Temporal Dynamics of High-Density Lipoprotein Proteome in Diet-Controlled Subjects with Type 2 Diabetes

 ,
,
Abstract
:1. Background
2. Methods
2.1. Study Subjects
2.2. Kinetic Study
2.3. Analytical Procedures
2.3.1. Body Water 2H2O Enrichment Measurement
2.3.2. High-Density Lipoprotein (HDL) Isolation and Proteome Composition
2.3.3. HDL Dynamics Analysis
2.4. Data Presentation and Statistical Analysis
3. Results
3.1. Subject Characteristics
3.2. HDL Cholesterol Turnover
3.3. HDL Proteome Composition and Dynamics
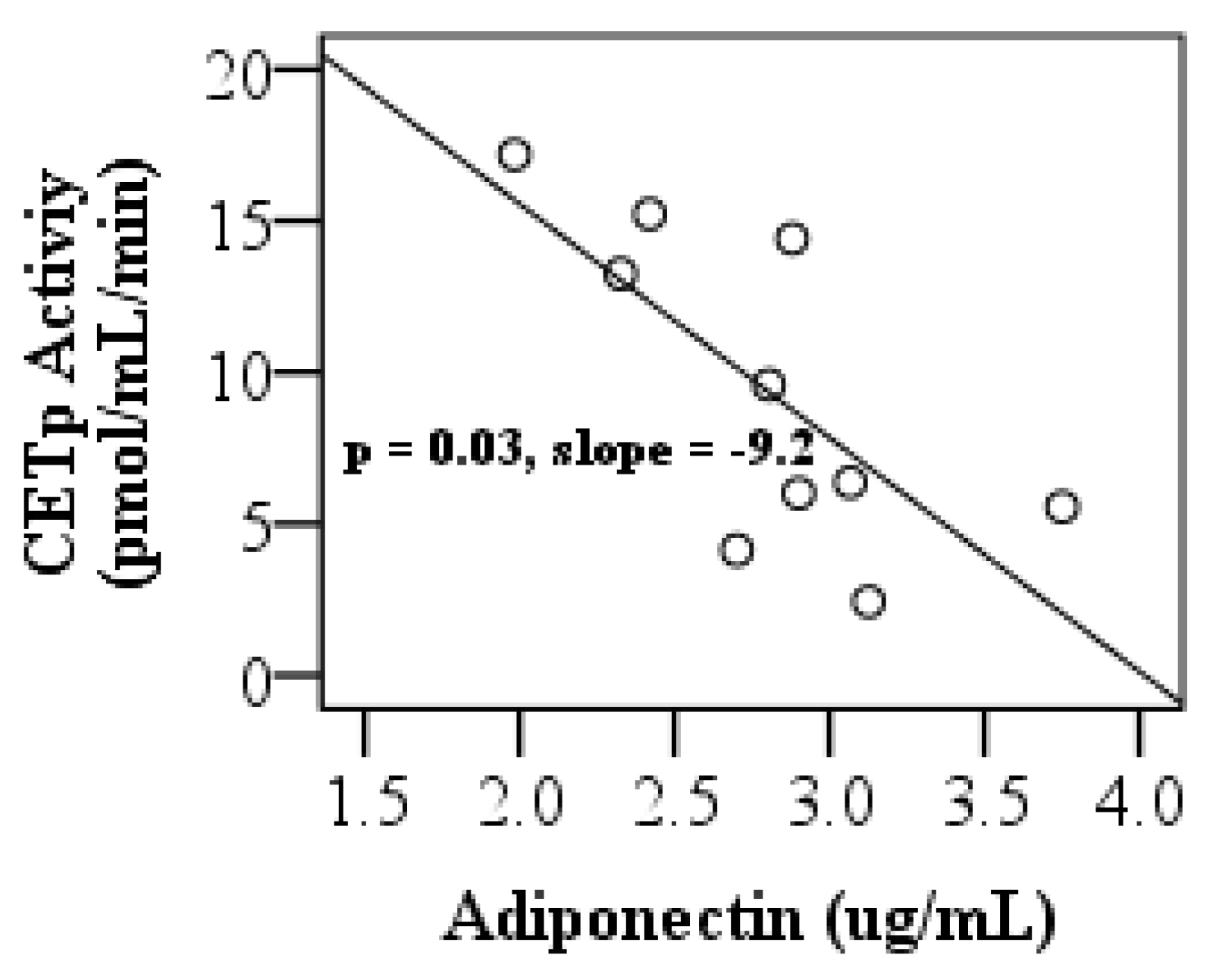
3.4. Adiponectin Levels, LCAT and, CETP Activities, and Pro-inflammatory Index of HDL
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Cardiovascular disease | CVD |
| Type 1 diabetes | T1D |
| High-density lipoprotein | HDL |
| High- sensitivity C-reactive protein | hsCRP |
| Reverse cholesterol transport | RCT |
| Apolipoprotein A-1 | ApoA-1 |
| Type 2 diabetes | T2D |
| Cholesterol ester transfer protein | CETP |
| Lecithin–cholesterol acyltransferase | LCAT |
| Liquid chromatography-tandem mass spectrometry | LC-MS/MS |
| Myeloperoxidase | MPO |
| Gas chromatography-mass spectrometry | GC-MS |
| Complement C3 | C3 |
| Production rate | PR |
| Hemoglobin A1c | HbA1c |
| Fractional catabolic rate | FCR |
| Confidence interval | CI |
| Posttranslational modification | PTM |
| Paraoxonase 1 | PON1 |
References
- Perreault, L.; Boardman, M.; Pak, J. The association between type 2 diabetes and cardiovascular disease: The “for your sweetheart™” survey. Adv. Ther. 2019, 36, 746–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, W.; Probstfield, J.; Anderson, T.; Chaitman, B.; Desvignes-Nickens, P.; Koprowicz, K.; McBride, R.; Teo, K.; Weintraub, W. Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N. Engl. J. Med. 2011, 365, 2255–2267. [Google Scholar]
- Tall, A.; Yvan-Charvet, L.; Wang, N. The failure of torcetrapib: Was it the molecule or the mechanism? Arterioscler. Thromb. Vasc. Biol. 2007, 27, 257–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoni, P.; Khetarpal, S.; Larach, D.; Hancock-Cerutti, W.; Millar, J.; Cuchel, M. Rare variant in scavenger receptor BI raises HDL cholesterol and increases risk of coronary heart disease. Science 2016, 351, 1166–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleheen, D.; Scott, R.; Javad, S.; Zhao, W.; Rodrigues, A.; Picataggi, A.; Lukmanova, D.; Mucksavage, M.L.; Luben, R.; Billheimer, J.; et al. Association of HDL cholesterol efflux capacity with incident coronary heart disease events: A prospective case-control study. Lancet Diabetes Endocrinol. 2015, 3, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Farbstein, D.; Levy, A. HDL dysfunction in diabetes: Causes and possible treatments. Expert Rev. Cardiovasc. Ther. 2012, 10, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, E.; McNamara, J.; Shah, P.; Nakajima, K.; Cupples, L.; Ordovas, J.; Wilson, P. Elevated remnant-like particle cholesterol and triglyceride levels in diabetic men and women in the Framingham Offspring Study. Diabetes Care 2002, 25, 989–994. [Google Scholar] [CrossRef] [Green Version]
