Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics

Abstract
:1. Introduction
2. Materials and Methods
2.1. Probiotic Cultures and Vegetable Juice Fermentation
2.2. Comprehensive Metabolic Profiling Using GC-MS
2.3. Comprehensive Lipid Profiling Using DI-MS
2.4. Carotenoid Analysis Using LC-MS
2.5. Statistical Analysis
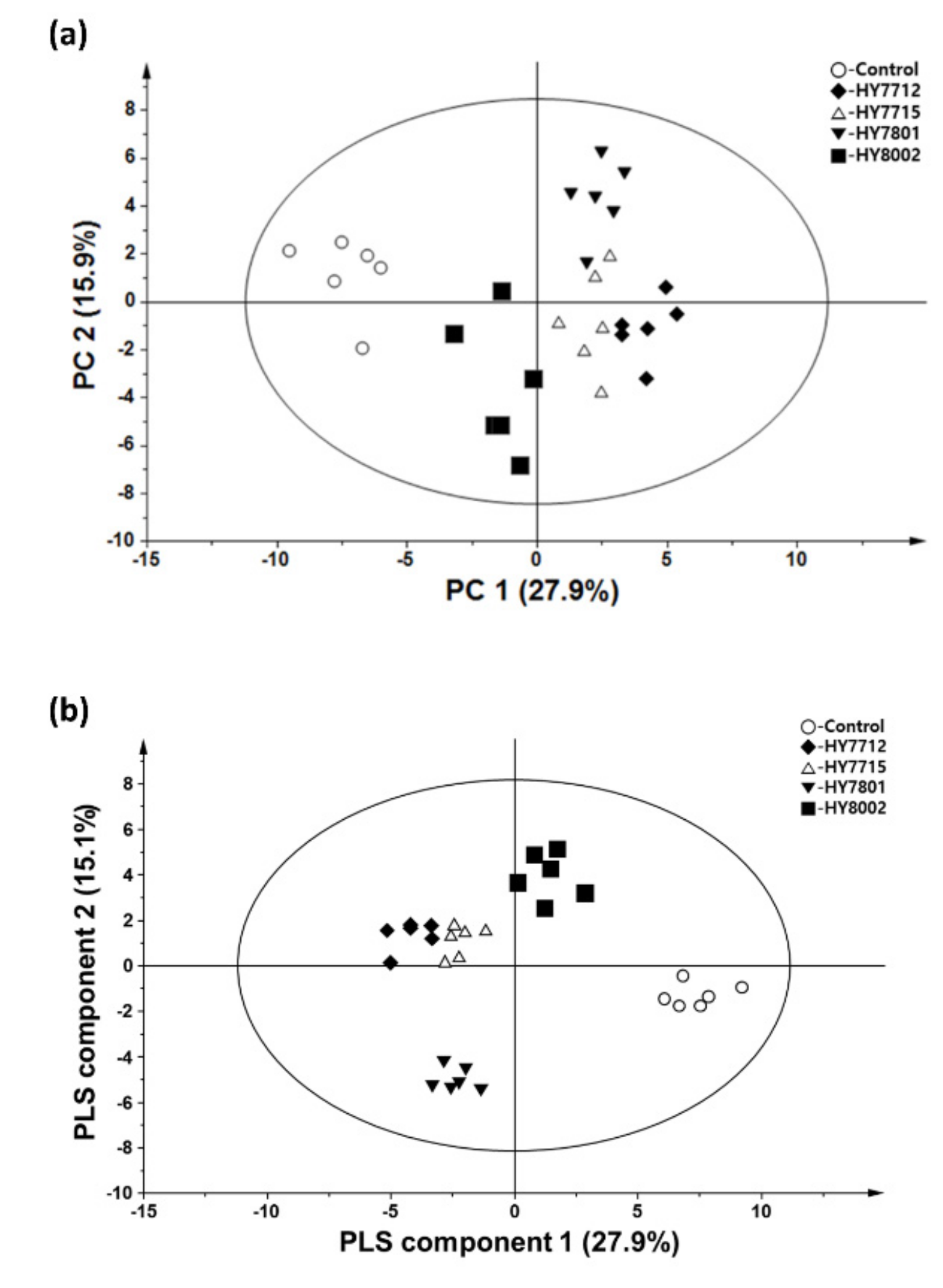
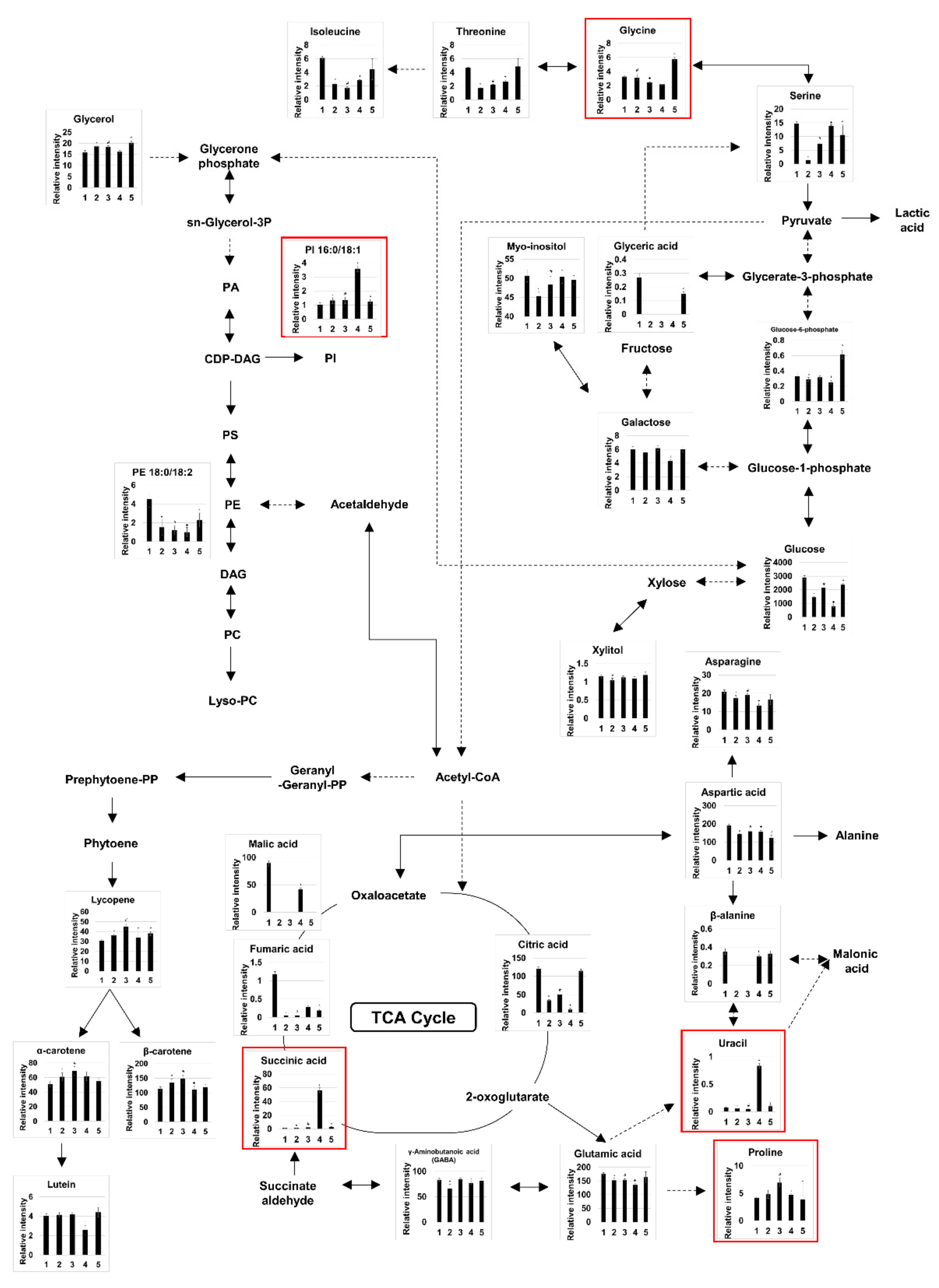
3. Results
3.1. Identification and Quantification of Metabolites and Lipids in Fermented and Non-Fermented VJs Using GC-MS, DI-MS, and LC-MS
3.2. Probiotic Fermentation of VJ Alters its Metabolic and Lipidomic Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Report of a joint FAO/WHO working group on drafting guidelines for the evaluation of probiotics in food. In Guidelines for the Evaluation of Probiotics in Food; FAO/WHO: London, ON, Canada, 2002. [Google Scholar]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013. [Google Scholar] [CrossRef] [Green Version]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligne, B.; Ganzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2019, 73, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Yang, J.; Shao, P.; Lee, R.P.; Huang, J.; Ly, A.; Hsu, M.; Lu, Q.Y.; Thames, G.; Heber, D.; et al. Health benefit of vegetable/fruit juice-based diet: Role of microbiome. Sci. Rep. 2017, 7, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havas, P.; Kun, S.; Styevkó, G.; Slačanac, V.; Hardi, J.; Rezessy-Szabó, J. Fruit and vegetable juice fermentation with bifidobacteria. Acta Alimentaria 2014, 43, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Alissa, E.M.; Ferns, G.A. Dietary fruits and vegetables and cardiovascular diseases risk. Crit. Rev. Food Sci. Nutr. 2017, 57, 1950–1962. [Google Scholar] [CrossRef] [Green Version]
- Kosewski, G.; Gorna, I.; Boleslawska, I.; Kowalowka, M.; Wieckowska, B.; Glowka, A.K.; Morawska, A.; Jakubowski, K.; Dobrzynska, M.; Miszczuk, P.; et al. Comparison of antioxidative properties of raw vegetables and thermally processed ones using the conventional and sous-vide methods. Food Chem. 2018, 240, 1092–1096. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Parveen Rani, R. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014. [Google Scholar] [CrossRef]
