
Mechanism of Zn2+ and Ca2+ Binding to Human S100A1

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Purification
2.2. Electrospray Mass-Spectrometry (ESI-MS)
2.3. Isothermal Titration Calorimetry
2.4. Dynamic Light Scattering (DLS)
2.5. Differential Scanning Fluorimetry (nanoDSF) and Light Scattering
2.6. Circular Dichroism
2.7. QM/MM Molecular Modeling
3. Results
3.1. Stoichiometry of S100A1-Metal Binding
3.2. Affinity of S100A1-Metal Binding
3.3. Effect of Ca2+ and Zn2+ on Secondary Structure of S100A1
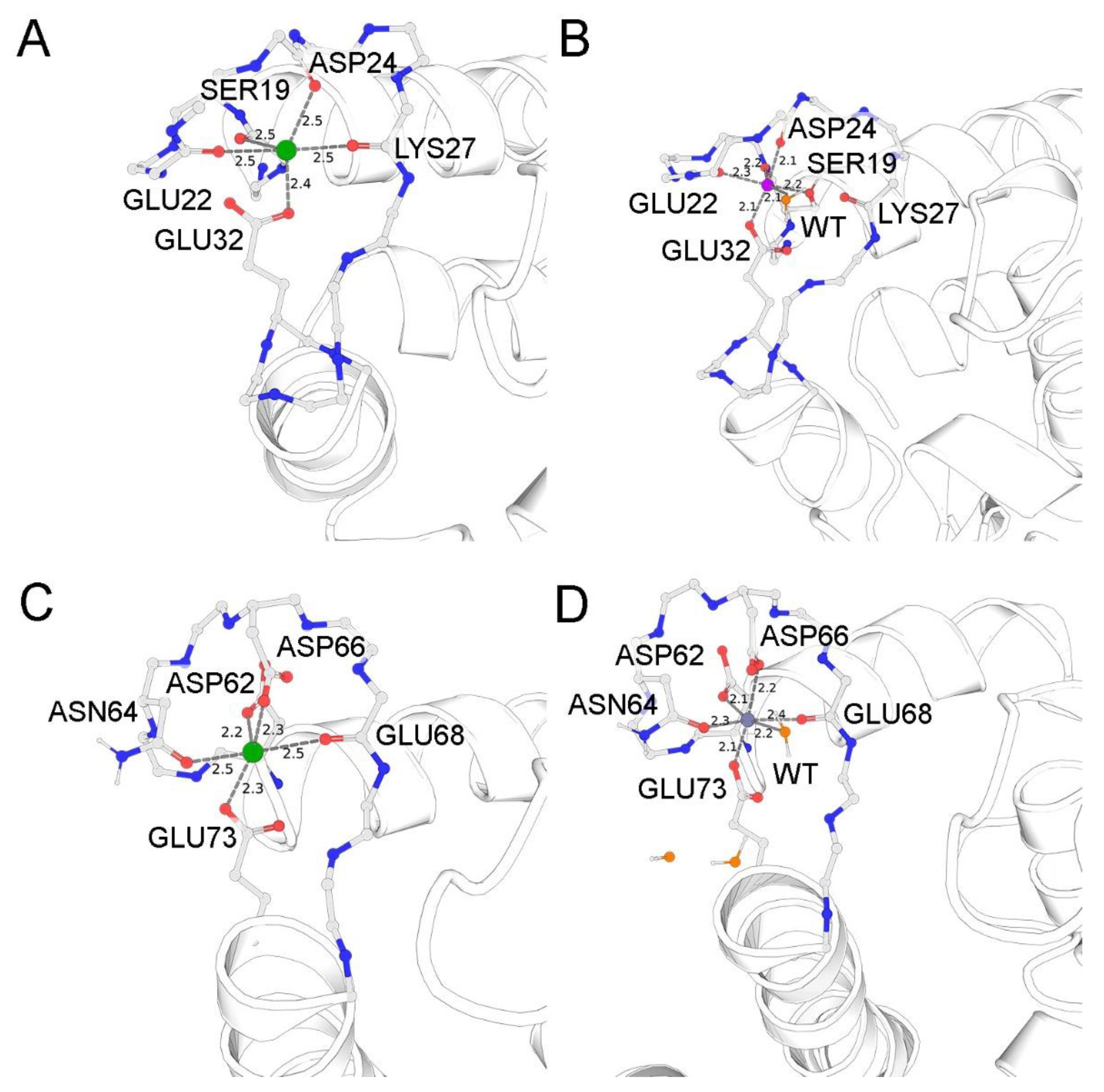
3.4. Putative Zn2+-Binding Sites in S100A1
3.5. Effects of Metal Binding on Dimeric Structure of S100A1
3.6. Effects of Metal Binding on Conformational Properties of S100A1
3.7. Effects of Metal Binding on Thermal Stability and Aggregation of S100A1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Donato, R. Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type. Biochim. Biophys. Acta 1999, 1450, 191–231. [Google Scholar] [CrossRef] [Green Version]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [Green Version]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Cristovao, J.S.; Gomes, C.M. S100 Proteins in Alzheimer’s Disease. Front. Neurosci. 2019, 13, 463. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, H.; Zhang, L.; Xu, Y.; Deng, W.; Zhu, H.; Qin, C. S100B transgenic mice develop features of Parkinson’s disease. Arch. Med. Res. 2011, 42, 1–7. [Google Scholar] [CrossRef]
- Mori, T.; Koyama, N.; Arendash, G.W.; Horikoshi-Sakuraba, Y.; Tan, J.; Town, T. Overexpression of human S100B exacerbates cerebral amyloidosis and gliosis in the Tg2576 mouse model of Alzheimer’s disease. Glia 2010, 58, 300–314. [Google Scholar] [CrossRef] [Green Version]
- Gerlai, R.; Wojtowicz, J.M.; Marks, A.; Roder, J. Overexpression of a calcium-binding protein, S100 beta, in astrocytes alters synaptic plasticity and impairs spatial learning in transgenic mice. Learn. Mem. 1995, 2, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliuchnikova, A.A.; Samokhina, N.I.; Ilina, I.Y.; Karpov, D.S.; Pyatnitskiy, M.A.; Kuznetsova, K.G.; Toropygin, I.Y.; Kochergin, S.A.; Alekseev, I.B.; Zgoda, V.G.; et al. Human aqueous humor proteome in cataract, glaucoma, and pseudoexfoliation syndrome. Proteomics 2016, 16, 1938–1946. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Harper, M.M.; Kecova, H.; Adamus, G.; Kardon, R.H.; Grozdanic, S.D.; Kuehn, M.H. Neuroinflammation in advanced canine glaucoma. Mol. Vis. 2010, 16, 2092–2108. [Google Scholar]
- Baudier, J.; Deloulme, J.C.; Shaw, G.S. The Zn2+ and Ca2+-binding S100B and S100A1 proteins: Beyond the myths. Biol. Rev. Camb. Philos. Soc. 2020, 95, 738–758. [Google Scholar] [CrossRef]