- Ronsein, G.; Reyes-Soffer, G.; He, Y.; Oda, M.; Ginsberg, H.; Heinecke, J. Targeted proteomics identifies paraoxonase/arylesterase 1 (PON1) and apolipoprotein Cs as potential risk factors for hypoalphalipoproteinemia in diabetic subjects treated with fenofibrate and rosiglitazone. Mol. Cell. Proteom. 2016, 15, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Manjunatha, S.; Distelmaier, K.; Dasari, S.; Carter, R.; Kudva, Y.; Nair, K. Functional and proteomic alterations of plasma high density lipoproteins in type 1 diabetes mellitus. Metabolism 2016, 65, 1421–1431. [Google Scholar] [CrossRef]
- Gordon, S.; Davidson, W.; Urbina, E.; Dolan, L.; Heink, A.; Zang, H.; Lu, L.; Shah, A. The effects of type 2 diabetes on lipoprotein composition and arterial stiffness in male youth. Diabetes 2013, 62, 2958–2967. [Google Scholar] [CrossRef] [Green Version]
- Cubedo, J.; Padro, T.; Garcia-Arguinzonis, M.; Vilahur, G.; Minambres, I.; Pou, J.M.; Ybarra, J.; Badimon, L. A novel truncated form of apolipoprotein A-I transported by dense LDL is increased in diabetic patients. J. Lipid Res. 2015, 56, 1762–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, E.; Feig, J.; Hewing, B.; Hazen, S.; Smith, J. High-density lipoprotein function, dysfunction, and reverse cholesterol transport. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2813–2820. [Google Scholar] [CrossRef] [Green Version]
- Rosenson, R.; Brewer, H.; Davidson, W.; Fayad, Z.; Fuster, V.; Goldstein, J.; Hellerstein, M.; Jiang, X.; Phillips, M.; Rader, D.; et al. Cholesterol efflux and atheroprotection: Advancing the concept of reverse cholesterol transport. Circulation 2012, 125, 1905–1919. [Google Scholar] [CrossRef] [Green Version]
- Toth, P.; Barter, P.; Rosenson, R.; Boden, W.; Chapman, M.; Cuchel, M.; D’Agostino, R.; Davidson, M.; Davidson, W.; Heinecke, J.; et al. High-density lipoproteins: A consensus statement from the National Lipid Association. J. Clin. Lipidol. 2013, 7, 484–525. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, S.; Besler, C.; Rohrer, L.; Meyer, M.; Heinrich, K.; Bahlmann, F.; Mueller, M.; Horvath, T.; Doerries, C.; Heinemann, M.; et al. Endothelial-vasoprotective effects of high-density lipoprotein are impaired in patients with type 2 diabetes mellitus but are improved after extended-release niacin therapy. Circulation 2010, 121, 110–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaleel, A.; Henderson, G.; Madden, B.; Klaus, K.; Morse, D.; Gopala, S.; Nair, K. Identification of de novo synthesized and relatively older proteins accelerated oxidative damage to de novo synthesized apolipoprotein A-1 in type 1 diabetes. Diabetes 2010, 59, 2366–2374. [Google Scholar] [CrossRef] [Green Version]