- Corona, O.; Randazzo, W.; Miceli, A.; Guarcello, R.; Francesca, N.; Erten, H.; Moschetti, G.; Settanni, L. Characterization of kefir-like beverages produced from vegetable juices. LWT Food Sci. Technol. 2016, 66, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y. Production of fermented kale juices with Lactobacillus strains and nutritional composition. Prev. Nutr. Food Sci. 2017, 22, 231–236. [Google Scholar] [PubMed]
- Tomita, S.; Saito, K.; Nakamura, T.; Sekiyama, Y.; Kikuchi, J. Rapid discrimination of strain-dependent fermentation characteristics among Lactobacillus strains by NMR-based metabolomics of fermented vegetable juice. PLoS ONE 2017, 12, e0182229. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Xu, M.; Zhong, F.; Zhu, J. Rapid differentiation of Lactobacillus species via metabolic profiling. J. Microbiol. Methods 2018, 154, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, S.; Nakamura, T.; Okada, S. NMR- and GC/MS-based metabolomic characterization of sunki, an unsalted fermented pickle of turnip leaves. Food Chem. 2018, 258, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Seo, S.H.; Lee, K.I.; Na, C.S.; Son, H.S. Metabolite profiling of fermented ginseng extracts by gas chromatography mass spectrometry. J. Ginseng Res. 2018, 42, 57–67. [Google Scholar] [CrossRef]
- Chemat, F.; Rombaut, N.; Sicaire, A.G.; Meullemiestre, A.; Fabiano-Tixier, A.S.; Abert-Vian, M. Ultrasound assisted extraction of food and natural products. Mechanisms, techniques, combinations, protocols and applications. A review. Ultrason. Sonochem. 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, H.Y.; Jeon, J.Y.; Kim, D.M.; Zhou, Y.; Lee, J.S.; Lee, H.; Choi, H.K. Effects of coronatine elicitation on growth and metabolic profiles of Lemna paucicostata culture. PLoS ONE 2017, 12, e0187622. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lim, S.R.; Hong, S.J.; Cho, B.K.; Lee, H.; Lee, C.G.; Choi, H.K. Effect of ethephon as an ethylene-releasing compound on the metabolic profile of Chlorella vulgaris. J. Agric. Food Chem. 2016, 64, 4807–4816. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://www.lipidmaps.org/ (accessed on 24 October 2018).
