
Identification, Synthesis, Conformation and Activity of an Insulin-like Peptide from a Sea Anemone

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insulin-like Peptide Data and Bioinformatic Analysis
2.2. ILP Synthesis
2.3. Spectroscopic Studies
2.3.1. Circular Dichroism Spectroscopy
2.3.2. Nuclear Magnetic Resonance (NMR) Spectroscopy
2.4. Functional Studies
2.4.1. Binding Assays
2.4.2. Ion Channel Assays
3. Results
3.1. ILP Sequence Similarity
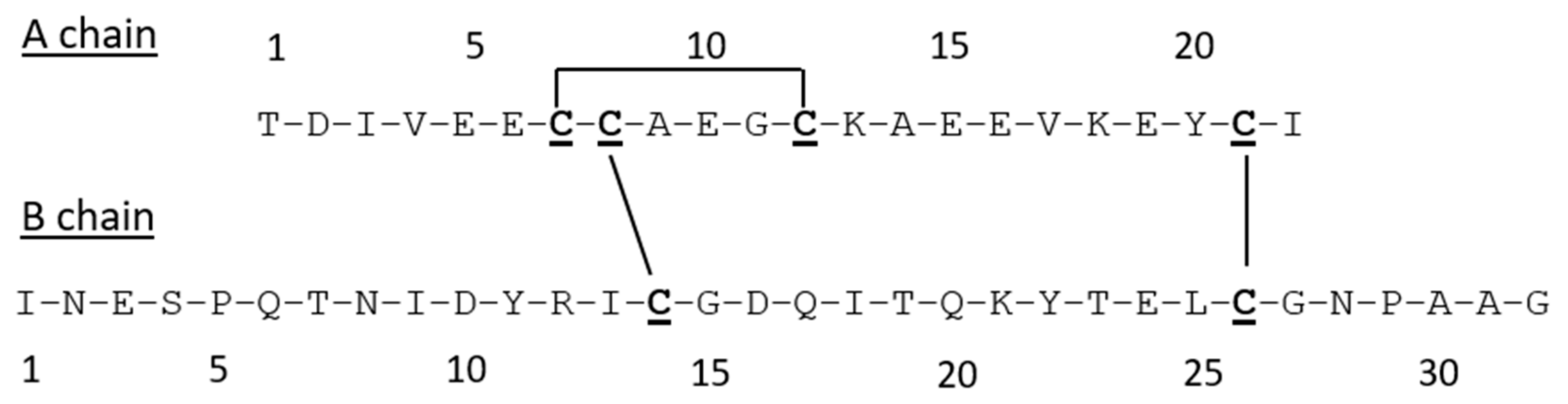
3.2. Synthesis and Characterization
3.3. Spectroscopic Studies
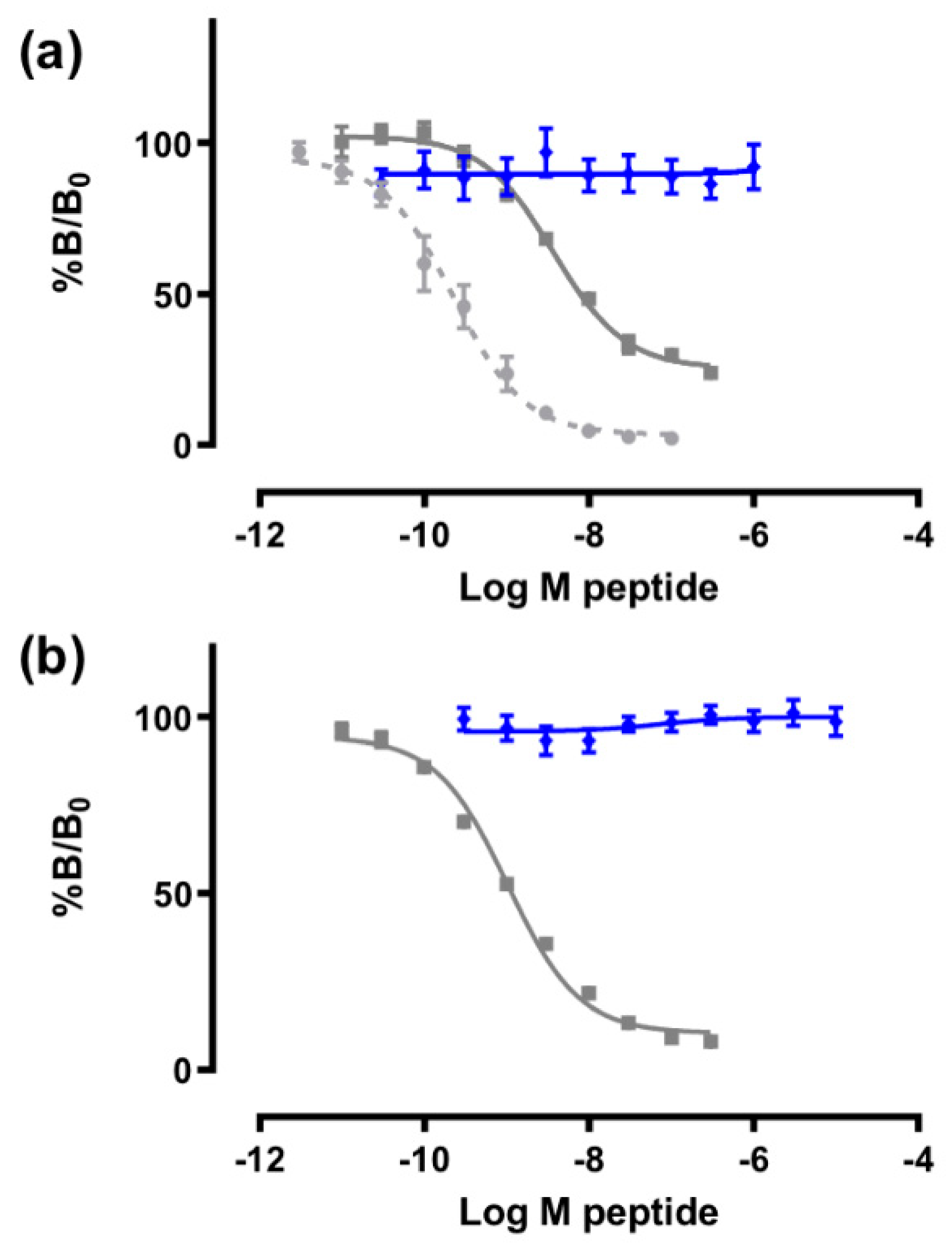
3.4. Receptor Binding Assays
3.5. Potassium, Sodium, and Calcium Channel Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018, 217, 2273–2289. [Google Scholar] [CrossRef] [Green Version]
- Ebberink, R.H.M.; Smit, A.B.; Van Minnen, J. The insulin family: Evolution of structure and function in vertebrates and invertebrates. Biol. Bull. 1989, 177, 176–182. [Google Scholar] [CrossRef]
- Steiner, D.F.; Chan, S.J.; Welsh, J.M.; Kwok, S.C. Structure and evolution of the insulin gene. Annu. Rev. Genet. 1985, 19, 463–484. [Google Scholar] [CrossRef]
- Grönke, S.; Clarke, D.-F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Brown, M.R. Signaling and function of insulin-like peptides in insects. Annu. Rev. Entomol. 2005, 51, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Chowański, S.; Walkowiak-Nowicka, K.; Winkiel, M.; Marciniak, P.; Urbański, A.; Pacholska-Bogalska, J. Insulin-like peptides and cross-talk with other factors in the regulation of insect metabolism. Front. Physiol. 2021, 12, 701203. [Google Scholar] [CrossRef]
- Semaniuk, U.; Piskovatska, V.; Strilbytska, O.; Strutynska, T.; Burdyliuk, N.; Vaiserman, A.; Bubalo, V.; Storey, K.B.; Lushchak, O. Drosophila insulin-like peptides: From expression to functions—A review. Entomol. Exp. Appl. 2021, 169, 195–208. [Google Scholar] [CrossRef]
- Iwami, M.; Kawakami, A.; Ishizaki, H.; Takahashi, S.Y.; Adachi, T.; Suzuki, Y.; Nagasawa, H.; Suzuki, A. Cloning of a gene encoding bombyxin, an insulin-like brain secretory peptide of the silkmoth Bombyx mori with prothoracicotropic activity. Dev. Growth Differ. 1989, 31, 31–37. [Google Scholar] [CrossRef]
- Zheng, S.; Chiu, H.; Boudreau, J.; Papanicolaou, T.; Bendena, W.; Chin-Sang, I. A functional study of all 40 Caenorhabditis elegans insulin-like peptides. J. Biol. Chem. 2018, 293, 16912–16922. [Google Scholar] [CrossRef] [Green Version]