- Shao, B.; Heinecke, J. Quantifying HDL proteins by mass spectrometry: How many proteins are there and what are their functions? Expert Rev. Proteom. 2018, 15, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Swertfeger, D.; Li, H.; Rebholz, S.; Zhu, X.; Shah, A.; Davidson, W.; Lu, L. Mapping atheroprotective functions and related proteins/lipoproteins in size fractionated human plasma. Mol. Cell. Proteom. 2017, 16, 680–693. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.; Ng, T.; Watts, G. Apolipoprotein A-II: Evaluating its significance in dyslipidaemia, insulin resistance, and atherosclerosis. Ann. Med. 2012, 44, 313–324. [Google Scholar] [CrossRef]
- Fujimoto, K.; Fukagawa, K.; Sakata, T.; Tso, P. Suppression of food intake by apolipoprotein A-IV is mediated through the central nervous system in rats. J. Clin. Invest. 1993, 91, 1830–1833. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, M.; Wang, F.; Kohan, A.; Haas, M.; Yang, Q.; Lou, D.; Obici, S.; Davidson, W.; Tso, P. Apolipoprotein A-IV reduces hepatic gluconeogenesis through nuclear receptor NR1D1. J. Biol. Chem. 2014, 289, 2396–2404. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Kohan, A.; Kindel, T.; Corbin, K.; Nunemaker, C.; Obici, S.; Woods, S.; Davidson, W.; Tso, P. Apolipoprotein A-IV improves glucose homeostasis by enhancing insulin secretion. Proc. Natl. Acad. Sci. USA 2012, 109, 9641–9646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, N.; Atger, V.; Paul, J.; Sturm, M.; Duverger, N.; Rothblat, G.; Moatti, N. Human ApoA-IV overexpression in transgenic mice induces cAMP-stimulated cholesterol efflux from J774 macrophages to whole serum. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1283–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duverger, N.; Tremp, G.; Caillaud, J.; Emmanuel, F.; Castro, G.; Fruchart, J.; Steinmetz, A.; Denefle, P. Protection against atherogenesis in mice mediated by human apolipoprotein A-IV. Science 1996, 273, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.; Shih, D.; Hama, S.; Villa, N.; Navab, M.; Reddy, S. The paraoxonase gene family and atherosclerosis. Free Radic. Biol. Med. 2005, 38, 153–163. [Google Scholar] [CrossRef]
- Sviridov, D.; Nestel, P. Dynamics of reverse cholesterol transport: Protection against atherosclerosis. Atherosclerosis 2002, 161, 245–254. [Google Scholar] [CrossRef]