- Duarte-Costa, S.; Castro-Ferreira, R.; Neves, J.S.; Leite-Moreira, A.F. S100A1: A major player in cardiovascular performance. Physiol. Res. 2014, 63, 669–681. [Google Scholar] [CrossRef]
- Donato, R.; Heizmann, C.W. S100B Protein in the Nervous System and Cardiovascular Apparatus in Normal and Pathological Conditions. Cardiovasc. Psychiatry Neurol. 2010, 2010, 929712. [Google Scholar] [CrossRef] [Green Version]
- Baudier, J.; Glasser, N.; Gerard, D. Ions binding to S100 proteins. I. Calcium- and zinc-binding properties of bovine brain S100 alpha alpha, S100a (alpha beta), and S100b (beta beta) protein: Zn2+ regulates Ca2+ binding on S100b protein. J. Biol. Chem. 1986, 261, 8192–8203. [Google Scholar] [CrossRef]
- Zimmer, D.B.; Weber, D.J. The Calcium-Dependent Interaction of S100B with Its Protein Targets. Cardiovasc. Psychiatry Neurol. 2010, 2010, 728052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treves, S.; Scutari, E.; Robert, M.; Groh, S.; Ottolia, M.; Prestipino, G.; Ronjat, M.; Zorzato, F. Interaction of S100A1 with the Ca2+ release channel (ryanodine receptor) of skeletal muscle. Biochemistry 1997, 36, 11496–11503. [Google Scholar] [CrossRef] [PubMed]
- Frederickson, C.J.; Koh, J.Y.; Bush, A.I. The neurobiology of zinc in health and disease. Nat. Rev. Neurosci. 2005, 6, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Krall, R.F.; Tzounopoulos, T.; Aizenman, E. The Function and Regulation of Zinc in the Brain. Neuroscience 2021, 457, 235–258. [Google Scholar] [CrossRef]
- Li, Y.; Andereggen, L.; Yuki, K.; Omura, K.; Yin, Y.; Gilbert, H.Y.; Erdogan, B.; Asdourian, M.S.; Shrock, C.; de Lima, S.; et al. Mobile zinc increases rapidly in the retina after optic nerve injury and regulates ganglion cell survival and optic nerve regeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E209–E218. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, M.; Osborne, N.N. Recent advances in the understanding of the role of zinc in ocular tissues. Metallomics 2014, 6, 189–200. [Google Scholar] [CrossRef]
- Tsvetkov, P.O.; Roman, A.Y.; Baksheeva, V.E.; Nazipova, A.A.; Shevelyova, M.P.; Vladimirov, V.I.; Buyanova, M.F.; Zinchenko, D.V.; Zamyatnin, A.A., Jr.; Devred, F.; et al. Functional Status of Neuronal Calcium Sensor-1 Is Modulated by Zinc Binding. Front. Mol. Neurosci. 2018, 11, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Permyakov, S.E.; Cherskaya, A.M.; Wasserman, L.A.; Khokhlova, T.I.; Senin, I.I.; Zargarov, A.A.; Zinchenko, D.V.; Zernii, E.Y.; Lipkin, V.M.; Philippov, P.P.; et al. Recoverin is a zinc-binding protein. J. Proteome. Res. 2003, 2, 51–57. [Google Scholar] [CrossRef]
