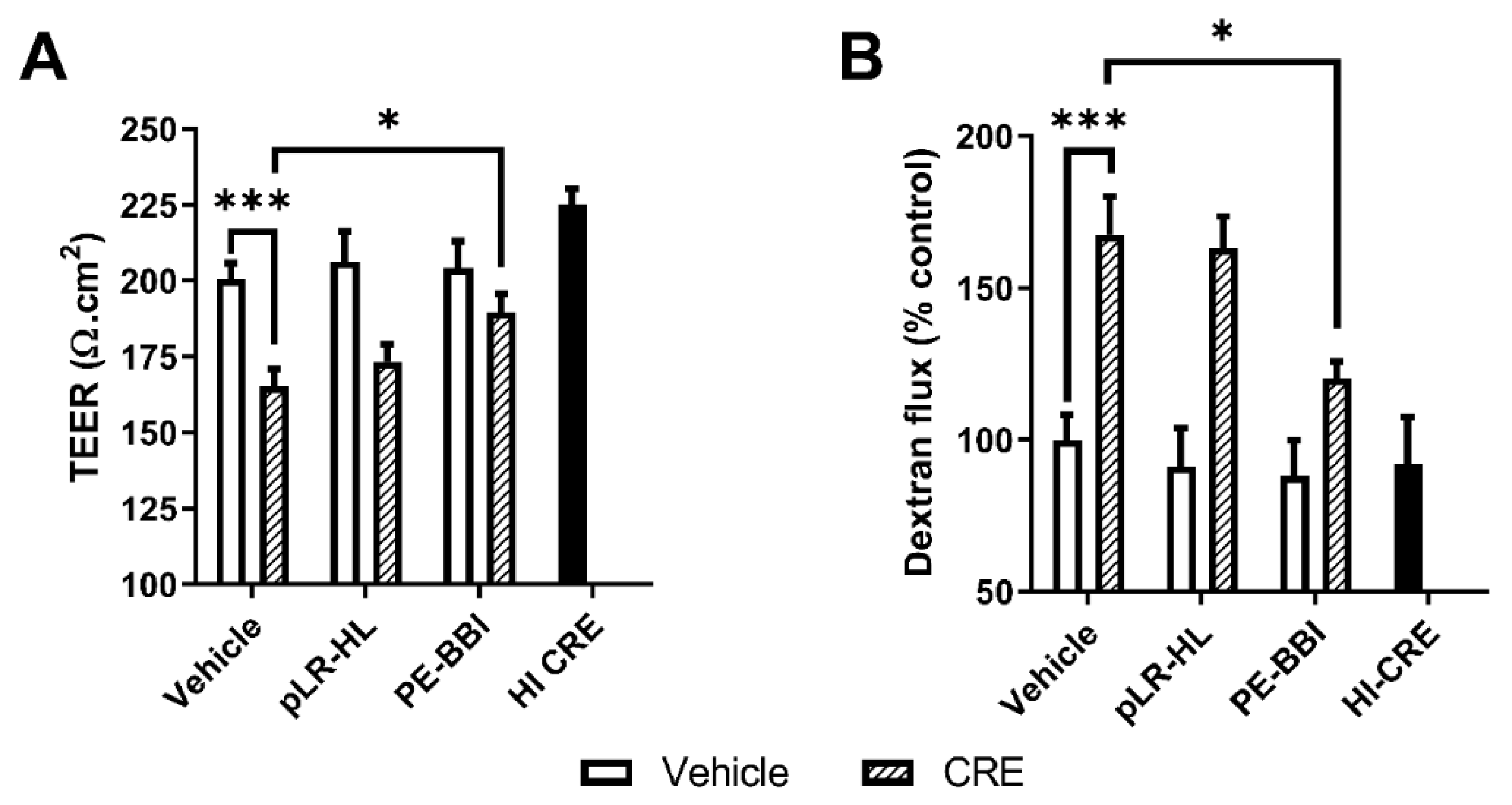
3.4. PE-BBI Prevented CRE-Mediated Epithelial Barrier Disruption
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- The Global Asthma Report 2018; Global Asthma Network: Auckland, New Zealand, 2018.
