Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Testing
2.2.1. Open Field
2.2.2. Muscle Endurance
2.2.3. The Rotarod Test
2.2.4. Sunflower Seed Task and Vermicelli Handling Test
2.2.5. Bilateral Tactile Stimulation
2.2.6. Light-Dark Box Test
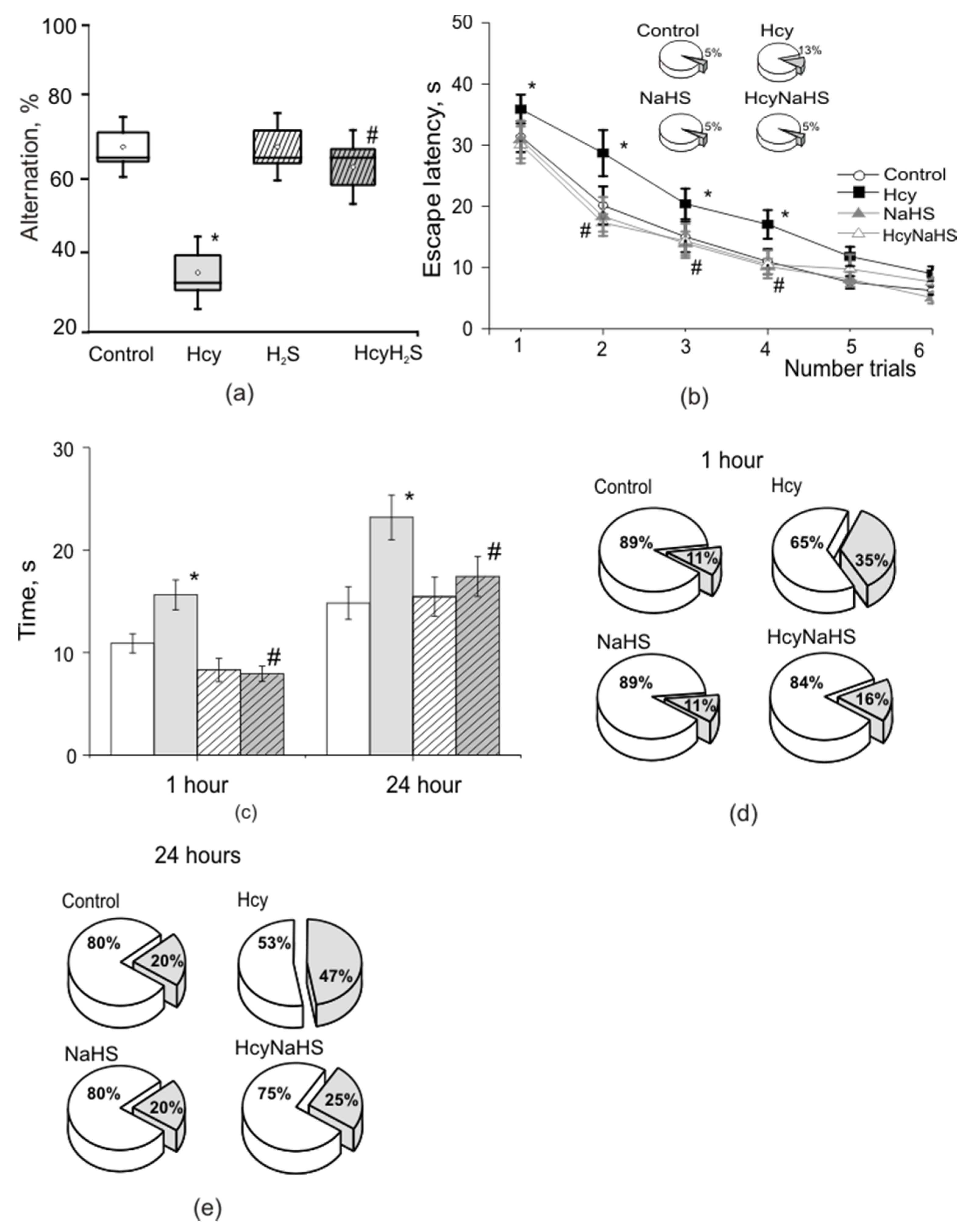
2.2.7. T Maze
2.2.8. Morris Water Maze
2.3. Biochemical Analysis
2.3.1. Measurements of Plasma Homocysteine Level
2.3.2. Assay of H2S Generation
2.3.3. Lipid Peroxidation and the Activity of Glutathione Peroxidases
2.4. Western Blot
2.5. Statistical Analysis
3. Results
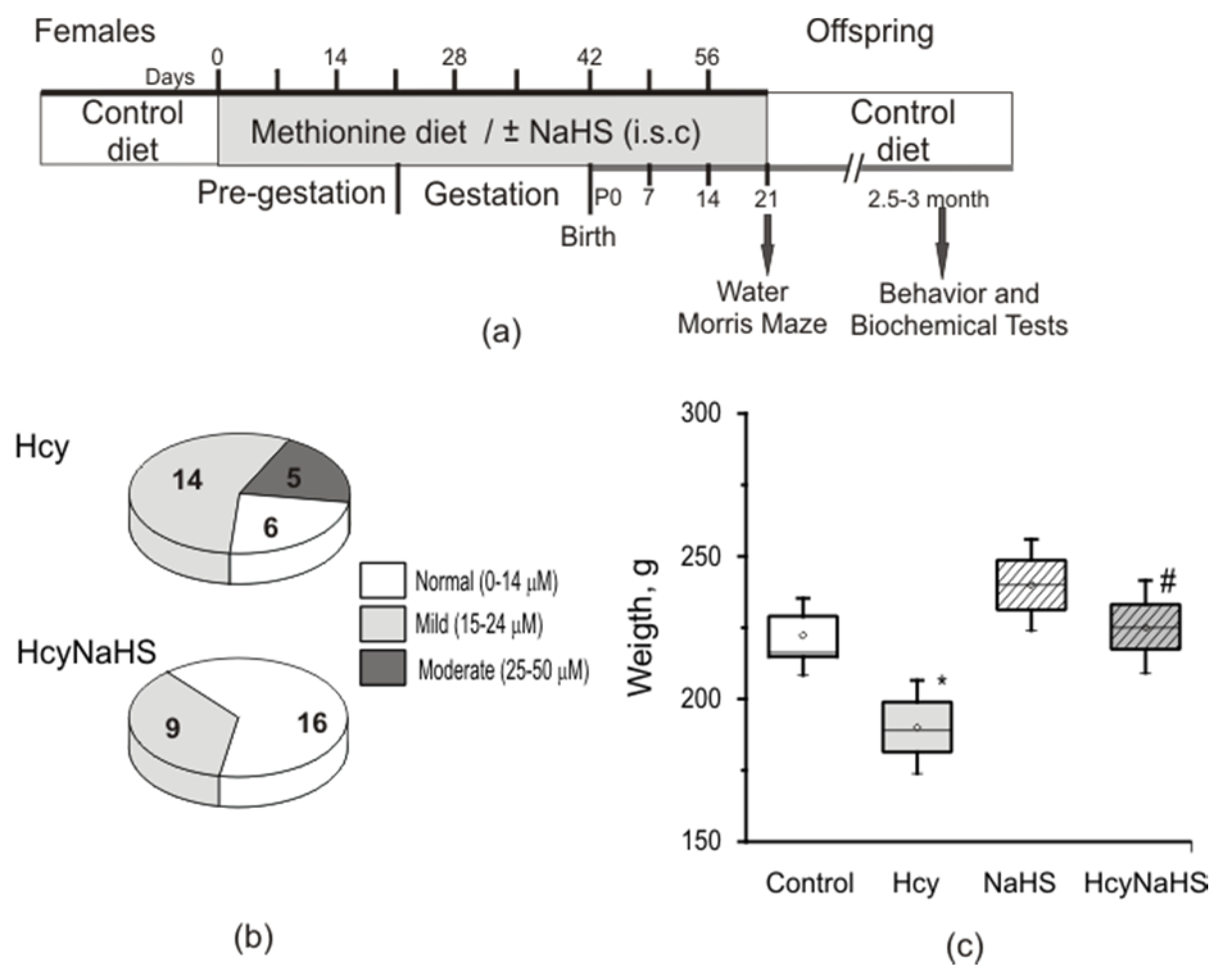
3.1. Maternal hHcy Increases Plasma Hcy Level and Decreases the Weight of the Rat Offspring. Effects of H2S Donor
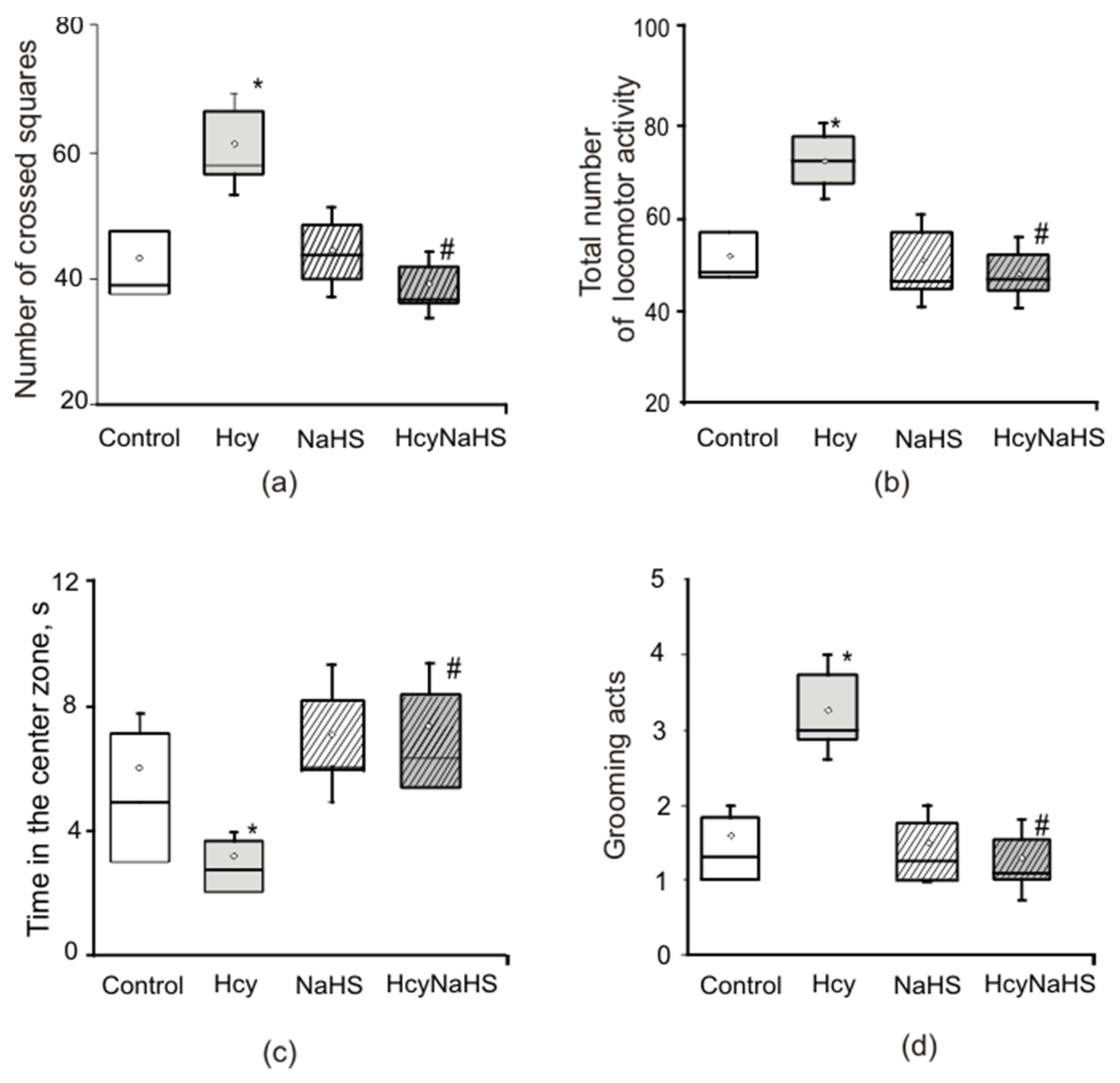
3.2. Effects of Maternal hHcy and NaHS on Behavior in the Open Field Test
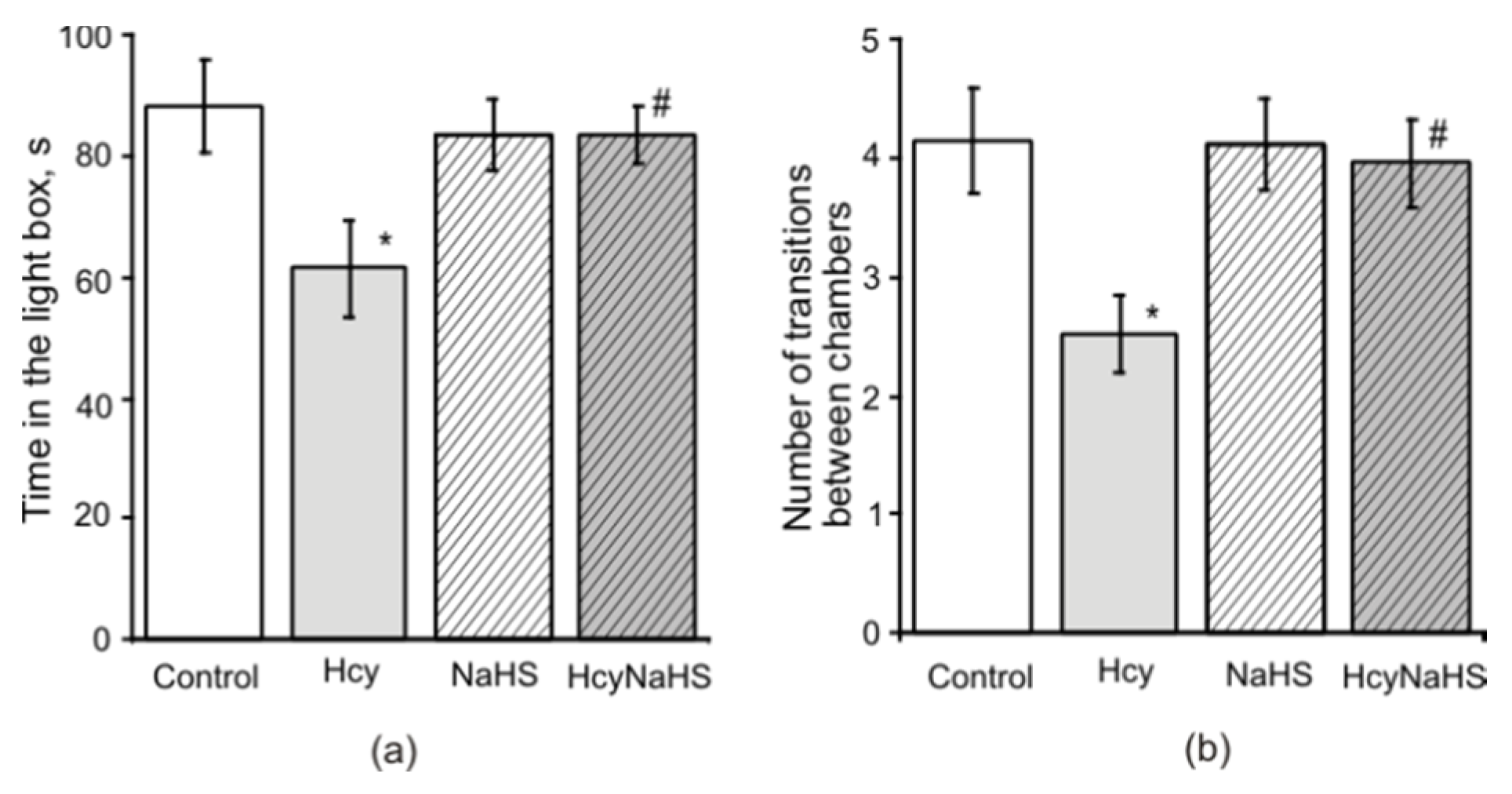
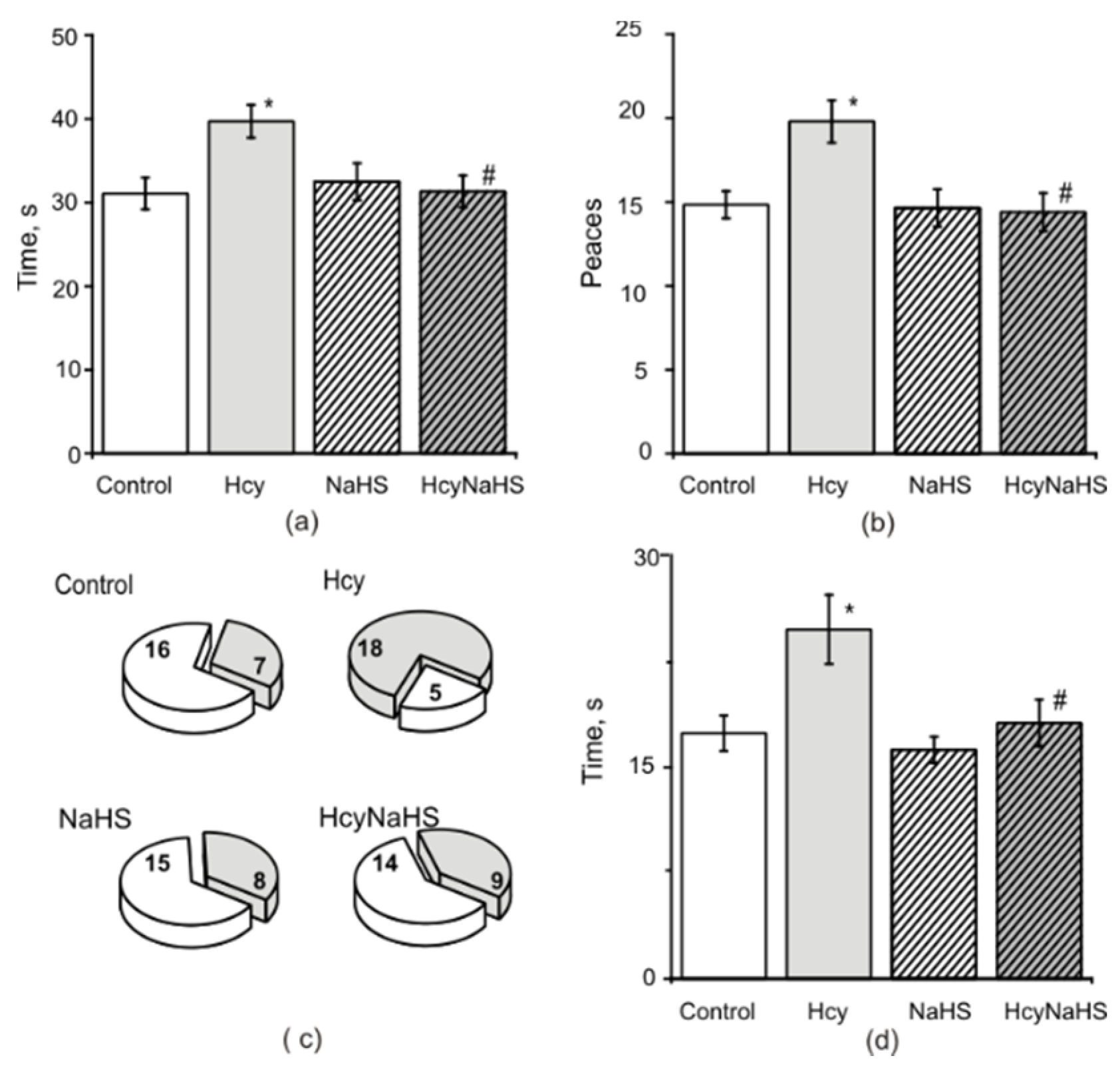
3.3. NaHS Decreases the Anxiety Level Measured in the Light-Dark Box in Rats with Maternal hHcy
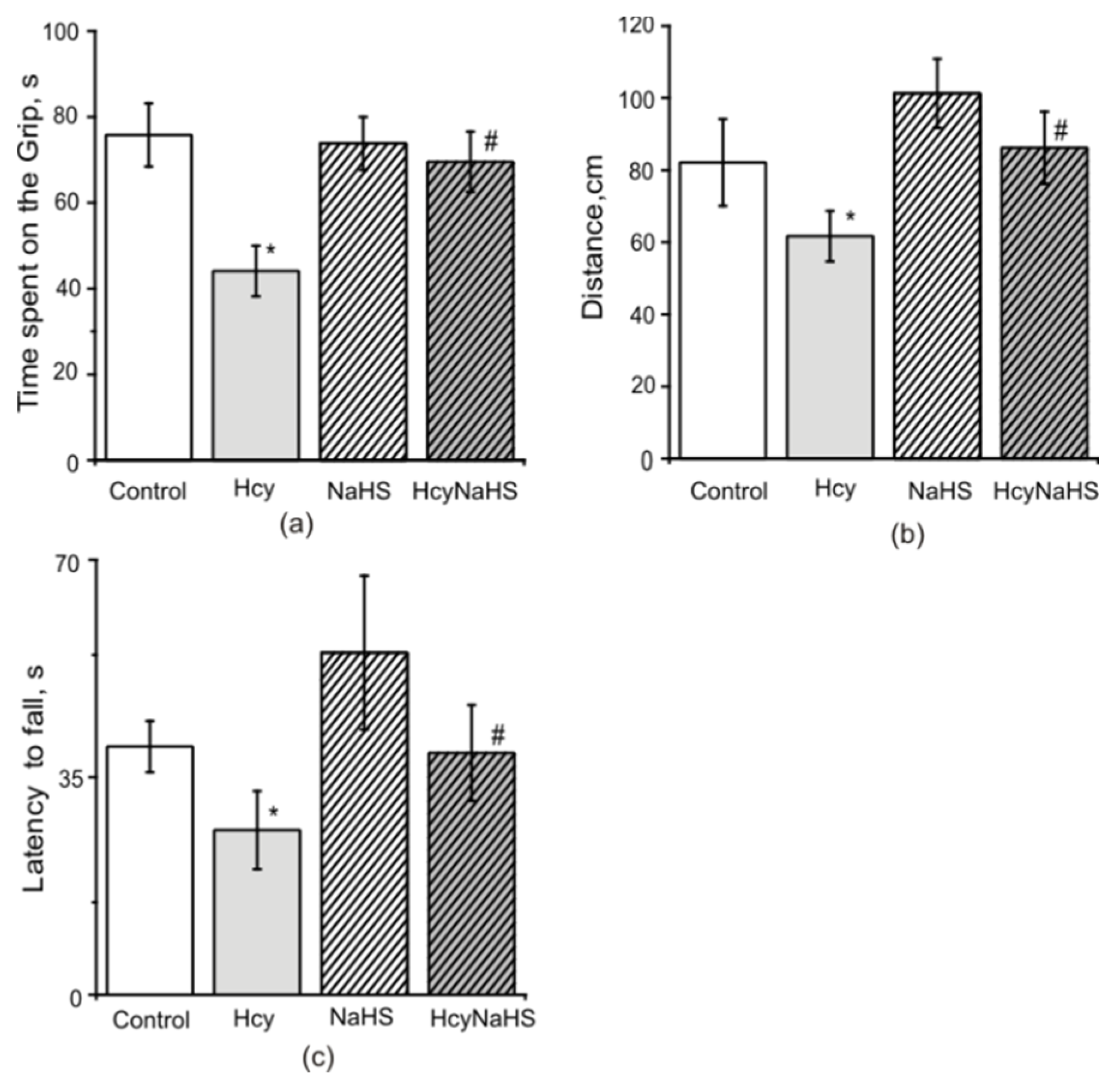
3.4. NaHS Increases Muscle Endurance and Motor Coordination of Rats with Maternal hHcy
3.5. Fine Motor Control and Sensory Motor Asymmetry in Rats with Maternal hHcy. Effects of NaHS
3.6. NaHS Improves Spatial Learning and Memory of Rats with Maternal hHcy
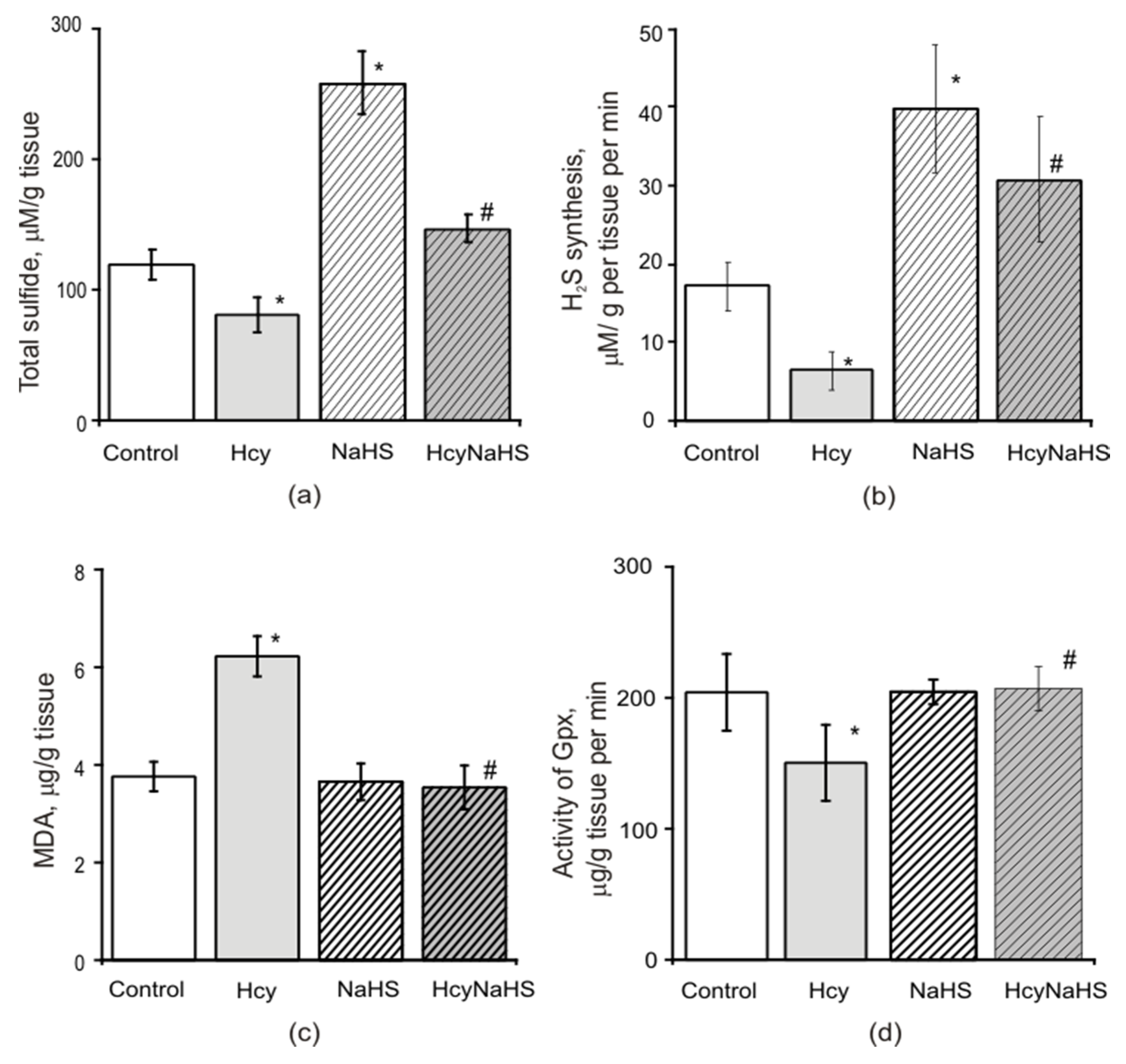
3.7. Effects of Maternal hHcy and NaHS Administration on the Level of Oxidative Stress, H2S Concentration, CBS Activity and Expression in Brain Tissues
3.7.1. H2S Level
3.7.2. Lipid Peroxidation and Activity of Glutathione Peroxidase
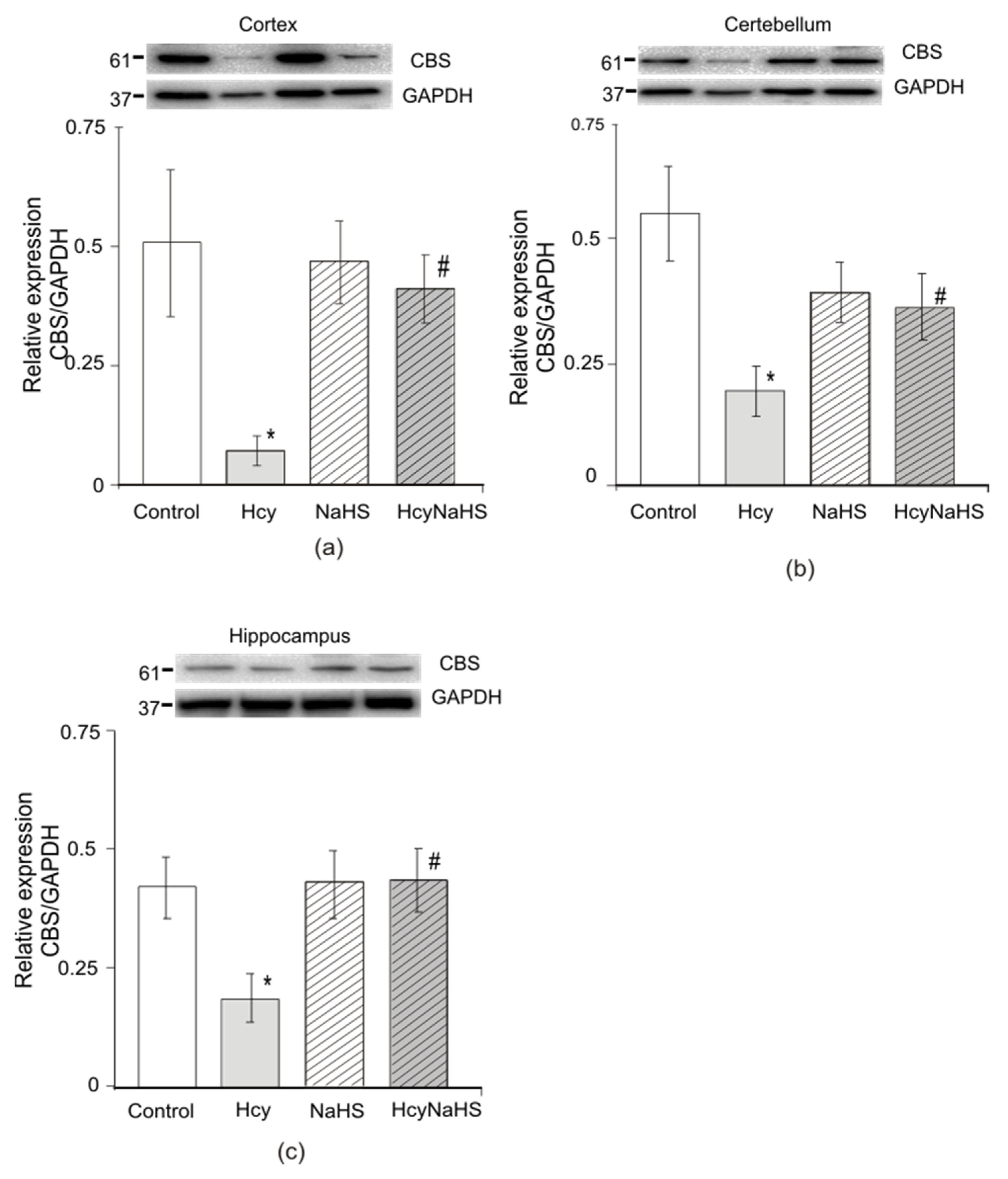
3.7.3. Expression of Cystathionine-Beta Synthase (CBS)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hermann, A.; Sitdikova, G.F.; Weiger, T.M. Modulated by Gaso-Transmitters: BK Channels. In Gasotransmitters: Physiology and Pathophysiology; Hermann, A., Sitdikova, G.F., Weiger, T.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 6, pp. 163–201. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H. Gasotransmitter Hydrogen Sulfide Signaling in Neuronal Health and Disease. Biochem. Pharmacol. 2018, 149, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.B.; Wu, W.N.; Wang, W.; Gu, X.H.; Yu, B.; Wei, B.; Yang, Y.J. Cystathionine-beta-synthase-derived hydrogen sulfide is required for amygdalar long-term potentiation and cued fear memory in rats. Pharmacol. Biochem. Behav. 2017, 155, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Kuksis, M.; Smith, P.M.; Ferguson, A.V. Hydrogen sulfide regulates cardiovascular function by influencing the excitability of subfornical organ neurons. PLoS ONE 2014, 9, e105772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakovlev, A.V.; Kurmasheva, E.D.; Giniatullin, R.; Khalilov, I.; Sitdikova, G.F. Hydrogen sulfide inhibits giant depolarizing potentials and abolishes epileptiform activity of neonatal rat hippocampal slices. Neuroscience 2017, 340, 153–165. [Google Scholar] [CrossRef] [PubMed]
- García-Bereguía, M.A.; Samhan-Arias, A.K.; Martín-Romero, F.J.; Gutiérrez-Merino, C. Hydrogen sulfide raises cytosolic calcium in neurons through activation L-type Ca2+-channels. Antioxid. Redox Signal. 2008, 10, 31–42. [Google Scholar] [CrossRef]