- Bohoyo-Gil, D.; Dominguez-Valhondo, D.; Garcia-Parra, J.; González-Gómez, D. UHPLC as a suitable methodology for the analysis of carotenoids in food matrix. Eur. Food Res. Technol. 2012, 235, 1055–1061. [Google Scholar] [CrossRef]
- Kim, S.H.; Liu, K.H.; Lee, S.Y.; Hong, S.J.; Cho, B.K.; Lee, H.; Lee, C.G.; Choi, H.K. Effects of light intensity and nitrogen starvation on glycerolipid, glycerophospholipid, and carotenoid composition in Dunaliella tertiolecta culture. PLoS ONE 2013, 8, e72415. [Google Scholar] [CrossRef] [Green Version]
- Available online: http://www.metaboanalyst.ca (accessed on 24 October 2018).
- Desjardins, M.-L.; Roy, D.; Goulet, J. Growth of bifidobacteria and their enzyme profiles. J. Dairy Sci. 1990, 73, 299–307. [Google Scholar] [CrossRef]
- Premi, L.; Sandine, W.E.; Elliker, P.R. Lactose-hydrolyzing enzymes of Lactobacillus species. Appl. Microbiol. 1972, 24, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Caspritz, G.; Radler, F. Malolactic enzyme of Lactobacillus plantarum. Purification, properties, and distribution among bacteria. J. Biol. Chem. 1983, 258, 4907–4910. [Google Scholar]
- Pham, P.L.; Dupont, I.; Roy, D.; Lapointe, G.; Cerning, J. Production of exopolysaccharide by Lactobacillus rhamnosus R and analysis of its enzymatic degradation during prolonged fermentation. Appl. Environ. Microbiol. 2000, 66, 2302–2310. [Google Scholar] [CrossRef] [Green Version]
- Degeest, B.; Janssens, B.; De Vuyst, L. Exopolysaccharide (EPS) biosynthesis by Lactobacillus sakei 0–1: Production kinetics, enzyme activities and EPS yields. J. Appl. Microbiol. 2001, 91, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Palasz, A.; Ciez, D. In search of uracil derivatives as bioactive agents. Uracils and fused uracils: Synthesis, biological activity and applications. Eur. J. Med. Chem. 2015, 97, 582–611. [Google Scholar] [CrossRef]
- Elli, M.; Zink, R.; Rytz, A.; Reniero, R.; Morelli, L. Iron requirement of Lactobacillus spp. in completely chemically defined growth media. J. Appl. Microbiol. 2000, 88, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Patra, J.K.; Lee, S.Y.; Kim, C.; Park, J.G.; Baek, K.H. Analysis of metabolomic profile of fermented Orostachys japonicus A. Berger by capillary electrophoresis time of flight mass spectrometry. PLoS ONE 2017, 12, e0181280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, E.M. Escherichia coli-derived uracil increases the antibacterial activity and growth rate of Lactobacillus plantarum. J. Microbiol. Biotechnol. 2016, 26, 975–987. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zeng, S.Y.; Leu, Y.L.; Tsai, T.Y. Antihypertensive effect of a combination of uracil and glycerol derived from Lactobacillus plantarum strain TWK10-fermented soy milk. J. Agric. Food Chem. 2015, 63, 7333–7342. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.C.; Sun, D.; Chen, G.; Sharma, A.K.; Amin, S.; Ropson, I.J.; Spratt, T.E.; Lazarus, P. Characterization of dibenzo[a,l]pyrene-trans-11,12-diol(dibenzo[def,p]chrysene) glucuronidation by UDP-glucuronosyltransferases. Chem. Res. Toxicol. 2011, 24, 1549–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W.; et al. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-kB signaling. PLoS ONE 2013, 8, e73877. [Google Scholar]
- Zimin, Y.S.; Borisova, N.; Timerbaeva, G.; Gimadieva, A.; Mustafin, A. Preparation, toxicity, and anti-inflammatory activity of complexes of uracil derivatives with polyfunctional acids. Pharm. Chem. J. 2017, 50, 649–653. [Google Scholar] [CrossRef]
- Kim, J.-E.; Chae, C.S.; Kim, G.-C.; Hwang, W.; Hwang, J.-S.; Hwang, S.-M.; Kim, Y.; Ahn, Y.-T.; Park, S.-G.; Jun, C.-D. Lactobacillus helveticus suppresses experimental rheumatoid arthritis by reducing inflammatory T cell responses. J. Funct. Foods 2015, 13, 350–362. [Google Scholar] [CrossRef]
- Joo, H.M.; Kim, K.A.; Myoung, K.S.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Han, M.J.; Kim, D.H. Lactobacillus helveticus HY7801 ameliorates vulvovaginal candidiasis in mice by inhibiting fungal growth and NF-κB activation. Int. Immunopharmacol. 2012, 14, 39–46. [Google Scholar] [CrossRef]