- Lopez-Rodriguez, J.C.; Benede, S.; Barderas, R.; Villalba, M.; Batanero, E. Airway Epithelium Plays a Leading Role in the Complex Framework Underlying Respiratory Allergy. J. Investig. Allergol. Clin. Immunol. 2017, 27, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T. The sentinel role of the airway epithelium in asthma pathogenesis. Immunol. Rev. 2011, 242, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Do, D.C.; Zhao, Y.; Gao, P. Cockroach allergen exposure and risk of asthma. Allergy 2016, 71, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Sudha, V.T.; Arora, N.; Singh, B.P. Serine protease activity of Per a 10 augments allergen-induced airway inflammation in a mouse model. Eur. J. Clin. Investig. 2009, 39, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.R.; Hales, B.J.; Smith, W.-A. House dust mite allergens in asthma and allergy. Trends Mol. Med. 2010, 16, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.W.; Venaille, T.J.; Mendis, A.H.; McAleer, R. Allergens as proteases: An Aspergillus fumigatus proteinase directly induces human epithelial cell detachment. J. Allergy Clin. Immunol. 1990, 86, 726–731. [Google Scholar] [CrossRef]
- Kale, S.L.; Agrawal, K.; Gaur, S.N.; Arora, N. Cockroach protease allergen induces allergic airway inflammation via epithelial cell activation. Sci. Rep. 2017, 7, 42341. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.E.; Jee, H.M.; Hong, J.Y.; Kim, M.N.; Oh, M.S.; Kim, Y.S.; Kim, K.W.; Kim, K.E.; Sohn, M.H. German Cockroach Extract Induces Matrix Metalloproteinase-1 Expression, Leading to Tight Junction Disruption in Human Airway Epithelial Cells. Yonsei Med. J. 2018, 59, 1222–1231. [Google Scholar] [CrossRef]
- Page, K.; Lierl, K.M.; Herman, N.; Wills-Karp, M. Differences in susceptibility to German cockroach frass and its associated proteases in induced allergic inflammation in mice. Respir. Res. 2007, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.K.; Page, K.; Tan, A.; Hershenson, M.B. German cockroach extract increases bronchial epithelial cell interleukin-8 expression. Clin. Exp. Allergy 2003, 33, 35–42. [Google Scholar] [CrossRef]
- Page, K.; Strunk, V.S.; Hershenson, M.B. Cockroach proteases increase IL-8 expression in human bronchial epithelial cells via activation of protease-activated receptor (PAR)-2 and extracellular-signal-regulated kinase. J. Allergy Clin. Immunol. 2003, 112, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.F.; Chang, C.W.; Wang, N.M.; Lin, S.J.; Chen, Y.H. Serine protease inhibitor gabexate mesilate attenuates american cockroach-induced bronchial damage and inflammatory cytokine release. J. Investig. Allergol. Clin. Immunol. 2014, 24, 338–345. [Google Scholar] [PubMed]
- Lyu, P.; Ge, L.; Ma, R.; Wei, R.; McCrudden, C.M.; Chen, T.; Shaw, C.; Kwok, H.F. Identification and pharmaceutical evaluation of novel frog skin-derived serine proteinase inhibitor peptide-PE-BBI (Pelophylax esculentus Bowman-Birk inhibitor) for the potential treatment of cancer. Sci. Rep. 2018, 8, 14502. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Hang, H.; Chen, T.; Zhou, M.; Wang, L.; Shaw, C. pLR-HL: A Novel Amphibian Bowman-Birk-type Trypsin Inhibitor from the Skin Secretion of the Broad-folded Frog, Hylarana latouchii. Chem. Biol. Drug Des. 2016, 87, 91–100. [Google Scholar] [CrossRef]
- Laskowski, M., Jr.; Kato, I. Protein inhibitors of proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Brauer, A.B.; Kelly, G.; Matthews, S.J.; Leatherbarrow, R.J. The (1)H-NMR solution structure of the antitryptic core peptide of Bowman-Birk inhibitor proteins: A minimal canonical loop. J. Biomol. Struct. Dyn. 2002, 20, 59–70. [Google Scholar] [CrossRef]