- Baksheeva, V.E.; Baldin, A.V.; Zalevsky, A.O.; Nazipova, A.A.; Kazakov, A.S.; Vladimirov, V.I.; Gorokhovets, N.V.; Devred, F.; Philippov, P.P.; Bazhin, A.V.; et al. Disulfide Dimerization of Neuronal Calcium Sensor-1: Implications for Zinc and Redox Signaling. Int. J. Mol. Sci. 2021, 22, 12602. [Google Scholar] [CrossRef]
- Ostendorp, T.; Diez, J.; Heizmann, C.W.; Fritz, G. The crystal structures of human S100B in the zinc- and calcium-loaded state at three pH values reveal zinc ligand swapping. Biochim. Biophys. Acta 2011, 1813, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Wilder, P.T.; Varney, K.M.; Weiss, M.B.; Gitti, R.K.; Weber, D.J. Solution structure of zinc- and calcium-bound rat S100B as determined by nuclear magnetic resonance spectroscopy. Biochemistry 2005, 44, 5690–5702. [Google Scholar] [CrossRef]
- Barber, K.R.; McClintock, K.A.; Jamieson, G.A., Jr.; Dimlich, R.V.; Shaw, G.S. Specificity and Zn2+ enhancement of the S100B binding epitope TRTK-12. J. Biol. Chem. 1999, 274, 1502–1508. [Google Scholar] [CrossRef] [Green Version]
- Marlatt, N.M.; Spratt, D.E.; Shaw, G.S. Codon optimization for enhanced Escherichia coli expression of human S100A11 and S100A1 proteins. Protein. Expr. Purif. 2010, 73, 58–64. [Google Scholar] [CrossRef]
- Wang, B.; Boeckel, G.R.; Huynh, L.; Nguyen, L.; Cao, W.; De La Cruz, E.M.; Kaftan, E.J.; Ehrlich, B.E. Neuronal Calcium Sensor 1 Has Two Variants with Distinct Calcium Binding Characteristics. PLoS ONE 2016, 11, e0161414. [Google Scholar] [CrossRef]
- Kazakov, A.S.; Sofin, A.D.; Avkhacheva, N.V.; Denesyuk, A.I.; Deryusheva, E.I.; Rastrygina, V.A.; Sokolov, A.S.; Permyakova, M.E.; Litus, E.A.; Uversky, V.N.; et al. Interferon Beta Activity Is Modulated via Binding of Specific S100 Proteins. Int. J. Mol. Sci. 2020, 21, 9473. [Google Scholar] [CrossRef]
- Makarov, A.A.; Tsvetkov, P.O.; Villard, C.; Esquieu, D.; Pourroy, B.; Fahy, J.; Braguer, D.; Peyrot, V.; Lafitte, D. Vinflunine, a novel microtubule inhibitor, suppresses calmodulin interaction with the microtubule-associated protein STOP. Biochemistry 2007, 46, 14899–14906. [Google Scholar] [CrossRef]
- Petrushanko, I.Y.; Lobachev, V.M.; Kononikhin, A.S.; Makarov, A.A.; Devred, F.; Kovacic, H.; Kubatiev, A.A.; Tsvetkov, P.O. Oxidation of capital ES, Cyrillicsmall a, Cyrillic2+-Binding Domain of NADPH Oxidase 5 (NOX5): Toward Understanding the Mechanism of Inactivation of NOX5 by ROS. PLoS ONE 2016, 11, e0158726. [Google Scholar] [CrossRef]
- Garnier, C.; Devred, F.; Byrne, D.; Puppo, R.; Roman, A.Y.; Malesinski, S.; Golovin, A.V.; Lebrun, R.; Ninkina, N.N.; Tsvetkov, P.O. Zinc binding to RNA recognition motif of TDP-43 induces the formation of amyloid-like aggregates. Sci. Rep. 2017, 7, 6812. [Google Scholar] [CrossRef] [Green Version]