- Bach-Ngohou, K.; Ouguerram, K.; Frenais, R.; Maugere, P.; Ripolles-Piquer, B.; Zair, Y.; Krempf, M.; Bard, J. Influence of atorvastatin on apolipoprotein E and AI kinetics in patients with type 2 diabetes. J. Pharmacol. Exp. Ther. 2005, 315, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, S.; Osme, A.; Ilchenko, S.; Golizeh, M.; Lee, K.; Wang, S.; Bena, J.; Previs, S.; Smith, J.; Kasumov, T. Glycation reduces the stability of ApoAI and increases HDL dysfunction in diet-controlled type 2 diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.; Chan, D.; Hamilton, S.; Tenneti, V.; Watts, G.; Barrett, P. Effect of niacin on high-density lipoprotein apolipoprotein A-I kinetics in statin-treated patients with type 2 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chetiveaux, M.; Lalanne, F.; Lambert, G.; Zair, Y.; Ouguerram, K.; Krempf, M. Kinetics of prebeta1 HDL and alphaHDL in type II diabetic patients. Eur. J. Clin. Invest. 2006, 36, 29–34. [Google Scholar] [CrossRef]
- Kasumov, T.; Willard, B.; Li, L.; Li, M.; Conger, H.; Buffa, J.; Previs, S.; McCullough, A.; Hazen, S.; Smith, J. 2H2O-based high-density lipoprotein turnover method for the assessment of dynamic high-density lipoprotein function in mice. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1994–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Bebek, G.; Previs, S.; Smith, J.; Sadygov, R.; McCullough, A.; Willard, B.; Kasumov, T. Proteome dynamics reveals pro-inflammatory remodeling of plasma proteome in a mouse model of NAFLD. J. Proteome Res. 2016, 15, 3388–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golizeh, M.; Lee, K.; Ilchenko, S.; Osme, A.; Bena, J.; Sadygov, R.; Kashyap, S.; Kasumov, T. Increased serotransferrin and ceruloplasmin turnover in diet-controlled patients with type 2 diabetes. Free Radic. Biol. Med. 2017, 113, 461–469. [Google Scholar] [CrossRef]
- Ji, X.; Xu, H.; Zhang, H.; Hillery, C.; Gao, H.; Pritchard, K. Anion exchange HPLC isolation of high-density lipoprotein (HDL) and on-line estimation of proinflammatory HDL. PloS ONE 2014, 9, e91089. [Google Scholar] [CrossRef] [Green Version]
- Navab, M.; Hama, S.; Hough, G.; Subbanagounder, G.; Reddy, S.; Fogelman, A. A cell-free assay for detecting HDL that is dysfunctional in preventing the formation of or inactivating oxidized phospholipids. J. Lipid Res. 2001, 42, 1308–1317. [Google Scholar]
- Shah, V.; Herath, K.; Previs, S.; Hubbard, B.; Roddy, T. Headspace analyses of acetone: A rapid method for measuring the 2H-labeling of water. Anal. Biochem. 2010, 404, 235–237. [Google Scholar] [CrossRef]
- Li, L.; Willard, B.; Rachdaoui, N.; Kirwan, J.; Sadygov, R.; Stanley, W.; Previs, S.; McCullough, A.; Kasumov, T. Plasma proteome dynamics: Analysis of lipoproteins and acute phase response proteins with 2H2O metabolic labeling. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Sadygov, R.; Avva, J.; Rahman, M.; Lee, K.; Ilchenko, S.; Kasumov, T.; Borzou, A. d2ome, software for in vivo protein turnover analysis using heavy water labeling and LC-MS, reveals alterations of hepatic proteome dynamics in a mouse model of NAFLD. J. Proteome Res. 2018, 17, 3740–3748. [Google Scholar] [CrossRef]