- Okubo, K.; Matsumura, M.; Kawaishi, Y.; Aoki, Y.; Matsunami, M.; Okawa, Y.; Sekiguchi, F.; Kawabata, A. Hydrogen sulfide-induced mechanical hyperalgesia and allodynia require activation of both Cav3.2 and TRPA1 channels in mice. Br. J. Pharmacol. 2012, 166, 1738–1743. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, K.; Mustafina, A.; Yakovlev, A.; Hermann, A.; Giniatullin, R.; Sitdikova, G. Receptor mechanisms mediating the pro-nociceptive action of hydrogen sulfide in rat trigeminal neurons and meningeal afferents. Front. Cell. Neurosci. 2017, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Sandhir, R. Hydrogen sulfide in physiological and pathological mechanisms in brain. CNS Neurol. Disord. Drug Targets 2018, 17, 654–670. [Google Scholar] [CrossRef]
- Yang, Q.; He, G.W. Imbalance of Homocysteine and H2S: Significance, Mechanisms, and Therapeutic Promise in Vascular Injury. Oxidative Med. Cell. Longev. 2019, 22, 7629673. [Google Scholar] [CrossRef] [Green Version]
- Petras, M.; Tatarkova, Z.; Kovalska, M.; Mokra, D.; Dobrota, D.; Lehotsky, J.; Drgova, A. Hyperhomocysteinemia as a risk factor for the neuronal system disorders. J. Physiol. Pharmacol. 2014, 65, 15–23. [Google Scholar] [PubMed]
- Hu, L.F.; Lu, M.; Tiong, C.X.; Dawe, G.S.; Hu, G.; Bian, J.S. Neuroprotective effects of hydrogen sulfide on Parkinson’s disease rat models. Aging Cell 2010, 9, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Qin, J.; Chang, X.; Yang, Z.; Tang, X.; Du, J. Hydrogen sulfide may improve the hippocampal damage induced by recurrent febrile seizures in rats. Biochem. Biophys. Res. Commun. 2005, 327, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Shi, Y.; Cui, M.M.; Li, M.; Wen, X.R.; Zhou, X.Y.; Lou, H.Q.; Wang, Y.L.; Qi, D.S.; Tang, M.; et al. H2S attenuates injury after ischemic stroke by diminishing the assembly of CaMKII with ASK1-MKK3-p38 signaling module. Behav. Brain Res. 2020, 384, 112520. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, R.; Jeong, N.Y. Potential for therapeutic use of hydrogen sulfide in oxidative stress-induced neurodegenerative diseases. Int. J. Med. Sci. 2019, 16, 1386. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Goto, Y.-I.; Kimura, H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxid. Redox Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef]
- Dongo, E.; Beliczai-Marosi, G.; Dybvig, A.S.; Kiss, L. The Mechanism of Action and Role of Hydrogen Sulfide in the Control of Vascular Tone. Nitric Oxide 2018, 81, 75–87. [Google Scholar] [CrossRef]
- Liu, Y.H.; Lu, M.; Hu, L.F.; Wong, P.T.; Webb, G.D.; Bian, J.S. Hydrogen sulfide in the mammalian cardiovascular system. Antioxid. Redox Signal. 2012, 17, 141–185. [Google Scholar] [CrossRef]
- Sitdikova, G.F.; Hermann, A.; Yakovlev, A.V. Neurotoxic and neuroprotective effects of homocysteine and hydrogen sulfide. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2018, 160, 686–704. [Google Scholar]
- Sharma, M.; Tiwari, M.; Tiwari, R.K. Hyperhomocysteinemia: Impact on neurodegenerative diseases. Basic Clin. Pharmacol. Toxicol. 2015, 117, 287–296. [Google Scholar] [CrossRef]
- Yakovleva, O.V.; Ziganshina, A.R.; Gerasimova, E.V.; Arslanova, A.N.; Yarmiev, I.Z.; Zefirov, A.L.; Sitdikova, G.F. The Effect of B Vitamins on the Early Development of Rat Pups with Prenatal Hyperhomocysteinemia. Russ. J. Physiol. 2019, 105, 1247–1261. [Google Scholar] [CrossRef]
- Moretti, R.; Caruso, P. The Controversial Role of Homocysteine in Neurology: From Labs to Clinical Practice. Int. J. Mol. Sci. 2019, 20, 231. [Google Scholar] [CrossRef] [Green Version]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Cervenová, T.; Halašová, E.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Singh, N. Homocysteine excess: Delineating the possible mechanism of neurotoxicity and depression. Fundam. Clin. Pharmacol. 2015, 29, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Gaifullina, A.S.; Lazniewska, J.; Gerasimova, E.V.; Burkhanova, G.F.; Rzhepetskyy, Y.; Tomin, A.; Rivas-Ramirez, P.; Huang, J.; Cmarko, L.; Zamponi, G.W.; et al. A potential role for T-type calcium channels in homocysteinemia-induced peripheral neuropathy. Pain 2019, 160, 2798–2810. [Google Scholar] [CrossRef] [PubMed]