- Floyd, P.D.; Li, L.; Rubakhin, S.S.; Sweedler, J.V.; Horn, C.C.; Kupfermann, I.; Alexeeva, V.Y.; Ellis, T.A.; Dembrow, N.C.; Weiss, K.R.; et al. Insulin prohormone processing, distribution, and relation to metabolism in Aplysia californica. J. Neurosci. 1999, 19, 7732–7741. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.-S.; Choi, Y.H. Role of the insulin-like growth factor system in gonad sexual maturation in Pacific oyster Crassostrea gigas. Fish. Aquat. Sci. 2020, 23, 3. [Google Scholar] [CrossRef]
- Perillo, M.; Arnone, M.I. Characterization of insulin-like peptides (ILPs) in the sea urchin Strongylocentrotus purpuratus: Insights on the evolution of the insulin family. Gen. Comp. Endocrinol. 2014, 205, 68–79. [Google Scholar] [CrossRef]
- Chandler, J.C.; Aizen, J.; Elizur, A.; Hollander-Cohen, L.; Battaglene, S.C.; Ventura, T. Discovery of a novel insulin-like peptide and insulin binding proteins in the Eastern rock lobster Sagmariasus verreauxi. Gen. Comp. Endocrinol. 2015, 215, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Safavi-Hemami, H. Insulin as a weapon. Toxicon 2016, 123, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Menting, J.G.; Disotuar, M.M.; Smith, N.A.; Delaine, C.A.; Ghabash, G.; Agrawal, R.; Wang, X.; He, X.; Fisher, S.J.; et al. A structurally minimized yet fully active insulin based on cone-snail venom insulin principles. Nat. Struct. Mol. Biol. 2020, 27, 615–624. [Google Scholar] [CrossRef]
- Lauritano, C.; Ianora, A. Marine organisms with anti-diabetes properties. Mar. Drugs 2016, 14, 220. [Google Scholar] [CrossRef]
- Pascual, I.; Lopéz, A.; Gómez, H.; Chappé, M.; Saroyán, A.; González, Y.; Cisneros, M.; Charli, J.L.; de los Angeles Cháveza, M. Screening of inhibitors of porcine dipeptidyl peptidase IV activity in aqueous extracts from marine organisms. Enzyme Microb. Technol. 2007, 40, 414–419. [Google Scholar] [CrossRef]
- Antonova, Y.; Arik, A.J.; Moore, W.; Riehle, M.A.; Brown, M.R. Insulin-Like Peptides: Structure, signaling, and function. In Insect Endocrinology; Academic Press: San Diego, CA, USA, 2012; pp. 63–92. ISBN 978-0-12-384749-2. [Google Scholar]
- Nässel, D.R.; Vanden Broeck, J. Insulin/IGF signaling in Drosophila and other insects: Factors that regulate production, release and post-release action of the insulin-like peptides. Cell. Mol. Life Sci. 2016, 73, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Altindis, E.; Cai, W.; Sakaguchi, M.; Zhang, F.; GuoXiao, W.; Liu, F.; De Meyts, P.; Gelfanov, V.; Pan, H.; DiMarchi, R.; et al. Viral insulin-like peptides activate human insulin and IGF-1 receptor signaling: A paradigm shift for host–microbe interactions. Proc. Natl. Acad. Sci. USA 2018, 115, 2461–2466. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, Y.; Kawano, T. The C. elegans insulin-like peptides (ILPs). AIMS Biophys. 2018, 5, 217–230. [Google Scholar] [CrossRef]