- Hong, Y.S.; Ahn, Y.T.; Park, J.C.; Lee, J.H.; Lee, H.; Huh, C.S.; Kim, D.H.; Ryu, D.H.; Hwang, G.S. 1H NMR-based metabonomic assessment of probiotic effects in a colitis mouse model. Arch. Pharm. Res. 2010, 33, 1091–1101. [Google Scholar] [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Zeikus, J.; Jain, M.; Elankovan, P. Biotechnology of succinic acid production and markets for derived industrial products. Appl. Microbiol. Biotechnol. 1999, 51, 545–552. [Google Scholar] [CrossRef]
- Dudley, E.G.; Steele, J.L. Succinate production and citrate catabolism by Cheddar cheese nonstarter lactobacilli. J. Appl. Microbiol. 2005, 98, 14–23. [Google Scholar] [CrossRef]
- Hillier, A.J. The metabolism of [14C]bicarbonate by Streptococcus lactis: The synthesis of succinic acid. J. Dairy Res. 1978, 45, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, R.; Adriany, T.; Verbrugghe, K.; De Vuyst, L. Kinetic analysis of bifidobacterial metabolism reveals a minor role for succinic acid in the regeneration of NAD+ through its growth-associated production. Appl. Environ. Microbiol. 2006, 72, 5204–5210. [Google Scholar] [CrossRef] [Green Version]
- Giorgi-Coll, S.; Amaral, A.I.; Hutchinson, P.J.A.; Kotter, M.R.; Carpenter, K.L.H. Succinate supplementation improves metabolic performance of mixed glial cell cultures with mitochondrial dysfunction. Sci. Rep. 2017, 7, 1003. [Google Scholar] [CrossRef] [Green Version]
- Jalloh, I.; Helmy, A.; Howe, D.J.; Shannon, R.J.; Grice, P.; Mason, A.; Gallagher, C.N.; Stovell, M.G.; van der Heide, S.; Murphy, M.P.; et al. Focally perfused succinate potentiates brain metabolism in head injury patients. J. Cereb. Blood Flow Metab. 2017, 37, 2626–2638. [Google Scholar] [CrossRef]
- Iplik, E.S.; Catmakas, T.; Cakmakoglu, B. A new target for the treatment of endometrium cancer by succinic acid. Cell Mol. Biol. 2018, 64, 60–63. [Google Scholar] [CrossRef]
- Zarubina, I.V.; Lukk, M.V.; Shabanov, P.D. Antihypoxic and antioxidant effects of exogenous succinic acid and aminothiol succinate-containing antihypoxants. Bull. Exp. Biol. Med. 2012, 153, 336–339. [Google Scholar] [CrossRef]
- Carman, G.M.; Henry, S.A. Phosphatidic acid plays a central role in the transcriptional regulation of glycerophospholipid synthesis in Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 37293–37297. [Google Scholar] [CrossRef] [Green Version]
- Henry, S.A.; Kohlwein, S.D.; Carman, G.M. Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae. Genetics 2012, 190, 317–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, Z.; Kohlwein, S.; Paltauf, F. Role of phosphatidylinositol (PI) in ethanol production and ethanol tolerance by a high ethanol producing yeast. J. Ind. Microbiol. Biotechnol. 1999, 22, 58–63. [Google Scholar] [CrossRef]
- Holub, B.J. The nutritional significance, metabolism, and function of myo-inositol and phosphatidylinositol in health and disease. Adv. Nutr. Res. 1982, 4, 107–141. [Google Scholar] [PubMed]
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, J.W.; Neville, T.A.; Rouillard, P.; Harder, Z.; Beanlands, D.S.; Sparks, D.L. Phosphatidylinositol increases HDL-C levels in humans. J. Lipid Res. 2005, 46, 350–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunton, J.A.; Baldwin, M.P.; Hanna, R.A.; Bertolo, R.F. Proline supplementation to parenteral nutrition results in greater rates of protein synthesis in the muscle, skin, and small intestine in neonatal Yucatan miniature piglets. J. Nutr. 2012, 142, 1004–1008. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wu, G. Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef]