- Ansari, R.; Mahta, A.; Mallack, E.; Luo, J.J. Hyperhomocysteinemia and neurologic disorders: A review. J. Clin. Neurol. 2014, 10, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Yakovleva, O.V.; Ziganshina, A.R.; Dmitrieva, S.A.; Arslanova, A.N.; Yakovlev, A.V.; Minibayeva, F.V.; Khaertdinov, N.N.; Ziyatdinova, G.K.; Giniatullin, R.A.; Sitdikova, G.F. Hydrogen Sulfide Ameliorates Developmental Impairments of Rat Offspring with Prenatal Hyperhomocysteinemia. Oxidative Med. Cell. Longev. 2018, 2018, 2746873. [Google Scholar] [CrossRef] [Green Version]
- Gerasimova, E.; Yakovleva, O.; Burkhanova, G.; Ziyatdinova, G.; Khaertdinov, N.; Sitdikova, G. Effects of Maternal Hyperhomocysteinemia on the Early Physical Development and Neurobehavioral Maturation of Rat Offspring. BioNanoScience 2017, 7, 155–158. [Google Scholar] [CrossRef]
- Makhro, A.V.; Mashkina, A.P.; Solenaya, O.A.; Trunova, O.A.; Kozina, L.S.; Arutyunian, A.V.; Bulygina, E.R. Prenatal hyperhomocysteinemia as a model of oxidative stress of the brain. Bull. Exp. Biol. Med. 2008, 146, 3–35. [Google Scholar] [CrossRef]
- Blaise, S.A.; Nédélec, E.; Schroeder, H.; Alberto, J.M.; Bossenmeyer-Pourié, C.; Guéant, J.L.; Daval, J.L. Gestational vitamin B deficiency leads to homocysteine-associated brain apoptosis and alters neurobehavioral development in rats. Am. J. Pathol. 2007, 170, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Baydas, G.; Koz, S.T.; Tuzcu, M.; Nedzvetsky, V.S.; Etem, E. Effects of maternal hyperhomocysteinemia induced by high methionine diet on the learning and memory performance in offspring. Int. J. Dev. Neurosci. 2007, 25, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Arutjunyan, A.; Kozina, L.; Stvolinskiy, S.; Bulygina, Y.; Mashkina, A.; Khavinson, V. Pinealon protects the rat offspring from prenatal hyperhomocysteinemia. Int. J. Clin. Exp. Med. 2012, 5, 179–185. [Google Scholar] [PubMed]
- Ars, C.L.; Nijs, I.M.; Marroun, H.E.; Muetzel, R.; Schmidt, M.; Steenweg-de Graaff, J.; van der Lugt, A.; Jaddoe, V.W.; Hofman, A.; Steegers, E.A.; et al. Prenatal folate, homocysteine and vitamin B12 levels and child brain volumes, cognitive development and psychological functioning: The Generation R Study. Br. J. Nutr. 2019, 122 (Suppl. 1), S1–S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.H.; Tang, J.P.; Zhang, P.; Li, X.; Wang, C.-Y.; Wei, H.-J.; Yang, X.-F.; Zou, W.; Tang, X.-Q. Disturbance of endogenous hydrogen sulfide generation and endoplasmic reticulum stress in hippocampus are involved in homocysteine-induced defect in learning and memory of rats. Behav. Brain Res. 2014, 262, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.K.; Kyles, P.; Kalani, A.; Tyagi, N. Hydrogen sulfide ameliorates homocysteine-induced Alzheimer’s disease-likepathology, blood-brain barrier disruption, and synaptic disorder. Mol. Neurobiol. 2016, 53, 2451–2467. [Google Scholar] [CrossRef] [PubMed]