- Kasumov, T.; Dabkowski, E.; Shekar, K.; Li, L.; Ribeiro, R.; Walsh, K.; Previs, S.; Sadygov, R.; Willard, B.; Stanley, W. Assessment of cardiac proteome dynamics with heavy water: Slower protein synthesis rates in interfibrillar than subsarcolemmal mitochondria. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1201–H1214. [Google Scholar] [CrossRef] [Green Version]
- Vizcaino, J.; Deutsch, E.; Wang, R.; Csordas, A.; Reisinger, F.; Rios, D.; Dianes, J.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]
- Vizcaino, J.; Csordas, A.; del-Toro, N.; Dianes, J.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef] [PubMed]
- Kasumov, T.; Ilchenko, S.; Li, L.; Rachdaoui, N.; Sadygov, R.; Willard, B.; McCullough, A.; Previs, S. Measuring protein synthesis using metabolic H-2 labeling, high-resolution mass spectrometry, and an algorithm. Anal. Biochem. 2011, 412, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, R. Introduction to Robust Estimation and Hypothesis Testing, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Wilcox, R.; Keselman, H. Modern robust data analysis methods: Measures of central tendency. Psychol. Methods 2003, 8, 254–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuen, K. The two sample trimmed t for unequal population variances. Biometrika 1974, 61, 165–170. [Google Scholar] [CrossRef]
- Keselman, H.; Othman, A.; Wilcox, R.; Fradette, K. The new and improved two-sample T test. Psychol. Sci. 2004, 15, 47–51. [Google Scholar] [CrossRef]
- Wilcox, R. A heteroscedastic method for comparing regression lines at specified design points when using a robus regression estimator. J. Data Sci. 2013, 11, 281–291. [Google Scholar]
- Stahlman, M.; Davidsson, P.; Kanmert, I.; Rosengren, B.; Boren, J.; Fagerberg, B.; Camejo, G. Proteomics and lipids of lipoproteins isolated at low salt concentrations in D2O/sucrose or in KBr. J. Lipid Res. 2008, 49, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Tan, L.; Long, J.; Davidson, W. Proteomic diversity of high density lipoproteins: Our emerging understanding of its importance in lipid transport and beyond. J. Lipid Res. 2013, 54, 2575–2585. [Google Scholar] [CrossRef] [Green Version]
- Vaisar, T.; Pennathur, S.; Green, P.; Gharib, S.; Hoofnagle, A.; Cheung, M.; Byun, J.; Vuletic, S.; Kassim, S.; Singh, P.; et al. Shotgun proteomics implicates protease inhibition and complement activation in the antiinflammatory properties of HDL. J. Clin. Invest. 2007, 117, 746–756. [Google Scholar] [CrossRef]
- Vergès, B.; Petit, J.; Duvillard, L.; Dautin, G.; Florentin, E.; Galland, F.; Gambert, P. Adiponectin is an important determinant of apoA-I catabolism. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1364–1369. [Google Scholar] [CrossRef] [Green Version]