- Barbul, A. Proline precursors to sustain mammalian collagen synthesis. J. Nutr. 2008, 138, 2021s–2024s. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Dickman, M.B.; Becker, D.F. Proline biosynthesis is required for endoplasmic reticulum stress tolerance in Saccharomyces cerevisiae. J. Biol. Chem. 2014, 289, 27794–27806. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Kato, J.; Horie, S.; Komatsubara, S.; Kisumi, M.; Chibata, I. Production of L-proline by Kurthia catenaforma. Appl. Microbiol. 1968, 16, 1200–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prockop, D.J.; Juva, K. Synthesis of hydroxyproline in vitro by the hydroxylation of proline in a precursor of collagen. Proc. Natl. Acad. Sci. USA 1965, 53, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Guo, Q.; Yin, Y.; Blachier, F.; Kong, X. Dietary proline supplementation alters colonic luminal microbiota and bacterial metabolite composition between days 45 and 70 of pregnancy in Huanjiang mini-pigs. J. Anim. Sci. Biotechnol. 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, K.; Stoimenova, A.; Obreshkova, D.; Saso, L. Biotechnology in the production of pharmaceutical industry ingredients: Amino acids. Biotechnol. Biotechnol. Equip. 2013, 27, 3620–3626. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, H.-J.; Park, S.-K. Amino acids analysis during lactic acid fermentation by single strain cultures of lactobacilli and mixed culture starter made from them. Afr. J. Biotechnol. 2014, 13, 2867–2873. [Google Scholar]
- Bannai, M.; Kawai, N.; Ono, K.; Nakahara, K.; Murakami, N. The effects of glycine on subjective daytime performance in partially sleep-restricted healthy volunteers. Front Neurol. 2012, 3, 61. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.L.; Madren, J.; Bunzendahl, H.; Thurman, R.G. Dietary glycine inhibits the growth of B16 melanoma tumors in mice. Carcinogenesis 1999, 20, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Fernandez, J.; Maldonado-Barragan, A.; Caballero-Guerrero, B.; Hornero-Mendez, D.; Ruiz-Barba, J.L. Carotenoid production in Lactobacillus plantarum. Int. J. Food Microbiol. 2010, 140, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Arab, L.; Steck, S. Lycopene and cardiovascular disease. Am. J. Clin. Nutr. 2000, 71, 1691S–1695S. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Vidmantiene, D.; Juodeikiene, G.; Viskelis, P.; Urbonaviciene, D. Lactic acid fermentation of tomato: Effects on cis/trans lycopene isomer ratio, β-carotene mass fraction and formation of L (+)-and D (–)-lactic acid. Food Technol. Biotech. 2013, 51, 471. [Google Scholar]
- Breithaupt, D.E.; Schwack, W.; Wolf, G.; Hammes, W.P. Characterization of the triterpenoid 4, 4′-diaponeurosporene and its isomers in food-associated bacteria. Eur. Food Res. Technol. 2001, 213, 231–233. [Google Scholar] [CrossRef]
- Sanchez-Contreras, A.; Jimenez, M.; Sanchez, S. Bioconversion of lutein to products with aroma. Appl. Microbiol. Biotechnol. 2000, 54, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ahn, Y.T.; Park, S.H.; Park, D.Y.; Jin, Y.W.; Kim, C.S.; Sung, S.H.; Huh, C.S.; Kim, D.H. Lactobacillus plantarum HY7712 protects against the impairment of NK-cell activity caused by whole-body γ-irradiation in mice. J. Microbiol. Biotechnol. 2014, 24, 127–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.E.; Joh, E.H.; Lee, H.Y.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Han, M.J.; Kim, D.H. Lactobacillus plantarum HY7712 ameliorates cyclophosphamide-induced immunosuppression in mice. J. Microbiol. Biotechnol. 2013, 23, 414–421. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| No. | Compound | m/z | RT (min) | Fragmentation Ions (m/z) | TMS | Control | L. plantarum (HY7712) | L. plantarum (HY7715) | L. helveticus (HY7801) | B. lactis (HY8002) |
|---|---|---|---|---|---|---|---|---|---|---|
| Amino acids | ||||||||||
| 1 | β-alanine | 174 | 17.37 | 100, 174, 248, 290 | 3 | 0.352 ± 0.032 | ND | ND | 0.302 ± 0.011 + | 0.328 ± 0.032 |
| 2 | γ-aminobutanoic acid | 174 | 19.85 | 174, 216, 246, 304 | 3 | 83.630 ± 2.806 | 66.176 ± 8.063 * | 84.661 ± 1.777 | 77.351 ± 2.740 + | 81.219 ± 6.131 |
| 3 | Alanine | 116 | 8.80 | 100, 116, 190, 218 | 2 | 50.422 ± 1.816 | 50.480 ± 2.053 | 50.026 ± 5.861 | 53.394 ± 3.054 | 40.845 ± 24.358 |
| 4 | Asparagine | 116 | 23.13 | 116, 132, 188, 231 | 3 | 20.862 ± 1.038 | 17.253 ± 1.558 * | 18.952 ± 0.744 # | 13.289 ± 1.015 + | 16.524 ± 2.890 ^ |
| 5 | Aspartic acid | 232 | 19.68 | 100, 202, 218, 232 | 3 | 190.073 ± 8.232 | 142.789 ± 2.909 * | 159.894 ± 1.455 # | 157.900 ± 5.864 + | 122.123 ± 15.734 ^ |