- Bełtowski, J.; Wójcicka, G.; Wojtak, A. Effect of experimental hyperhomocysteinemia on plasma lipid profile, insulin sensitivity and paraoxonase 1 in the rat. Adipobiology 2012, 4, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Naik, A.A.; Patro, I.K.; Patro, N. Slow Physical Growth, Delayed Reflex Ontogeny, and Permanent Behavioral as Well as Cognitive Impairments in Rats Following Intra-generational Protein Malnutrition. Front. Neurosci. 2015, 9, 446. [Google Scholar] [CrossRef] [Green Version]
- Weydt, P.; Hong, S.Y.; Kliot, M.; Moller, T. Assessing disease onset and progression in the SOD1 mouse model of ALS. Neuroreport 2003, 14, 1051–1054. [Google Scholar] [CrossRef]
- Barlow, C.; Hirotsune, S.; Paylor, R.; Liyanage, M.; Eckhaus, M.; Collins, F.; Shiloh, Y.; Crawley, J.N.; Ried, T.; Tagle, D.; et al. Atm-deficient mice: A paradigm of ataxia telangiectasia. Cell 1996, 86, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Karl, T.; Pabst, R.; Von HÖrsten, S. Behavioral phenotyping of mice in pharmacological and toxicological research. Exp. Toxic Pathol. 2003, 55, 69–83. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Gorny, B.; Sarna, J. Paw and limb use in skilled and spontaneous reaching after pyramidal tract, red nucleus and combined lesions in the rat: Behavioral and anatomical dissociations. Behav. Brain Res. 1998, 93, 167–183. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Coles, B.L. Varieties of paw and digit movement during spontaneous food handling in rats: Postures, bimanual coordination, preferences, and the effect of forelimb cortex lesions. Behav. Brain Res. 1996, 77, 135–148. [Google Scholar] [CrossRef]
- Allred, R.P.; Adkins, D.A.L.; Woodlee, M.T.; Husbands, L.C.; Maldonado, M.A.; Kane, d.J.R.; Schallert, T.; Jones, T.A. The Vermicelli Handling Test: A Simple Quantitative Measure of Dexterous Forepaw Function in Rats. J. Neurosci. Methods 2008, 170, 229–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouet, V.; Freret, T. Chapter 3: A Master Key to Assess Stroke Consequences Across Species: The Adhesive Removal Test. In Advances in the Preclinical Study of Ischemic Stroke; Maurizio, B., Ed.; Intech Open Limited: London, UK, 2012; pp. 47–68. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M.; Hascoet, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Wenk, G.L. Assessment of spatial memory using the T maze. Curr. Protoc. Neurosci. 2001, 4, 8.5B.1–8.5A.7. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Assessing Spatial Learning and Memory in Rodents. ILAR J. 2014, 55, 310–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vouros, A.; Gehring, T.V.; Szydlowska, K.; Janusz, A.; Tu, Z.; Croucher, M.; Lukasiuk, K.; Konopka, W.; Sandi, C.; Vasilaki, E. A generalised framework for detailed classification of swimming paths inside the Morris Water Maze. Sci. Rep. 2018, 8, 15089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.T.; Lowinsohn, D.; Compton, R.G. The selective electrochemical detection of homocysteine in the presence of glutathione, cysteine and ascorbic acid using carbon electrodes. Analyst 2014, 139, 3755–3762. [Google Scholar] [CrossRef]
- Ziyatdinova, G.; Grigor’eva, L.; Morozov, M.; Gilmutdinov, A.; Budnikov, H. Electrochemical oxidation of sulfur-containing amino acids on an electrode modified with multi-walled carbon nanotubes. Microchim. Acta 2009, 165, 353–359. [Google Scholar] [CrossRef]
- Yusuf, M.; Huat, B.T.K.; Hsu, A.; Whiteman, M.; Bhatia, M.; Moore, P.K. Streptozocin-induced diabetes in the rat is associated with enhanced tissue hydrogen sulphide biosynthesis. Biochem. Biophys. 2005, 333, 1146–1152. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Razygraev, V.; Yushina, A.D.; Titovich, I.A. A method of measuring glutathione peroxidase activity in murine brain: Application in Pharmacological Experiment. Bull. Exper. Biol. Med. 2018, 165, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar]