- Yassine, H.; Borges, C.; Schaab, M.; Billheimer, D.; Stump, C.; Reaven, P.; Lau, S.; Nelson, R. Mass spectrometric immunoassay and MRM as targeted MS-based quantitative approaches in biomarker development: Potential applications to cardiovascular disease and diabetes. Proteom. Clin. Appl. 2013, 7, 528–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheniser, K.; Kashyap, S.; Kasumov, T. A systematic review: The appraisal of the effects of metformin on lipoprotein modification and function. Obes. Sci. Pract. 2018, 5, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R. The role of vitamin D binding protein, total and free 25-hydroxyvitamin D in diabetes. Front. Endocrinol. (Lausanne) 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.; Zhao, J.; Sims, P. Interaction between apolipoproteins A-I and A-II and the membrane attack complex of complement. Affinity of the apoproteins for polymeric C9. J. Biol. Chem. 1993, 268, 3632–3638. [Google Scholar]
- Hoofnagle, A.; Wu, M.; Gosmanova, A.; Becker, J.; Wijsman, E.; Brunzell, J.; Kahn, S.; Knopp, R.; Lyons, T.; Heinecke, J. Low clusterin levels in high-density lipoprotein associate with insulin resistance, obesity, and dyslipoproteinemia. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2528–2534. [Google Scholar] [CrossRef] [Green Version]
- Buono, C.; Come, C.; Witztum, J.; Maguire, G.; Connelly, P.; Carroll, M.; Lichtman, A. Influence of C3 deficiency on atherosclerosis. Circulation 2002, 105, 3025–3031. [Google Scholar] [CrossRef] [Green Version]
- Hovland, A.; Jonasson, L.; Garred, P.; Yndestad, A.; Aukrust, P.; Lappegard, K.; Espevik, T.; Mollnes, T. The complement system and toll-like receptors as integrated players in the pathophysiology of atherosclerosis. Atherosclerosis 2015, 241, 480–494. [Google Scholar] [CrossRef] [Green Version]
- Gourgari, E.; Ma, J.; Playford, M.; Mehta, N.; Goldman, R.; Remaley, A.; Gordon, S. Proteomic alterations of HDL in youth with type 1 diabetes and their associations with glycemic control: A case-control study. Cardiovasc. Diabetol. 2019, 18, 43. [Google Scholar] [CrossRef]
- Holzer, M.; Wolf, P.; Curcic, S.; Birner-Gruenberger, R.; Weger, W.; Inzinger, M.; El-Gamal, D.; Wadsack, C.; Heinemann, A.; Marsche, G. Psoriasis alters HDL composition and cholesterol efflux capacity. J. Lipid Res. 2012, 53, 1618–1624. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, L.; Yamada-Fowler, N.; Smith, J.; Thornalley, P.; Rabbani, N. Arginine-directed glycation and decreased HDL plasma concentration and functionality. Nutr. Diabetes 2014, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.; Mahroof, F.; Cook, N.; van Reyk, D.; Davies, M. Hydrazine compounds inhibit glycation of low-density lipoproteins and prevent the in vitro formation of model foam cells from glycolaldehyde-modified low-density lipoproteins. Diabetologia 