- Qi, R.F.; Song, Z.W.; Chi, C.W. Structural features and molecular evolution of Bowman-Birk protease inhibitors and their potential application. Acta Biochim. Biophys. Sin. 2005, 37, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.R. Chemopreventive agents: Protease inhibitors. Pharmacol. Ther. 1998, 78, 167–209. [Google Scholar] [CrossRef]
- Malkowicz, S.B.; McKenna, W.G.; Vaughn, D.J.; Wan, X.S.; Propert, K.J.; Rockwell, K.; Marks, S.H.; Wein, A.J.; Kennedy, A.R. Effects of Bowman-Birk inhibitor concentrate (BBIC) in patients with benign prostatic hyperplasia. Prostate 2001, 48, 16–28. [Google Scholar] [CrossRef]
- Armstrong, W.B.; Kennedy, A.R.; Wan, X.S.; Taylor, T.H.; Nguyen, Q.A.; Jensen, J.; Thompson, W.; Lagerberg, W.; Meyskens, F.L., Jr. Clinical modulation of oral leukoplakia and protease activity by Bowman-Birk inhibitor concentrate in a phase IIa chemoprevention trial. Clin. Cancer Res. 2000, 6, 4684–4691. [Google Scholar]
- Cozens, A.; Yezzi, M.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.; Widdicombe, J.; Gruenert, D. CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1994, 10, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Swindle, E.J.; Collins, J.E.; Davies, D.E. Breakdown in epithelial barrier function in patients with asthma: Identification of novel therapeutic approaches. J. Allergy Clin. Immunol. 2009, 124, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ilowite, J.S.; Bennett, W.D.; Sheetz, M.S.; Groth, M.L.; Nierman, D.M. Permeability of the bronchial mucosa to 99mTc-DTPA in asthma. Am. Rev. Respir. Dis. 1989, 139, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, K.; Kale, S.L.; Arora, N. Protease activity of Per a 10 potentiates Th2 polarization by increasing IL-23 and OX40L. Eur. J. Immunol. 2015, 45, 3375–3385. [Google Scholar] [CrossRef] [PubMed]
- Kale, S.L.; Arora, N. Per a 10 activates human derived epithelial cell line in a protease dependent manner via PAR-2. Immunobiology 2015, 220, 525–532. [Google Scholar] [CrossRef]
- Day, S.B.; Ledford, J.R.; Zhou, P.; Lewkowich, I.P.; Page, K. German cockroach proteases and protease-activated receptor-2 regulate chemokine production and dendritic cell recruitment. J. Innate. Immun. 2012, 4, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.F.; Wang, N.M.; Liu, S.W.; Lin, S.J.; Chen, Y.H. Induction of interleukin 8 by American cockroach allergens from human airway epithelial cells via extracellular signal regulatory kinase and jun N-terminal kinase but not p38 mitogen-activated protein kinase. Ann. Allergy Asthma Immunol. 2010, 105, 234–240. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; Xu, X.; Wang, J.; Yu, H.; Lai, R.; Gong, W. Trypsin inhibitory loop is an excellent lead structure to design serine protease inhibitors and antimicrobial peptides. FASEB J. 2007, 21, 2466–2473. [Google Scholar] [CrossRef] [Green Version]
- Vliagoftis, H.; Forsythe, P. Should we target allergen protease activity to decrease the burden of allergic airway inflammation? Inflamm. Allergy Drug Targets 2008, 7, 288–295. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reihill, J.A.; Ouyang, X.; Yang, Z.; Douglas, L.E.J.; Zhou, M.; Chen, T.; Martin, S.L. A Novel Serine Protease Inhibitor PE-BBI Ameliorates Cockroach Extract-Mediated Airway Epithelial Barrier Dysfunction. Biomolecules 2020, 10, 515. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040515
Reihill JA, Ouyang X, Yang Z, Douglas LEJ, Zhou M, Chen T, Martin SL. A Novel Serine Protease Inhibitor PE-BBI Ameliorates Cockroach Extract-Mediated Airway Epithelial Barrier Dysfunction. Biomolecules. 2020; 10(4):515. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040515
Chicago/Turabian StyleReihill, James A., Xuan Ouyang, Zhixuan Yang, Lisa E. J. Douglas, Mei Zhou, Tianbao Chen, and S. Lorraine Martin. 2020. "A Novel Serine Protease Inhibitor PE-BBI Ameliorates Cockroach Extract-Mediated Airway Epithelial Barrier Dysfunction" Biomolecules 10, no. 4: 515. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040515