- Cheng, Z.; Shen, X.; Jiang, X.; Shan, H.; Cimini, M.; Fang, P.; Ji, Y.; Park, J.Y.; Drosatos, K.; Yang, X.; et al. Hyperhomocysteinemia potentiates diabetes-impaired EDHF-induced vascular relaxation: Role of insufficient hydrogen sulfide. Redox Biol. 2018, 16, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Sandhir, R. Neuroprotective Effect of Hydrogen Sulfide in Hyperhomocysteinemia Is Mediated Through Antioxidant Action Involving Nrf2. Neuromol. Med. 2018, 20, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Prathapasinghe, G.A.; Siow, Y.L.; Xu, Z.; O, K. Inhibition of cystathionine-beta-synthase activity during renal ischemiareperfusion: Role of pH and nitric oxide. Am. J. Physiol. Ren. Physiol. 2008, 295, F912–F922. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N. Exploratory behavior models of anxiety in mice. Neurosci. Biobehav. Rev. 1985, 9, 37–44. [Google Scholar] [CrossRef]
- Shcherbitskaya, A.D.; Milyutina, Y.P.; Zaloznyaya, I.V.; Arutjunyan, A.V.; Nalivaeva, N.N.; Zhuravin, I.A. The Effects of Prenatal Hyperhomocysteinemia on the Formation of Memory and the Contents of Biogenic Amines in the Rat Hippocampus. Neurochem. J. 2017, 11, 296–301. [Google Scholar] [CrossRef]
- Kumar, M.; Modi, M.; Sandhir, R. Hydrogen sulfide attenuates homocysteine-induced cognitive deficits and neurochemical alterations by improving endogenous hydrogen sulfide levels. Biofactors 2017, 43, 434–450. [Google Scholar] [CrossRef]
- Makhro, A.V.; Bulygina, E.R.; Boldyrev, A.A. Effects of Homocysteine and Homocystein Acid on Cerebellar Granule Cells. Neurochem. J. 2007, 1, 127–132. [Google Scholar] [CrossRef]
- Sibarov, D.A.; Giniatullin, R.; Antonov, S.M. High sensitivity of cerebellar neurons to homocysteine is determined by expression of GluN2C and GluN2D subunits of NMDA receptors. Biochem. Biophys. Res. Commun. 2018, 506, 648–652. [Google Scholar] [CrossRef]
- Jacqmain, J.; Nudi, T.E.; Fluharty, S.; Smith, J.S. mFC Pre and post-injury environmental enrichment effects functional recovery following medial frontal cortical contusion injury in rats. Behav. Brain Res. 2014, 275, 201–211. [Google Scholar] [CrossRef]
- Vidal-Martinez, G.; Najera, K.; Miranda, J.D.; Gil-Tommee, C.; Yang, B.; Vargas-Medrano, J.; Diaz-Pacheco, V.; Perez, R.G. FTY720 Improves Behavior, Increases Brain Derived Neurotrophic Factor Levels and Reduces α-Synuclein Pathology in Parkinsonian GM2 +/− Mice. Neuroscience 2019, 411, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, N.; Borah, A. Oxidative stress and mitochondrial dysfunction are the underlying events of dopaminergic neurodegeneration in homocysteine rat model of Parkinson’s disease. Neurochem. Int. 2016, 101, 48–55. [Google Scholar] [CrossRef]
- Avisek, M.; Mahavir, S.; Akash, K.; George, S.C. Tyagi Restoration of skeletal muscle homeostasis by hydrogen sulfide during hyperhomocysteinemia-mediated oxidative/ER stress condition. Can. J. Physiol. Pharmacol. 2019, 97, 441–456. [Google Scholar] [CrossRef]
- Khuzakhmetova, V.; Yakovleva, O.; Dmitrieva, S.; Khaertdinov, N.; Ziyatdinova, G.; Giniatullin, R.; Bukharaeva, E.; Yakovlev, A.; Sitdikova, G. Prenatal hyperhomocysteinemia induces oxidative stress and accelerates ‘aging’ of mammalian neuromuscular synapses. Int. J. Dev. Neurosci. 2019, 75, 1–12. [Google Scholar] [CrossRef]