2006, 49, 775–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krikken, J.; Gansevoort, R.; Dullaart, R. Lower HDL-C and apolipoprotein A-I are related to higher glomerular filtration rate in subjects without kidney disease. J. Lipid Res. 2010, 51, 1982–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubakio-Yamamoto, K.; Matsuura, F.; Koseki, M.; Oku, H.; Sandoval, J.; Inagaki, M.; Nakatani, K.; Nakaoka, H.; Kawase, R.; Yuasa-Kawase, M.; et al. Adiponectin prevents atherosclerosis by increasing cholesterol efflux from macrophages. Biochem. Biophys. Res. Commun. 2008, 375, 390–394. [Google Scholar] [CrossRef]
- Matsuura, F.; Oku, H.; Koseki, M.; Sandoval, J.; Yuasa-Kawase, M.; Tsubakio-Yamamoto, K.; Masuda, D.; Maeda, N.; Tsujii, K.; Ishigami, M.; et al. Adiponectin accelerates reverse cholesterol transport by increasing high density lipoprotein assembly in the liver. Biochem. Biophys. Res. Commun. 2007, 358, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.; Barrett, P.; Ooi, E.; Ji, J.; Chan, D.; Watts, G. Very low density lipoprotein metabolism and plasma adiponectin as predictors of high-density lipoprotein apolipoprotein A-I kinetics in obese and nonobese men. J. Clin. Endocrinol. Metab. 2009, 94, 989–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variables | T2D Median | (min, max) | Control Median | (min, max) | p-Value |
|---|---|---|---|---|---|
| Number (M:F) | 8(4:4) | 9(4:5) | |||
| Age (years) | 62 | (42, 71) | 55 | (35, 66) | 0.072 |
| Weight (kg) | 91.8 | (76, 121) | 82 | (57.1, 102.4) | 0.175 |
| Body mass index (kg/m2) | 32.2 | (26, 36) | 30 | (24, 32) | 0.175 |
| Systolic blood pressure (mmHg) | 131 | (101, 158) | 123 | (99, 143) | 0.16 |
| Diastolic blood pressure (mmHg) | 72 | (55, 97) | 77 | (60, 90) | 0.69 |
| Glucose (mg/dL) | 110 | (91, 143) | 96.5 | (78, 108) | 0.001 |
| Fasting insulin (mIU/dL) | 16.21 | (12, 39) | 7.0 | (4.1, 16.5) | <0.001 |
| HOMA IR | 5.2 | (3, 10) | 1.6 | (1, 4) | <0.001 |
| HbA1c (%) | 6.4 | (5.8, 6.7) | 5.4 | (5.1, 5.8) | <0.001 |
| Triglycerides (mg/dL) | 94 | (28, 149) | 85 | (50, 173) | 0.64 |
| High-sensitivity C-reactive protein (mg/L) | 2.5 | (0.5, 15) | 1.1 | (0.2, 3.6) | 0.253 |
| Myeloperoxidase activity (mOD/µL/min) | 7.8 | (6.2, 24.1) | 6.0 | (3.3, 17.8) | 0.720 |
| Total cholesterol (mg/dL) | 195 | (122, 237) | 180.5 | (110, 254) | 0.44 |
| HDL-cholesterol (mg/dL) | 48 | (33, 67) | 52 | (34, 79) | 0.75 |
| Lecithin-cholesterol acyltransferase activity | 1.14 | (1, 1.2) | 1.1 | (1, 1.2) | 0.62 |
| Cholesteryl ester transfer protein activity (pmol/mL/min) | 11.4 | (5.6, 17.2) | 9.7 | (5.6, 13.6) | 0.82 |
| Proinflammatory index of HDL (RFU mg HDLc/min) | 19.6 | (13.4, 31) | 19.9 | (12.9, 25.4) | 0.87 |
| Adiponectin (µg/mL) | 2.9 | (1.9, 3.8) | 9.5 | (2.7, 13) | <0.001 |
| Variables | Control | T2D | p-Value |