- Duret, L.; Guex, N.; Peitsch, M.C.; Bairoch, A. New insulin-like proteins with atypical disulfide bond pattern characterized in Caenorhabditis elegans by comparative sequence analysis and homology modeling. Genome Res. 1998, 8, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmetz, P.R.H.; Aman, A.; Kraus, J.E.M.; Technau, U. Gut-like ectodermal tissue in a sea anemone challenges germ layer homology. Nat. Ecol. Evol. 2017, 1, 1535–1542. [Google Scholar] [CrossRef]
- Anctil, M. Chemical transmission in the sea anemone Nematostella vectensis: A genomic perspective. Comp. Biochem. Physiol. Part D Genom. Proteom. 2009, 4, 268–289. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.L.; Tonkin-Hill, G.Q.; Morales, R.A.V.; Purcell, A.W.; Papenfuss, A.T.; Norton, R.S. Tentacle transcriptomes of the speckled anemone (Actiniaria: Actiniidae: Oulactis sp.): Venom-related components and their domain structure. Mar. Biotechnol. 2020, 22, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis tool web services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Steiner, D.F.; Park, S.-Y.; Støy, J.; Philipson, L.H.; Bell, G.I. A brief perspective on insulin production. Diabetes Obes. Metab. 2009, 11, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Nicol, D.S.H.W.; Smith, L.F. Amino-acid sequence of human insulin. Nature 1960, 187, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Okamoto, R.; Karas, J.A.; Praveen, P.; Liu, M.; Forbes, B.E.; Wade, J.D.; Kajihara, Y. Total chemical synthesis of a nonfibrillating human glycoinsulin. J. Am. Chem. Soc. 2020, 142, 1164–1169. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Pietrzkowski, Z.; Lammers, R.; Carpenter, G.; Soderquist, A.M.; Limardo, M.; Phillips, P.D.; Ullrich, A.; Baserga, R. Constitutive expression of insulin-like growth factor 1 and insulin-like growth factor 1 receptor abrogates all requirements for exogenous growth factors. Cell Growth Differ. 1992, 3, 199–205. [Google Scholar] [PubMed]