- Vologzhannikova, A.A.; Shevelyova, M.P.; Kazakov, A.S.; Sokolov, A.S.; Borisova, N.I.; Permyakov, E.A.; Kircheva, N.; Nikolova, V.; Dudev, T.; Permyakov, S.E. Strontium Binding to alpha-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family. Biomolecules 2021, 11, 1158. [Google Scholar] [CrossRef]
- Sreerama, N.; Venyaminov, S.Y.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Inclusion of denatured proteins with native proteins in the analysis. Anal. Biochem. 2000, 287, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Golovin, A.V.; Devred, F.; Yatoui, D.; Roman, A.Y.; Zalevsky, A.O.; Puppo, R.; Lebrun, R.; Guerlesquin, F.; Tsvetkov, P.O. Zinc Binds to RRM2 Peptide of TDP-43. Int. J. Mol. Sci. 2020, 21, 9080. [Google Scholar] [CrossRef] [PubMed]
- Melville, Z.; Aligholizadeh, E.; McKnight, L.E.; Weber, D.J.; Pozharski, E.; Weber, D.J. X-ray crystal structure of human calcium-bound S100A1. Acta Crystallogr. F Struct. Biol. Commun. 2017, 73, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, M.; Ruszczynska-Bartnik, K.; Budzinska, M.; Jaremko, L.; Jaremko, M.; Zdanowski, K.; Bierzynski, A.; Ejchart, A. Impact of calcium binding and thionylation of S100A1 protein on its nuclear magnetic resonance-derived structure and backbone dynamics. Biochemistry 2013, 52, 1149–1159. [Google Scholar] [CrossRef]
- Deng, L.; Sun, N.; Kitova, E.N.; Klassen, J.S. Direct quantification of protein-metal ion affinities by electrospray ionization mass spectrometry. Anal. Chem. 2010, 82, 2170–2174. [Google Scholar] [CrossRef]
- Potier, N.; Rogniaux, H.; Chevreux, G.; Van Dorsselaer, A. Ligand-metal ion binding to proteins: Investigation by ESI mass spectrometry. Methods Enzymol. 2005, 402, 361–389. [Google Scholar] [CrossRef] [PubMed]
- Zhukov, I.; Ejchart, A.; Bierzynski, A. Structural and motional changes induced in apo-S100A1 protein by the disulfide formation between its Cys 85 residue and beta-mercaptoethanol. Biochemistry 2008, 47, 640–650. [Google Scholar] [CrossRef]
- Goch, G.; Vdovenko, S.; Kozlowska, H.; Bierzynski, A. Affinity of S100A1 protein for calcium increases dramatically upon glutathionylation. FEBS J. 2005, 272, 2557–2565. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, L.C.; Donor, M.T.; Prell, J.S.; Harms, M.J. Multiple Evolutionary Origins of Ubiquitous Cu2+ and Zn2+ Binding in the S100 Protein Family. PLoS ONE 2016, 11, e0164740. [Google Scholar] [CrossRef]
- Laitaoja, M.; Valjakka, J.; Janis, J. Zinc coordination spheres in protein structures. Inorg. Chem. 2013, 52, 10983–10991. [Google Scholar] [CrossRef] [PubMed]