|---|---|---|---|
| HDL cholesterol pool (mg/kg) | 23.5 ± 6.1 | 23.9 ± 7.4 | NS |
| HDL cholesterol FCR (day−1) | 0.16 ± 0.01 | 0.29 ± 0.08 | 0.008 |
| HDL cholesterol residence time (day) | 8.31 ± 3.52 | 3.76 ± 1.20 | 0.004 |
| HDL cholesterol PR (mg/kg/d) | 3.5 ± 1.8 | 7.2 ± 3.2 | 0.015 |
| Accession Number | Protein | GO-Molecular Function | Control t ½ (hour ± SD) | T2D t ½ (hour ± SD) | p-Value (95% CI) |
|---|---|---|---|---|---|
| Immune Response | |||||
| P01024 | Complement C3 | Endopeptidase inhibitor | 54.5 ± 17.8 | 33.3 ± 13 | 0.002 (−49, −7) |
| P10909 | Apolipoprotein J (Clusterin) | Misfolded protein binding | 26.5 ± 10.6 | 19.8 ± 5.4 | 0.007 (−19, −2) |
| Antioxidant | |||||
| P27169 | Paraoxonase 1 | Antioxidant | 261.6 ± 66.0 | 474.8 ± 1001 | 0.006 (64, 301) |
| Lipid Metabolism | |||||
| P02652 | Apolipoprotein A-II | Cholesterol transporter | 91.9 ± 23.1 | 51.9 ± 17.3 | 0.003 (−59, 14) |
| P02656 | Apolipoprotein C-III | Lipid-binding | 24 ± 5.8 | 21.9 ± 6.3 | 0.6 (−9,4) |
| P06727 | Apolipoprotein A-IV | Antioxidant, cholesterol transporter | 36.5 ± 7.2 | 26 ± 7.2 | 0.004 (−19, −4) |
| P02766 | Transthyretin | Thyroid hormone-binding protein | 37.1 ± 12.9 | 28.3 ± 8.3 | 0.03 (−27, −1) |
| Vitamin Transport | |||||
| P02774 | Vitamin D-binding protein | Actin-binding, vitamin transporter activity | 50.8 ± 14.2 | 30.8 ± 8.3 | 0.02 (−25, −1) |
| Parameters | Lower CI | Upper CI | Correlation (rpb) | p-Value |
|---|---|---|---|---|
| LCAT activity Blood glucose concentration | −1.0 | −0.17 | −0.84 | 0.036 |
| Proinflammatory HDL | −0.98 | 0.08 | −0.75 | 0.07 |
| HDL concentration |
| Parameters | Slope | Slope CI | Intercept | p-Value |
|---|---|---|---|---|
| Proinflammatory HDL (x-variable) | −0.0008 | (−0.0013, −0.0003) | 0.03 | 0.005 |
| ApoA-II kinetics (y-variable) | ||||
| LCAT activity (x-variable) | −0.005 | (−0.012, −0.0018) | 0.007 | 0.04 |
| Paraoxonase 1 kinetics (y-variable) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kheniser, K.G.; Osme, A.; Kim, C.; Ilchenko, S.; Kasumov, T.; Kashyap, S.R. Temporal Dynamics of High-Density Lipoprotein Proteome in Diet-Controlled Subjects with Type 2 Diabetes. Biomolecules 2020, 10, 520. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040520
Kheniser KG, Osme A, Kim C, Ilchenko S, Kasumov T, Kashyap SR. Temporal Dynamics of High-Density Lipoprotein Proteome in Diet-Controlled Subjects with Type 2 Diabetes. Biomolecules. 2020; 10(4):520. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040520
Chicago/Turabian StyleKheniser, Karim G., Abdullah Osme, Chunki Kim, Serguei Ilchenko, Takhar Kasumov, and Sangeeta R. Kashyap. 2020. "Temporal Dynamics of High-Density Lipoprotein Proteome in Diet-Controlled Subjects with Type 2 Diabetes" Biomolecules 10, no. 4: 520. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040520