- Denley, A.; Bonython, E.R.; Booker, G.W.; Cosgrove, L.J.; Forbes, B.E.; Ward, C.W.; Wallace, J.C. Structural determinants for high-affinity binding of insulin-like growth factor II to insulin receptor (IR)-A, the exon 11 minus isoform of the IR. Mol. Endocrinol. 2004, 18, 2502–2512. [Google Scholar] [CrossRef] [Green Version]
- Soos, M.A.; Field, C.E.; Lammers, R.; Ullrich, A.; Zhang, B.; Roth, R.A.; Andersen, A.S.; Kjeldsen, T.; Siddle, K. A panel of monoclonal antibodies for the type I insulin-like growth factor receptor. Epitope mapping, effects on ligand binding, and biological activity. J. Biol. Chem. 1992, 267, 12955–12963. [Google Scholar] [CrossRef]
- Soos, M.A.; Siddle, K. Immunological relationships between receptors for insulin and insulin-like growth factor I. Evidence for structural heterogeneity of insulin-like growth factor I receptors involving hybrids with insulin receptors. Biochem. J. 1989, 263, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro-Junior, E.L.; Boldrini-França, J.; Takeda, A.A.S.; Costa, T.R.; Peigneur, S.; Cardoso, I.A.; de Oliveira, I.S.; Sampaio, S.V.; de Mattos Fontes, M.R.; Tytgat, J.; et al. Towards toxin PEGylation: The example of rCollinein-1, a snake venom thrombin-like enzyme, as a PEGylated biopharmaceutical prototype. Int. J. Biol. Macromol. 2021, 190, 564–573. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Surm, J.M.; Stewart, Z.K.; Papanicolaou, A.; Pavasovic, A.; Prentis, P.J. The draft genome of Actinia tenebrosa reveals insights into toxin evolution. Ecol. Evol. 2019, 9, 11314–11328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The genome of Aiptasia, a sea anemone model for coral symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef] [Green Version]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral Eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Ying, H.; Hayward, D.C.; Cooke, I.; Wang, W.; Moya, A.; Siemering, K.R.; Sprungala, S.; Ball, E.E.; Forêt, S.; Miller, D.J. The whole-genome sequence of the coral Acropora millepora. Genome Biol. Evol. 2019, 11, 1374–1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chiou, C.-Y.; Dai, C.-F.; Chen, C.A. Unique mitogenomic features in the scleractinian family Pocilloporidae (Scleractinia: Astrocoeniina). Mar. Biotechnol. 2008, 10, 538–553. [Google Scholar] [CrossRef]
- Flot, J.-F.; Tillier, S. The mitochondrial genome of Pocillopora (Cnidaria: Scleractinia) contains two variable regions: The putative D-loop and a novel ORF of unknown function. Gene 2007, 401, 80–87. [Google Scholar] [CrossRef]
- WormBase Release: W276. Available online: http://www.wormbase.org (accessed on 1 June 2020).