- Peschke, M.; Blades, A.T.; Kebarle, P. Binding Energies for Doubly-Charged Ions M2+ = Mg2+, Ca2+ and Zn2+ with the Ligands L = H2O, Acetone and N-methylacetamide in Complexes M for n = 1 to 7 from Gas Phase Equilibria Determinations and Theoretical Calculations. J. Am. Chem. Soc. 2000, 122, 10440–10449. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Monodentate versus Bidentate Carboxylate Binding in Magnesium and Calcium Proteins: What Are the Basic Principles? J. Phys. Chem. B 2004, 108, 4546–4557. [Google Scholar] [CrossRef]
- Sitaramayya, A. Calcium-dependent activation of guanylate cyclase by S100b. Adv. Exp. Med. Biol. 2002, 514, 389–398. [Google Scholar] [CrossRef]
- Egelman, D.M.; Montague, P.R. Calcium dynamics in the extracellular space of mammalian neural tissue. Biophys. J. 1999, 76, 1856–1867. [Google Scholar] [CrossRef] [Green Version]
- Assaf, S.Y.; Chung, S.H. Release of endogenous Zn2+ from brain tissue during activity. Nature 1984, 308, 734–736. [Google Scholar] [CrossRef]
- Lyubartseva, G.; Lovell, M.A. A potential role for zinc alterations in the pathogenesis of Alzheimer’s disease. Biofactors 2012, 38, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Haase, H.; Rink, L. Functional significance of zinc-related signaling pathways in immune cells. Annu. Rev. Nutr. 2009, 29, 133–152. [Google Scholar] [CrossRef]
- Brett, W.; Mandinova, A.; Remppis, A.; Sauder, U.; Ruter, F.; Heizmann, C.W.; Aebi, U.; Zerkowski, H.R. Translocation of S100A1(1) calcium binding protein during heart surgery. Biochem. Biophys. Res. Commun. 2001, 284, 698–703. [Google Scholar] [CrossRef]
- Huttunen, H.J.; Kuja-Panula, J.; Sorci, G.; Agneletti, A.L.; Donato, R.; Rauvala, H. Coregulation of neurite outgrowth and cell survival by amphoterin and S100 proteins through receptor for advanced glycation end products (RAGE) activation. J. Biol. Chem. 2000, 275, 40096–40105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zernii, E.Y.; Grigoriev, I.I.; Nazipova, A.A.; Scholten, A.; Kolpakova, T.V.; Zinchenko, D.V.; Kazakov, A.S.; Senin, I.I.; Permyakov, S.E.; Dell’Orco, D.; et al. Regulatory function of the C-terminal segment of guanylate cyclase-activating protein 2. Biochim. Biophys. Acta 2015, 1854, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Landar, A.; Rustandi, R.R.; Weber, D.J.; Zimmer, D.B. S100A1 utilizes different mechanisms for interacting with calcium-dependent and calcium-independent target proteins. Biochemistry 1998, 37, 17429–17438. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, D.B.; Cornwall, E.H.; Reynolds, P.D.; Donald, C.M. S100A1 regulates neurite organization, tubulin levels, and proliferation in PC12 cells. J. Biol. Chem. 1998, 273, 4705–4711. [Google Scholar] [CrossRef] [Green Version]