| 6 | Glutamic acid | 246 | 22.06 | 128, 156, 246, 348 | 3 | 175.620 ± 4.918 | 152.488 ± 7.547 * | 154.917 ± 3.394 # | 134.961 ± 5.251 + | 163.626 ± 19.308 |
| 7 | Glycine | 174 | 14.19 | 86, 174, 248, 276 | 2 | 3.261 ± 0.147 | 3.118 ± 0.448 | 2.453 ± 0.061 # | 2.163 ± 0.071 + | 5.763 ± 0.237 ^ |
| 8 | Isoleucine | 158 | 13.88 | 100, 158, 218, 232 | 2 | 6.115 ± 0.241 | 2.297 ± 0.237 * | 1.711 ± 0.171 # | 2.861 ± 0.080 + | 4.462 ± 1.543 |
| 9 | Proline | 142 | 13.96 | 100, 142, 144, 216 | 2 | 4.192 ± 0.234 | 4.804 ± 0.729 | 6.941 ± 0.784 # | 4.736 ± 0.149 + | 3.890 ± 3.297 |
| 10 | Pyroglutamic acid | 156 | 19.62 | 133, 156, 230, 258 | 2 | 562.497 ± 15.928 | 483.337 ± 18.371 * | 508.725 ± 11.686 # | 521.194 ± 20.262 + | 526.650 ± 16.360 ^ |
| 11 | Serine | 204 | 15.68 | 100, 188, 204, 218 | 3 | 14.700 ± 0.692 | 1.474 ± 0.232 * | 7.357 ± 0.522 # | 13.816 ± 0.457 + | 10.529 ± 3.635 ^ |
| 12 | Threonine | 218 | 16.32 | 101, 117, 218, 291 | 3 | 4.661 ± 0.119 | 1.712 ± 0.157 * | 2.217 ± 0.037 # | 2.617 ± 0.156 + | 4.863 ± 1.161 |
| 13 | Valine | 144 | 11.80 | 100, 133, 144, 218 | 2 | 10.732 ± 0.785 | 5.977 ± 0.323 * | 5.387 ± 0.257 # | 7.584 ± 0.239 + | 7.687 ± 3.507 |
| Fatty acids | ||||||||||
| 14 | 1-Monopalmitin | 371 | 45.26 | 103, 129, 205, 371 | 2 | 4.647 ± 0.403 | 3.781 ± 1.278 | 3.705 ± 0.421 # | 2.835 ± 0.433 + | 4.372 ± 1.145 |
| 15 | Linoleic acid | 75 | 37.82 | 67, 75, 81, 337 | 1 | 0.528 ± 0.074 | 0.394 ± 0.089 * | 0.268 ± 0.059 # | 0.357 ± 0.044 + | 0.337 ± 0.044 ^ |
| 16 | Palmitic acid | 117 | 33.33 | 117, 132, 145, 313 | 1 | 0.673 ± 0.049 | 0.636 ± 0.057 | 0.649 ± 0.053 | 0.617 ± 0.090 | 0.707 ± 0.080 |
| 17 | Stearic acid | 117 | 38.66 | 117, 132, 145, 341 | 1 | 0.240 ± 0.039 | 0.367 ± 0.083 * | 0.286 ± 0.103 | 0.267 ± 0.081 | 0.323 ± 0.076 |
| Organic acids | ||||||||||
| 18 | Acetic acid | 177 | 8.12 | 133, 161, 177, 205 | 2 | 0.143 ± 0.012 | 0.124 ± 0.008 * | 0.137 ± 0.020 | 0.143 ± 0.016 | 0.150 ± 0.017 |
| 19 | Citric acid | 273 | 26.52 | 273, 347, 363, 375 | 4 | 120.814 ± 4.491 | 34.142 ± 1.741 * | 50.563 ± 1.841 # | 9.052 ± 4.326 + | 114.971 ± 4.669 |
| 20 | Fumaric acid | 245 | 15.46 | 115, 132, 143, 245 | 2 | 1.181 ± 0.074 | 0.043 ± 0.005 * | 0.045 ± 0.004 # | 0.287 ± 0.033 + | 0.189 ± 0.036 ^ |
| 21 | Lactic acid | 117 | 7.73 | 117, 133, 191, 219 | 2 | 21.595 ± 1.210 | 602.047 ± 19.653 * | 563.702 ± 33.204 # | 547.899 ± 14.857 + | 420.494 ± 27.212 ^ |
| 22 | Malic acid | 233 | 18.93 | 133, 189, 233, 245 | 3 | 89.641 ± 5.199 | ND | ND | 41.678 ± 2.598 + | ND |
| 23 | Malonic acid | 75 | 11.51 | 66, 75, 133, 233 | 2 | 0.441 ± 0.035 | 0.429 ± 0.023 | 0.425 ± 0.028 | 0.419 ± 0.029 | 0.533 ± 0.115 |
| 24 | Succinic acid | 247 | 14.48 | 75, 129, 172, 247 | 2 | 1.544 ± 0.071 | 1.768 ± 0.071 * | 2.462 ± 0.108 # | 56.616 ± 3.446 + | 3.044 ± 0.131 ^ |
| 25 | Tartaric acid | 292 | 22.43 | 189, 219, 292, 423 | 4 | 0.776 ± 0.028 | 0.602 ± 0.084 * | 0.638 ± 0.037 # | 0.686 ± 0.019 + | 0.683 ± 0.034 ^ |
| Sugars | ||||||||||
| 26 | Fructose | 217 | 26.34 | 204, 217, 319, 437 | 5 | 526.553 ± 32.602 | 553.891 ± 8.751 | 520.415 ± 26.583 | 512.824 ± 9.722 | 521.351 ± 26.811 |
| 103 | 28.01 | 103, 133, 217, 307 | 5(MeOX) | |||||||
| 28.32 | ||||||||||
| 27 | Galactose | 204 | 28.89 | 129, 191, 204, 217 | 5 | 6.029 ± 0.373 | 5.561 ± 0.336 | 6.183 ± 0.352 | 4.300 ± 0.139 + | 5.980 ± 0.365 |
| 28 | Glucose | 204 | 28.61 | 129, 191, 204, 217 | 5 | 2905.897 ± 138.257 | 1486.807 ± 39.829 * | 2156.601 ± 58.234 # | 805.171 ± 102.179 + | 2385.734 ± 64.071 ^ |
| 31.40 | ||||||||||
| 319 | 28.72 | 160, 205, 217, 319 | 5(MeOX) | |||||||
| 29 | Glucose-6-phosphate | 204 | 40.26 | 204, 217, 299, 387 | 6 | 0.329 ± 0.032 | 0.288 ± 0.025 * | 0.316 ± 0.021 | 0.251 ± 0.026 + | 0.618 ± 0.058 ^ |
| 41.74 | ||||||||||
| 30 | Sedoheptulose | 319 | 35.74 | 205, 217, 262, 319 | 6(MeOX) | 50.748 ± 3.714 | 42.976 ± 1.234 * | 47.967 ± 1.442 | 49.610 ± 0.918 | 44.614 ± 3.511 ^ |
| 35.89 | ||||||||||
| 31 | Sucrose | 361 | 46.06 | 103, 217, 361, 437 | 8 | 437.362 ± 192.453 | 556.267 ± 20.170 | 535.635 ± 35.941 | 509.509 ± 14.353 | 507.215 ± 24.494 |