- Bukharaeva, E.; Shakirzyanova, A.; Khuzakhmetova, V.; Sitdikova, G.; Giniatullin, R. Homocysteine aggravates ROS-induced depression of transmitter release from motor nerve terminals: Potential mechanism of peripheral impairment in motor neuron diseases associated with hyperhomocysteinemia. Front. Cell. Neurosci. 2015, 9, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoccolella, S.; Tortorella, C.; Iaffaldano, P.; Direnzo, V.; D’Onghia, M.; Paolicelli, D.; Livrea, P.; Trojano, M. Elevated plasma homocysteine levels in patients with multiple sclerosis are associated with male gender. J. Neurol. 2012, 259, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Luiz, S.E.; Bavaresco, C.S.; Netto, C.A.; Wyse, A.T.S. Chronic hyperhomocysteinemia provokes a memory deficit in rats in the Morris water maze task. Behav. Brain Res. 2004, 153, 377–381. [Google Scholar] [CrossRef]
- Matte’, C.; Mackedanz, V.; Stefanello, F.M.; Scherer, E.B.; Andreazza, A.C.; Zanotto, C.; Moro, A.M.; Garcia, S.C.; Gonçalves, C.A.; Erdtmann, B.; et al. Chronic hyperhomocysteinemia alters antioxidant defenses and increases DNA damage in brain and blood of rats: Protective effect of folic acid. Neurochem. Int. 2009, 54, 7–13. [Google Scholar] [CrossRef]
- Jadavji, N.M.; Deng, L.; Malysheva, O.; Caudill, M.A.; Rozen, R. MTHFR Deficiency or Reduced Intake of Folate or Choline in Pregnant Mice Results in Impaired Short-Term Memory and Increased Apoptosis in the Hippocampus of Wild-Type Offspring. Neuroscience 2015, 300, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Mahajan, A.; Sapehia, D.; Kaur, J.; Sandhir, R. Effects of altered maternal folate and vitamin B12 on neurobehavioral outcomes in F1 male mice. Brain Res. Bull. 2019, 153, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R. The neurobiological basis of spontaneous alternation. Neurosci. Biobehav. 2002, 26, 91–104. [Google Scholar] [CrossRef]
- Wyse, A.T.; Zugno, A.I.; Streck, E.L.; Matte’, C.; Calcagnotto, T.; Wannmacher, C.M.; Wajner, M. Inhibition of Na, K-ATPase activity in hippocampus of rats to a cute administration of homocysteine is prevented by vitamin E and C treatment. Neurochem. Res. 2002, 27, 1685–1689. [Google Scholar] [CrossRef]
- Machado, F.R.; Ferreira, A.G.K.; da Cunha, A.A.; Tagliari, B.; Mussulini, B.H.M.; Wofchuk, S.; Wise, A.T.S. Homocysteine alters glutamate uptake and Na+, K+-ATPase activity and oxidative status in rats hippocampus: Protection by vitamin C. Metab. Brain Dis. 2011, 26, 61–67. [Google Scholar] [CrossRef]
- Zhuravin, I.A.; Dubrovskaya, N.M.; Vasilev, D.S.; Postnikova, T.Y.; Zaitsev, A.V. Prenatal hypoxia produces memory deficits associated with impairment of long-term synaptic plasticity in young rats. Neurobiol. Learn. Mem. 2019, 164, 107066. [Google Scholar] [CrossRef]
- Koz, T.; Gouwy, N.T.; Demir, N.; Nedzvetsky, V.S.; Etem, E.; Baydas, G. Effects of maternal hyperhomocysteinemia induced by methionine intake on oxidative stress and apoptosis in pup rat brain. Int. J. Dev. Neurosci. 2010, 28, 325–329. [Google Scholar] [CrossRef]
- Pustygina, A.V.; Milyutina, Y.P.; Zaloznyaya, I.V.; Arutyunyan, A.V. Indices of oxidative stress in the brain of newborn rat subjected to prenatal hyperhomocysteinemia. Neurochem. J. 2015, 9, 60–65. [Google Scholar] [CrossRef]
- Yakovlev, A.V.; Kurmashova, E.; Zakharov, A.; Sitdikova, G.F. Network-Driven Activity and Neuronal Excitability in Hippocampus of Neonatal Rats with Prenatal Hyperhomocysteinemia. BioNanoScience 2018, 8, 304–309. [Google Scholar] [CrossRef]