- Heierhorst, J.; Mann, R.J.; Kemp, B.E. Interaction of the recombinant S100A1 protein with twitchin kinase, and comparison with other Ca2+-binding proteins. Eur. J. Biochem. 1997, 249, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, A.Y.; Devred, F.; Byrne, D.; La Rocca, R.; Ninkina, N.N.; Peyrot, V.; Tsvetkov, P.O. Zinc Induces Temperature-Dependent Reversible Self-Assembly of Tau. J. Mol. Biol. 2019, 431, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Magneson, G.R.; Puvathingal, J.M.; Ray, W.J., Jr. The concentrations of free Mg2+ and free Zn2+ in equine blood plasma. J. Biol. Chem. 1987, 262, 11140–11148. [Google Scholar] [CrossRef]
- Hesketh, J.E. Zinc-stimulated microtubule assembly and evidence for zinc binding to tubulin. Int. J. Biochem. 1982, 14, 983–990. [Google Scholar] [CrossRef]
- Hesketh, J.E. Zinc binding to tubulin. Int. J. Biochem. 1983, 15, 743–746. [Google Scholar] [CrossRef]
- Deinum, J.; Baudier, J.; Briving, C.; Rosengren, L.; Wallin, M.; Gerard, D.; Haglid, K. The effect of S-100a and S-100b proteins and Zn2+ on the assembly of brain microtubule proteins in vitro. FEBS Lett. 1983, 163, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Faget, K.Y. Zinc takes the center stage: Its paradoxical role in Alzheimer’s disease. Brain Res. Brain Res. Rev. 2003, 41, 44–56. [Google Scholar] [CrossRef]
- Afanador, L.; Roltsch, E.A.; Holcomb, L.; Campbell, K.S.; Keeling, D.A.; Zhang, Y.; Zimmer, D.B. The Ca2+ sensor S100A1 modulates neuroinflammation, histopathology and Akt activity in the PSAPP Alzheimer’s disease mouse model. Cell Calcium 2014, 56, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Bhatty, A.; Baig, S.; Fawwad, A.; Rubab, Z.E.; Shahid, M.A.; Waris, N. Association of Zinc Transporter-8 Autoantibody (ZnT8A) with Type 1 Diabetes Mellitus. Cureus 2020, 12, e7263. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Huang, Y.L. Chromium, zinc and magnesium status in type 1 diabetes. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, X.; Hua, Z.; Ma, J.; Liu, Z.; Chen, H.; Cui, Z. S100A1 promotes cell proliferation and migration and is associated with lymph node metastasis in ovarian cancer. Discov. Med. 2017, 23, 235–245. [Google Scholar] [PubMed]
- Morii, K.; Tanaka, R.; Takahashi, Y.; Kuwano, R. Cloning of cDNAs encoding human S-100 alpha and beta subunits and their differential expression in human tumor cell lines. J. Neurosci. Res. 1992, 32, 27–33. [Google Scholar] [CrossRef]
- Kato, K.; Haimoto, H.; Ariyoshi, Y.; Horisawa, M.; Washida, H.; Kimura, S. High levels of S-100a0 (alpha alpha) protein in tumor tissues and in sera of patients with renal cell carcinoma. JPN J. Cancer Res. 1985, 76, 856–862. [Google Scholar]
- Cocchia, D.; Michetti, F.; Donato, R. Immunochemical and immuno-cytochemical localization of S-100 antigen in normal human skin. Nature 1981, 294, 85–87. [Google Scholar] [CrossRef]