| 32 | Xylose | 103 | 22.85 | 103, 189, 217, 307 | 4(MeOX) | 0.700 ± 0.042 | 0.686 ± 0.104 | 0.708 ± 0.051 | 0.715 ± 0.047 | 0.783 ± 0.022 ^ |
| Sugar acids | ||||||||||
| 33 | Glyceric acid | 189 | 14.88 | 103, 189, 205, 292 | 3 | 0.268 ± 0.024 | ND | ND | ND | 0.148 ± 0.016 ^ |
| 34 | Threonic acid | 292 | 20.66 | 117, 205, 220, 292 | 4 | 0.569 ± 0.030 | 0.455 ± 0.031 * | 0.502 ± 0.026 # | 0.494 ± 0.026 + | 0.554 ± 0.029 |
| Sugar alcohols | ||||||||||
| 35 | Erythritol | 217 | 19.22 | 103, 117, 205, 217 | 4 | 13.869 ± 0.680 | 1.057 ± 0.045 * | 1.119 ± 0.054 # | 6.792 ± 0.327 + | 1.647 ± 0.436 ^ |
| 19.39 | ||||||||||
| 36 | Glycerol | 205 | 13.41 | 103, 117, 133, 205 | 3 | 16.011 ± 0.751 | 18.614 ± 0.157 * | 18.262 ± 0.573 # | 16.246 ± 0.480 | 20.278 ± 0.708 ^ |
| 37 | Mannitol | 319 | 29.66 | 103, 205, 217, 319 | 6 | 11.188 ± 0.638 | 9.783 ± 0.434 * | 10.961 ± 0.551 | 10.554 ± 0.242 + | 18.955 ± 12.144 |
| 38 | Myo-Inositol | 305 | 34.45 | 191, 217, 305, 318 | 6 | 50.657 ± 1.580 | 45.353 ± 1.026 * | 48.372 ± 2.080 # | 50.395 ± 1.749 | 49.615 ± 1.095 |
| 39 | Xylitol | 217 | 24.25 | 103, 205, 217, 307 | 5 | 1.149 ± 0.043 | 1.050 ± 0.050 * | 1.122 ± 0.047 | 1.096 ± 0.051 | 1.185 ± 0.082 |
| Others | ||||||||||
| 40 | Phosphoric acid | 299 | 13.30 | 133, 211, 299, 314 | 3 | 262.434 ± 9.607 | 186.249 ± 12.682 * | 201.558 ± 3.812 # | 184.653 ± 4.847 + | 217.457 ± 7.081 ^ |
| 41 | Uracil | 241 | 15.03 | 99, 113, 241, 255 | 2 | 0.075 ± 0.011 | 0.057 ± 0.005 * | 0.054 ± 0.006# | 0.835 ± 0.046 + | 0.099 ± 0.064 |
| No. | Lipid Species | Ion Species | m/z | Control | L. plantarum (HY7712) | L. plantarum (HY7715) | L. helveticus (HY7801) | B. lactis (HY8002) |
|---|---|---|---|---|---|---|---|---|
| Positive ion mode | ||||||||
| Monogalactosyldiacylglycerol (MGDG) | ||||||||
| 1 | MGDG 18:2/18:3 | [M + Na]+ | 799 | 1.93 ± 0.62 | 1.74 ± 0.43 | 1.67 ± 0.47 | 1.88 ± 0.31 | 1.55 ± 0.52 |
| 2 | MGDG 18:2/18:2 | [M + Na]+ | 801 | 14.42 ± 4.00 | 13.24 ± 3.19 | 12.68 ± 3.14 | 13.64 ± 1.71 | 11.89 ± 3.22 |
| 3 | MGDG 18:1/18:2 | [M + Na]+ | 803 | 11.51 ± 3.39 | 10.38 ± 1.90 | 9.52 ± 1.93 | 10.37 ± 0.93 | 9.28 ± 2.10 |
| Lysophosphatidylcholine (Lyso-PC) | ||||||||
| 4 | Lyso-PC 18:2 | [M + H]+ | 520 | 5.30 ± 1.04 | 4.18 ± 1.05 | 4.36 ± 0.96 | 4.56 ± 0.70 | 3.73 ± 0.70 ^ |
| 5 | Lyso-PC 18:1 | [M + H]+ | 522 | 6.56 ± 1.60 | 6.53 ± 1.16 | 6.87 ± 1.16 | 6.38 ± 0.64 | 6.37 ± 1.01 |
| 6 | Lyso-PC 22:5 | [M + Na]+ | 592 | 2.81 ± 0.61 | 2.55 ± 0.52 | 2.31 ± 0.52 | 2.80 ± 0.38 | 2.39 ± 0.56 |
| Phosphatidylcholine (PC) | ||||||||
| 7 | PC 18:2/18:2 | [M + H]+ | 782 | 4.29 ± 0.50 | 4.16 ± 0.23 | 4.08 ± 0.28 | 4.06 ± 0.30 | 4.10 ± 0.30 |
| Phosphatidylethanolamine (PE) | ||||||||
| 8 | PE 16:0/20:0 | [M + H]+ | 748 | 1.17 ± 0.30 | 1.05 ± 0.11 | 0.98 ± 0.14 | 1.08 ± 0.11 | 1.08 ± 0.22 |
| Triacylglycerol (TG) | ||||||||
| 9 | TG 16:0/18:2/18:2 | [M + NH4]+ | 872 | 3.88 ± 0.26 | 4.19 ± 0.42 | 4.13 ± 0.49 | 4.17 ± 0.59 | 4.09 ± 0.31 |
| 10 | TG 18:2/18:2/18:3 | [M + NH4]+ | 894 | 2.85 ± 0.19 | 3.03 ± 0.32 | 3.01 ± 0.30 | 3.03 ± 0.39 | 2.98 ± 0.28 |
| 11 | TG 18:2/18:2/18:2 | [M + NH4]+ | 896 | 9.76 ± 0.61 | 10.66 ± 1.03 | 10.45 ± 1.07 | 10.49 ± 1.31 | 10.47 ± 0.83 |
| 12 | TG 18:1/18:2/18:2 | [M + NH4]+ | 898 | 3.33 ± 0.18 | 3.73 ± 0.35 | 3.68 ± 0.46 | 3.64 ± 0.56 | 3.64 ± 0.28 |
| 13 | TG 18:1/18:1/18:2 | [M + NH4]+ | 900 | 1.30 ± 0.04 | 1.46 ± 0.16 | 1.46 ± 0.21 | 1.41 ± 0.20 | 1.42 ± 0.13 |
| Negative ion mode | ||||||||
| Phosphatic acid (PA) | ||||||||
| 14 | PA 16:0/18:2 | [M − H]- | 671 | 1.81 ± 0.27 | 1.76 ± 0.21 | 1.81 ± 0.43 | 1.84 ± 0.30 | 1.67 ± 0.13 |
| 15 | PA 18:2/18:2 | [M − H]- | 695 | 1.74 ± 0.27 | 1.56 ± 0.35 | 1.48 ± 0.35 | 1.69 ± 0.32 | 1.44 ± 0.19 |
| Phosphatidylethanolamine (PE) | ||||||||
| 16 | PE 16:0/18:2 | [M − H]- | 714 | 1.46 ± 0.06 | 1.35 ± 0.08 | 1.35 ± 0.04 # | 1.41 ± 0.09 | 1.41 ± 0.10 |
| 17 | PE 18:2/18:2 | [M − H]- | 738 | 1.23 ± 0.05 | 3.75 ± 2.19 * | 2.72 ± 0.69 # | 4.43 ± 2.06 + | 1.96 ± 0.31 ^ |
| 18 | PE 18:1/18:2 | [M − H]- | 740 | 0.73 ± 0.05 | 1.18 ± 0.83 | 0.76 ± 0.17 | 1.15 ± 0.70 + | 0.62 ± 0.09 ^ |