- Li, J.J.; Li, Q.; Du, H.P.; Wang, Y.-L.; You, S.-J.; Wang, F.; Xu, X.-S.; Cheng, J.; Cao, Y.-J.; Liu, C.-F.; et al. Homocysteine triggers inflammatory responses in macrophages through inhibiting cse-h2s signaling via DNA hypermethylation of CSE promoter. Int. J. Mol. Sci. 2015, 16, 12560–12577. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, Z.; Xu, G. Notable epigenetic role of hyperhomocysteinemia in atherogenesis. Lipids Health Dis. 2014, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, J.; Tyagi, S.C.; Tyagi, N. Hyperhomocysteinemia induced endothelial progenitor cells dysfunction through hyper-methylation of CBS promoter. Biochem. Biophys. Res. Commun. 2019, 510, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.K.; Kalani, A.; Givvimani, S.; Sathnur, P.B.; Tyagi, S.C.; Tyagi, N. Hydrogen sulfide attenuates neurodegeneration and neurovascular dysfunction induced by intracerebral-administered homocysteine in mice. Neuroscience 2013, 252, 302–319. [Google Scholar] [CrossRef] [Green Version]
- Nalivaeva, N.N.; Turner, A.J.; Zhuravin, I.A. Role of Prenatal Hypoxia in Brain Development, Cognitive Functions, and Neurodegeneration. Front. Neurosci. 2018, 12, 825. [Google Scholar] [CrossRef] [Green Version]
- Babenko, O.; Kovalchuk, I.; Metz, G.A. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci. Biobehav. Rev. 2015, 48, 70–91. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; D’Angelo, G.; Manti, S.; Arrigo, T.; Barberi, I.; Reiter, R.J.; Gitto, E. Oxidative stress-mediated aging during the fetal and perinatal periods. Oxidative Med. Cell. Longev. 2014, 2014, 358375. [Google Scholar] [CrossRef] [Green Version]
- Enokido, Y.; Suzuki, E.; Iwasawa, K.; Namekata, K.; Okazawa, H.; Kimura, H. Cystathionine beta-synthase, a key enzyme for homocysteine metabolism, is preferentially expressed in the radial glia/astrocyte lineage of developing mouse CNS. FASEB J. 2005, 19, 1854–1856. [Google Scholar] [CrossRef]
- Bruintjes, J.J.; Henning, R.H.; Douwenga, W.; van der Zee, E.A. Hippocampal cystathionine beta synthase in young and aged mice. Neurosci. Lett. 2014, 563, 135–139. [Google Scholar] [CrossRef]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA excitatory/inhibitory shift in brain maturation and neurological disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef]
- Yakovlev, A.V.; Kurmasheva, E.D.; Ishchenko, Y.; Giniatullin, R.; Sitdikova, G.F. Age-dependent, subunit specific action of hydrogen sulfide on GluN1/2A and GluN1/2B NMDA receptors. Front. Cell. Neurosci. 2017, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.S.; Mishra, P.K. H2S and homocysteine control a novel feedback regulation of cystathionine beta synthase and cystathionine gamma lyase in cardiomyocytes. Sci. Rep. 2017, 7, 3639. [Google Scholar] [CrossRef] [PubMed]
- Carballal, S.; Trujillo, M.; Cuevasanta, E.; Bartesaghi, S.; Moller, M.N.; Folkes, L.K. Reactivity of hydrogen sulfide with peroxynitrite and other oxidants of biological interest. Free Radic. Biol. Med. 2010, 1, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Ray, R.S.; Sandhir, R. Hydrogen sulfide attenuates homocysteine-induced neurotoxicity by preventing mitochondrial dysfunctions and oxidative damage: In vitro and in vivo studies. Neurochem. Int. 2018, 120, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Calvert, J.W.; Jha, S.; Gundewar, S.; Elrod, J.W.; Ramachandran, A.; Pattillo, C.B.; Kevil, C.G.; Lefer, D.J. Hydrogen sulfide mediates cardioprotection through Nrf2 signaling. Circ. Res. 2009, 105, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.-Z.; Liu, Y.; Bian, J.-S. Hydrogen sulfide and cellular redox homeostasis. Oxidative Med. Cell. Longev. 2016, 2016, 6043038. [Google Scholar] [CrossRef] [Green Version]