- Sheline, C.T.; Cai, A.L.; Zhu, J.; Shi, C. Serum or target deprivation-induced neuronal death causes oxidative neuronal accumulation of Zn2+ and loss of NAD+. Eur. J. Neurosci. 2010, 32, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Makhov, P.; Golovine, K.; Uzzo, R.G.; Rothman, J.; Crispen, P.L.; Shaw, T.; Scoll, B.J.; Kolenko, V.M. Zinc chelation induces rapid depletion of the X-linked inhibitor of apoptosis and sensitizes prostate cancer cells to TRAIL-mediated apoptosis. Cell Death Differ. 2008, 15, 1745–1751. [Google Scholar] [CrossRef] [Green Version]
- Kontoghiorghes, G.J.; Efstathiou, A.; Ioannou-Loucaides, S.; Kolnagou, A. Chelators controlling metal metabolism and toxicity pathways: Applications in cancer prevention, diagnosis and treatment. Hemoglobin 2008, 32, 217–227. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion | N1 | K1a, M−1 | K1D, M | ∆H1, kcal M−1 | ∆S1, cal M−1 K−1 | N2 | K2a, M−1 | K1D, M | ∆H2, kcal M−1 | ∆S2, cal M−1 K−1 |
|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | 0.9 | 6.1 ± 2.1 × 106 | 1.6 ± 0.6× 10−7 | −16.9 ± 0.2 | −23.3 | 1.0 | 4.2 ± 0.6 × 104 | 2.4 ± 0.3× 10−5 | 4.0 ± 0.8 | 34.1 |
| Zn2+ | 0.9 | 2.6 ± 0.7 × 108 | 3.8 ± 1.0× 10−9 | −13.2 ± 0.2 | −4.2 | 0.9 | 1.3 ± 0.4 × 106 | 7.7 ± 2.4× 10−7 | 3.3 ± 0.4 | 38.5 |
| Ca2+ in Zn2+ | no binding | |||||||||
| Zn2+ in Ca2+ | no binding | |||||||||
| Protein State | α-Helices, % | β-Structure, % | Turns, % | Unordered Structure, % |
|---|---|---|---|---|
| apo | 70.58 ± 0.56 | 5.17 ± 0.09 | 8.01 ± 0.40 | 16.02 ± 0.25 |
| Ca2+-loaded | 68.05 ± 0.56 | 4.05 ± 0.09 | 9.28 ± 0.40 | 18.65 ± 0.25 |
| Zn2+-loaded | 65.10 ± 0.56 | 4.13 ± 0.09 | 11.17 ± 0.40 | 19.50 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baksheeva, V.E.; Roman, A.Y.; Villard, C.; Devred, F.; Byrne, D.; Yatoui, D.; Zalevsky, A.O.; Vologzhannikova, A.A.; Sokolov, A.S.; Permyakov, S.E.; et al. Mechanism of Zn2+ and Ca2+ Binding to Human S100A1. Biomolecules 2021, 11, 1823. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121823
Baksheeva VE, Roman AY, Villard C, Devred F, Byrne D, Yatoui D, Zalevsky AO, Vologzhannikova AA, Sokolov AS, Permyakov SE, et al. Mechanism of Zn2+ and Ca2+ Binding to Human S100A1. Biomolecules. 2021; 11(12):1823. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121823
Chicago/Turabian StyleBaksheeva, Viktoriia E., Andrei Yu. Roman, Claude Villard, François Devred, Deborah Byrne, Dahbia Yatoui, Arthur O. Zalevsky, Alisa A. Vologzhannikova, Andrey S. Sokolov, Sergei E. Permyakov, and et al. 2021. "Mechanism of Zn2+ and Ca2+ Binding to Human S100A1" Biomolecules 11, no. 12: 1823. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121823