| 19 | PE 18:0/18:2 | [M − H]- | 742 | 4.50 ± 0.84 | 1.51 ± 0.85 * | 1.22 ± 0.49 # | 1.01 ± 0.51 + | 2.29 ± 0.75 ^ |
| Phosphatidylglycerol (PG) | ||||||||
| 20 | PG 16:0/18:2 | [M − H]- | 745 | 1.28 ± 0.12 | 1.57 ± 0.25 * | 1.49 ± 0.30 | 1.66 ± 0.21 + | 1.47 ± 0.20 |
| Phosphatidylserine (PS) | ||||||||
| 21 | PS 18:2/20:0 | [M − H]- | 814 | 0.23 ± 0.04 | 0.33 ± 0.21 | 0.21 ± 0.05 | 0.15 ± 0.08 + | 0.27 ± 0.04 |
| 22 | PS 18:2/22:0 | [M − H]- | 842 | 0.75 ± 0.12 | 0.98 ± 0.18 * | 0.92 ± 0.09 # | 0.91 ± 0.11 | 0.98 ± 0.14 ^ |
| Phosphatidylinositol (PI) | ||||||||
| 23 | PI 16:0/18:2 | [M − H]- | 833 | 6.34 ± 0.86 | 7.59 ± 1.00 | 7.44 ± 1.13 | 8.04 ± 1.21 + | 7.50 ± 1.05 |
| 24 | PI 16:0/18:1 | [M − H]- | 835 | 1.01 ± 0.15 | 1.32 ± 0.21 * | 1.34 ± 0.23 # | 3.58 ± 0.21 + | 1.24 ± 0.15 ^ |
| Compound | Formula | RT (min) | m/z [M + H] + | Control | L. plantarum (HY7712) | L. plantarum (HY7715) | L. helveticus (HY7801) | B. lactis (HY8002) |
|---|---|---|---|---|---|---|---|---|
| LUT | C40H56O2 | 5.61 | 569.4 | 4.0 ± 0.2 | 4.2 ± 0.3 | 4.2 ± 0.1 | 2.6 ± 0.5 + | 4.4 ± 0.5 |
| LYC | C40H56 | 9.32 | 537.4 | 30.8 ± 0.9 | 36.3 ± 0.6 * | 45.1 ± 3.0 # | 34.0 ± 9.7 | 38.2 ± 2.1 ^ |
| α-CAR | C40H56 | 10.95 | 537.4 | 50.9 ± 3.5 | 60.9 ± 5.7 * | 69.0 ± 5.9 # | 61.8 ± 6.2 + | 54.8 ± 4.0 |
| β-CAR | C40H56 | 11.12 | 537.4 | 113.1 ± 7.6 | 134.4 ± 13.4 * | 147.7 ± 13.5 # | 111.3 ± 12.6 | 120.2 ± 8.7 |
| No. | Pathway Name | Compound a | Total b | Hits c | p d | Impact e |
|---|---|---|---|---|---|---|
| 1 | Alanine, aspartate and glutamate metabolism | alanine, aspartic acid, glutamic acid, asparagine, succinic acid, γ-aminobutanoic acid, fumaric acid | 20 | 7 | 8.18 × 10−6 | 0.60 |
| 2 | Glycine, serine and threonine metabolism | glycine, serine, threonine, glyceric acid, aspartic acid | 28 | 5 | 5.40× 10−3 | 0.42 |
| 3 | Citrate cycle (TCA cycle) | citric acid, fumaric acid, malic acid, succinic acid | 20 | 4 | 8.64 × 10−3 | 0.20 |
| 4 | Aminoacyl-tRNA biosynthesis | asparagine, glycine, aspartic acid, serine, valine, alanine, threonine, proline, glutamic acid, isoleucine | 66 | 10 | 2.52 × 10−4 | 0.18 |
| 5 | Starch and sucrose metabolism | fructose, glucose, glucose-6-phosphate, sucrose, xylose | 30 | 5 | 7.35 × 10−3 | 0.15 |
| 6 | Arginine and proline metabolism | aspartic acid, fumaric acid, proline, glutamic acid, γ-aminobutanoic acid | 40 | 5 | 2.47 × 10−2 | 0.10 |
| No. | Compound | VIP Value |
|---|---|---|
| 1 | γ-aminobutanoic acid | 1.71 |
| 2 | Glycine | 1.51 |
| 3 | Glucose-6-phosphate | 1.43 |
| 4 | Uracil | 1.42 |
| 5 | β-alanine | 1.42 |
| 6 | Succinic acid | 1.41 |
| 7 | Linoleic acid | 1.41 |
| 8 | Aspartic acid | 1.39 |
| 9 | Galactose | 1.38 |
| 10 | Phosphatidylinositol (PI) 16:0/18:1 | 1.36 |
| 11 | Proline | 1.35 |
| 12 | Asparagine | 1.33 |
| 13 | Serine | 1.29 |
| 14 | Glycerol | 1.28 |
| 15 | Sedoheptulose | 1.23 |
| 16 | Glucose | 1.23 |
| 17 | Myo-inositol | 1.20 |
| 18 | Malic acid | 1.16 |
| 19 | Erythritol | 1.14 |
| 20 | Isoleucine | 1.13 |
| 21 | Threonine | 1.13 |
| 22 | Fructose | 1.12 |
| 23 | Fumaric acid | 1.08 |
| 24 | Citric acid | 1.08 |
| 25 | Glyceric acid | 1.03 |
| 26 | Malonic acid | 1.03 |
| 27 | Xylitol | 1.01 |
| 28 | Phosphatidylethanolamine (PE) 18:0/18:2 | 1.00 |
| 29 | Glutamic acid | 1.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, H.-J.; Lee, H.; Na, G.; Jung, H.; Kim, D.-G.; Shin, S.-I.; Jung, S.-E.; Choi, I.-d.; Lee, J.-H.; Sim, J.-H.; et al. Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics. Biomolecules 2020, 10, 725. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050725
Chung H-J, Lee H, Na G, Jung H, Kim D-G, Shin S-I, Jung S-E, Choi I-d, Lee J-H, Sim J-H, et al. Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics. Biomolecules. 2020; 10(5):725. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050725
Chicago/Turabian StyleChung, Hyuk-Jin, Hwanhui Lee, Guknam Na, Heechul Jung, Dong-Gun Kim, Sang-Ick Shin, Seong-Eun Jung, Il-dong Choi, Jae-Hwan Lee, Jae-Hun Sim, and et al. 2020. "Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics" Biomolecules 10, no. 5: 725. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050725