- Cindrova-Davies, T.; Herrera, E.A.; Niu, Y.; Kingdom, J.; Giussani, D.A.; Burton, G.J. Reduced cystathionine γ -lyase and increased miR-21 expression are associated with increased vascular resistance in growth-restricted pregnancies: Hydrogen sulfide as a placental vasodilator. Am. J. Pathol. 2013, 182, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Kingdom, J.; Burton, G.J.; Cindrova-Davies, T. Placental stem villus arterial remodeling associated with reduced hydrogen sulfide synthesis contributes to human fetal growth restriction. Am. J. Pathol. 2017, 187, 908–920. [Google Scholar] [CrossRef] [Green Version]
- Damian, D.G.; Hurt, K.J. Gasotransmitters in pregnancy: From conception to uterine involution. Biol. Reprod. 2019, 101, 4–25. [Google Scholar] [CrossRef]
- Hu, M.; Zou, W.; Wang, C.Y.; Chen, X.; Tan, H.Y.; Zeng, H.Y.; Zhang, P.; Gu, H.F.; Tang, X.Q. Hydrogen Sulfide Protects against Chronic Unpredictable Mild Stress-Induced Oxidative Stress in Hippocampus by Upregulation of BDNF-TrkB Pathway. Oxidative Med. Cell. Longev. 2016, 2016, 2153745. [Google Scholar] [CrossRef]
- He, J.; Guo, R.; Qiu, P.; Su, X.; Yan, G.; Feng, J. Exogenous hydrogen sulfide eliminates spatial memory retrieval impairment and hippocampal CA1 LTD enhancement caused by acute stress via promoting glutamate uptake. Neuroscience 2017, 350, 110–123. [Google Scholar] [CrossRef]
- Yin, W.L.; Yin, W.G.; Huang, B.S.; Wu, L.X. Neuroprotective effects of lentivirus-mediated cystathionine-beta-synthase overexpression against 6-OHDA-induced Parkinson’s disease rats. Neurosci. Lett. 2017, 657, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.Q.; Wang, Y.L.; Yuan, B.S.; Yuan, X.; Hou, X.O.; Bian, J.S.; Liu, C.F.; Hu, L.F. Impaired CBS-H2S signaling axis contributes to MPTP-induced neurodegeneration in a mouse model of Parkinson’s disease. Brain Behav. Immun. 2018, 67, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Gerasimova, E.; Lebedeva, J.; Yakovlev, A.; Zefirov, A.; Giniatullin, R.; Sitdikova, G. Mechanisms of hydrogen sulfide (H2S) action on synaptic transmission at the mouse neuromuscular junction. Neuroscience 2015, 303, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Vandini, E.; Ottani, A.; Zaffe, D.; Calevro, A.; Canalini, F.; Cavallini, G.M.; Rossi, R.; Guarini, S.; Giuliani, D. Mechanisms of Hydrogen Sulfide against the Progression of Severe Alzheimer’s Disease in Transgenic Mice at Different Ages. Pharmacology 2019, 103, 50–60. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yakovleva, O.; Bogatova, K.; Mukhtarova, R.; Yakovlev, A.; Shakhmatova, V.; Gerasimova, E.; Ziyatdinova, G.; Hermann, A.; Sitdikova, G. Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress. Biomolecules 2020, 10, 995. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10070995
Yakovleva O, Bogatova K, Mukhtarova R, Yakovlev A, Shakhmatova V, Gerasimova E, Ziyatdinova G, Hermann A, Sitdikova G. Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress. Biomolecules. 2020; 10(7):995. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10070995
Chicago/Turabian StyleYakovleva, Olga, Ksenia Bogatova, Renata Mukhtarova, Aleksey Yakovlev, Viktoria Shakhmatova, Elena Gerasimova, Guzel Ziyatdinova, Anton Hermann, and Guzel Sitdikova. 2020. "Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress" Biomolecules 10, no. 7: 995. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10070995