
Regulation of Phosphoinositide Levels in the Retina by Protein Tyrosine Phosphatase 1B and Growth Factor Receptor-Bound Protein 14

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
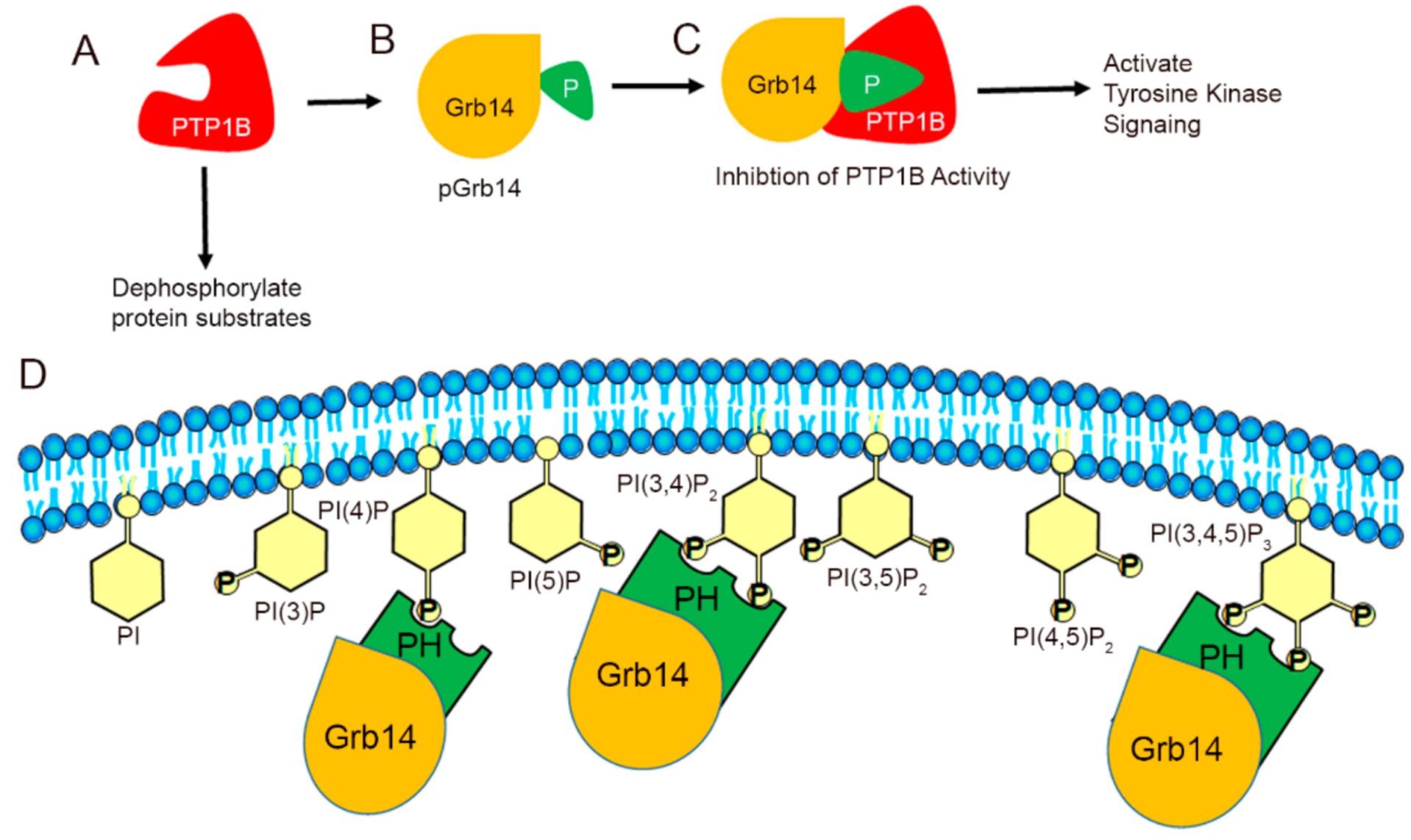
Abstract
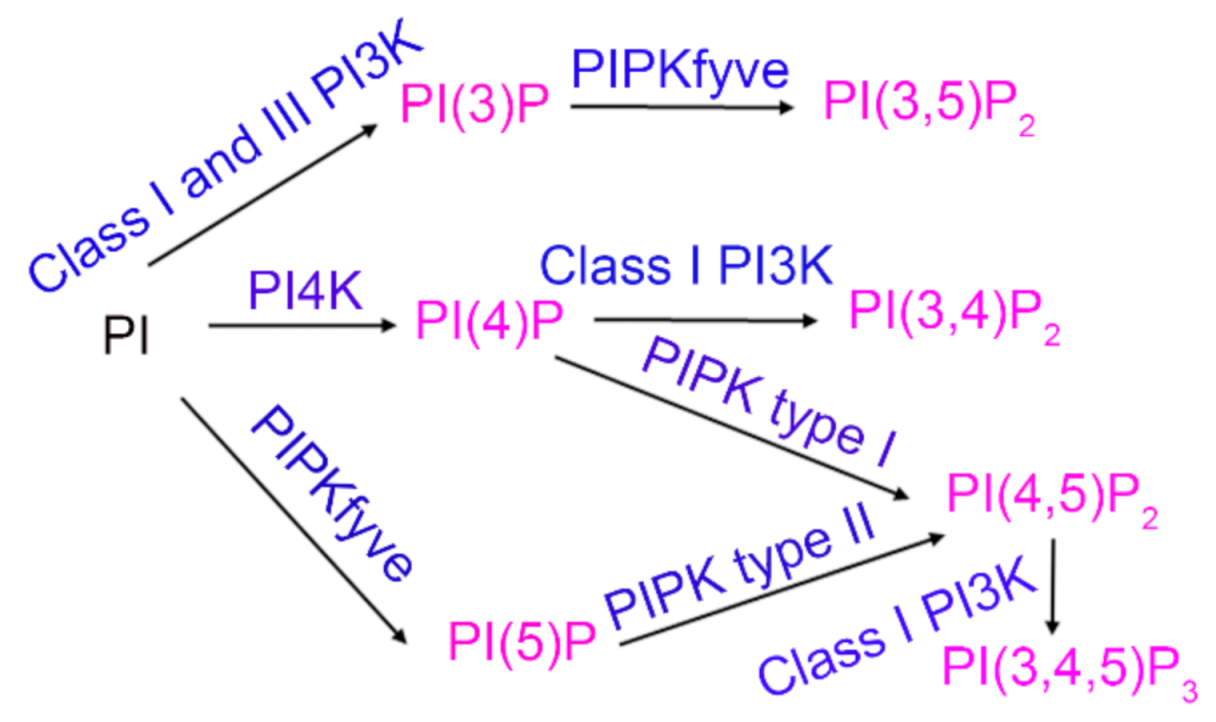