- Kass-Simon, G.; Scappaticci, A.A., Jr. The behavioral and developmental physiology of nematocysts. Can. J. Zool. 2002, 80, 1772–1794. [Google Scholar] [CrossRef]
- Shick, J.M. A Functional Biology of Sea Anemones; Chapman & Hall: London, UK, 1991. [Google Scholar]
- Ashwood, L.M.; Mitchell, M.L.; Madio, B.; Hurwood, D.A.; King, G.F.; Undheim, E.A.B.; Norton, R.S.; Prentis, P.J. Tentacle morphological variation coincides with differential expression of toxins in sea snemones. Toxins 2021, 13, 452. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; O’Brien-Simpson, N.M.; Hossain, M.A.; Wade, J.D. The 9-Fluorenylmethoxycarbonyl (Fmoc) group in chemical peptide synthesis—Its past, present, and future. Aust. J. Chem. 2020, 73, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Wade, J.D. Novel methods for the chemical synthesis of insulin superfamily peptides and of analogues containing disulfide isosteres. Acc. Chem. Res. 2017, 50, 2116–2127. [Google Scholar] [CrossRef]
- Lin, F.; Hossain, M.A.; Post, S.; Karashchuk, G.; Tatar, M.; De Meyts, P.; Wade, J.D. Total solid-phase synthesis of biologically active Drosophila insulin-like peptide 2 (DILP2). Aust. J. Chem. 2017, 70, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Norton, R.S. Peptide toxin structure and function by NMR. In Modern Magnetic Resonance; Webb, G.A., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 2081–2097. ISBN 9783319283883. [Google Scholar]
- Tudor, J.E.; Pennington, M.W.; Norton, R.S. Ionisation behaviour and solution properties of the potassium-channel blocker ShK toxin. Eur. J. Biochem. 1998, 251, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Menting, J.G.; Gajewiak, J.; Macraild, C.A.; Chou, D.H.; Disotuar, M.M.; Smith, N.A.; Miller, C.; Erchegyi, J.; Rivier, J.E.; Olivera, B.M.; et al. A minimized human insulin-receptor-binding motif revealed in a Conus geographus venom insulin. Nat. Struct. Mol. Biol. 2016, 23, 916–920. [Google Scholar] [CrossRef]
- Sajid, W.; Kulahin, N.; Schluckebier, G.; Ribel, U.; Henderson, H.R.; Tatar, M.; Hansen, B.F.; Svendsen, A.M.; Kiselyov, V.V.; Nørgaard, P.; et al. Structural and biological properties of the Drosophila insulin-like peptide 5 show evolutionary conservation. J. Biol. Chem. 2011, 286, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Ahorukomeye, P.; Disotuar, M.M.; Gajewiak, J.; Karanth, S.; Watkins, M.; Robinson, S.D.; Salcedo, P.F.; Smith, N.A.; Smith, B.J.; Schlege, A.; et al. Fish-hunting cone snail venoms are a rich source of minimized ligands of the vertebrate insulin receptor. Elife 2019, 8, e41574. [Google Scholar] [CrossRef] [PubMed]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea anemones: Quiet achievers in the field of peptide toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnarjuna, B.; MacRaild, C.A.; Sunanda, P.; Morales, R.A.V.; Peigneur, S.; Macrander, J.; Yu, H.H.; Daly, M.; Raghothama, S.; Dhawan, V.; et al. Structure, folding and stability of a minimal homologue from Anemonia sulcata of the sea anemone potassium channel blocker ShK. Peptides 2018, 99, 169–178. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Villegas-Moreno, J.; Mitchell, M.L.; Csoti, A.; Peigneur, S.; Amero, C.; Pennington, M.W.; Tytgat, J.; Panyi, G.; Norton, R.S. Synthesis, folding, structure and activity of a predicted peptide from the sea anemone Oulactis sp. with an ShKT fold. Toxicon 2018, 150, 50–59. [Google Scholar] [CrossRef]
- Sunanda, P.; Krishnarjuna, B.; Peigneur, S.; Mitchell, M.L.; Estrada, R.; Villegas-Moreno, J.; Pennington, M.W.; Tytgat, J.; Norton, R.S. Identification, chemical synthesis, structure, and function of a new KV1 channel blocking peptide from Oulactis sp. Pept. Sci. 2018, 110, e24073. [Google Scholar] [CrossRef]
- Stephenson, T.A. The British Sea Anemones; The Ray Society: London, UK, 1928; Volume I. [Google Scholar]
- Fitt, W.K. Photosynthesis, respiration, and contribution to community productivity of the symbiotic sea anemone Anthopleura elegantissima (Brandt, 1835). J. Exp. Mar. Bio. Ecol. 1982, 61, 213–232. [Google Scholar] [CrossRef]
- Sebens, K. The energetics of asexual reproduction and colony formation in benthic marine invertebrates. Am. Zool. 1979, 19, 683–699. [Google Scholar] [CrossRef]
- Chomsky, O.; Kamenir, Y.; Hyams, M.; Dubinsky, Z.; Chadwick-Furman, N.E. Effects of feeding regime on growth rate in the Mediterranean sea anemone Actinia equina (Linnaeus). J. Exp. Mar. Bio. Ecol. 2004, 299, 217–229. [Google Scholar] [CrossRef]
- Fautin, D.G. Reproduction of Cnidaria. Can. J. Zool. 2002, 80, 1735–1754. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insulin-like Protein (ILP) | Phylum/Class Species | Function/ Tissue Expression | A-Chain% Similarity | B-Chain% Similarity | Ref. |
|---|---|---|---|---|---|
| Nemve_207484 (A7S6C3) | Cnidaria/Anthozoa Nematostella vectensis | predicted ILP (larval whole animal) | 61.5 | 40.9 | [25,44] |
| Insulin (P010308) | Chordata/Mammalia Homo sapiens | metabolize energy (pancreas) | 54.5 | 24.4 | [33] |
| Con-Ins 1A (A0A0B5AC95) | Mollusca/Gastropoda Conus geographus | induce insulin coma (venom) | 50.0 | 34.2 | [14] |
| Bombyxin (Q17192) | Arthropoda/Insecta Bombyx mori | growth development (brain) | 50.0 | 32.5 | [8] |
| INS_APLCA (Q9NDE7) | Mollusca/Gastropoda Aplysia californica | metabolize energy (ganglia and cellular clusters) | 31.4 | 28.8 | [10] |
| INS-3 (Q09628; WBGene00002086) | Nematoda Caenorhabditis elegans | antagonist of the IIS 1 pathway (coelomocyte; egg-laying apparatus; gonad; head muscle; and nervous system) | 23.4 | - | [9,48] |
| INS-17 (G5EFH1; WBGene0000210) | Nematoda Caenorhabditis elegans | agonist of the IIS 1 pathway (egg-laying apparatus; gonad; head muscle; neurons; and somatic nervous system.) | 23.1 | - | [9,48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitchell, M.L.; Hossain, M.A.; Lin, F.; Pinheiro-Junior, E.L.; Peigneur, S.; Wai, D.C.C.; Delaine, C.; Blyth, A.J.; Forbes, B.E.; Tytgat, J.; et al. Identification, Synthesis, Conformation and Activity of an Insulin-like Peptide from a Sea Anemone. Biomolecules 2021, 11, 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121785
Mitchell ML, Hossain MA, Lin F, Pinheiro-Junior EL, Peigneur S, Wai DCC, Delaine C, Blyth AJ, Forbes BE, Tytgat J, et al. Identification, Synthesis, Conformation and Activity of an Insulin-like Peptide from a Sea Anemone. Biomolecules. 2021; 11(12):1785. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121785
Chicago/Turabian StyleMitchell, Michela L., Mohammed Akhter Hossain, Feng Lin, Ernesto L. Pinheiro-Junior, Steve Peigneur, Dorothy C. C. Wai, Carlie Delaine, Andrew J. Blyth, Briony E. Forbes, Jan Tytgat, and et al. 2021. "Identification, Synthesis, Conformation and Activity of an Insulin-like Peptide from a Sea Anemone" Biomolecules 11, no. 12: 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121785