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Methods
2.3.1. Extraction and Measurement of Phosphoinositides from the Retina
2.3.2. Immunoblot Analysis
2.3.3. In Silico Modeling of Grb14 and PTP1B Interactions
3. Results
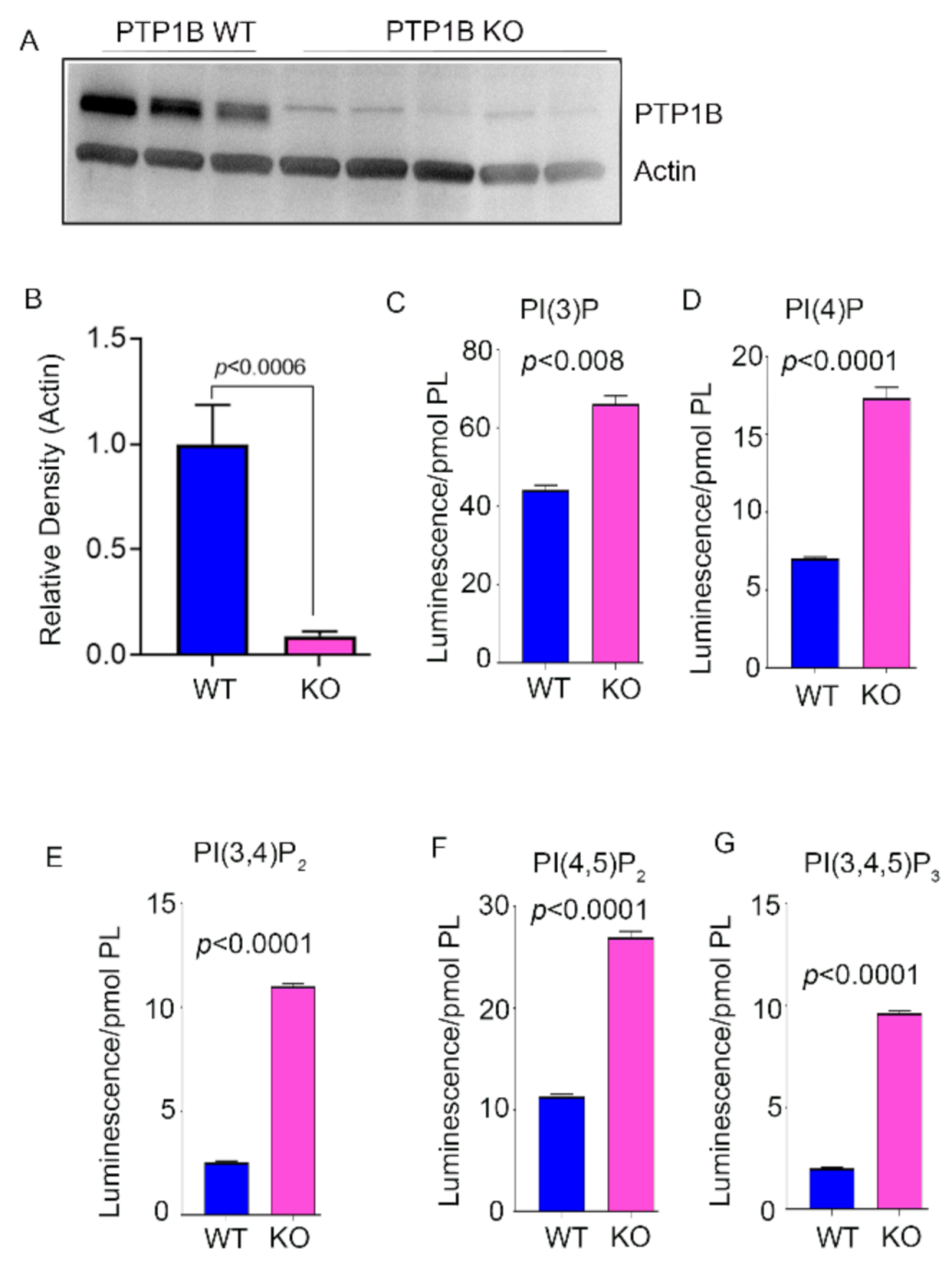
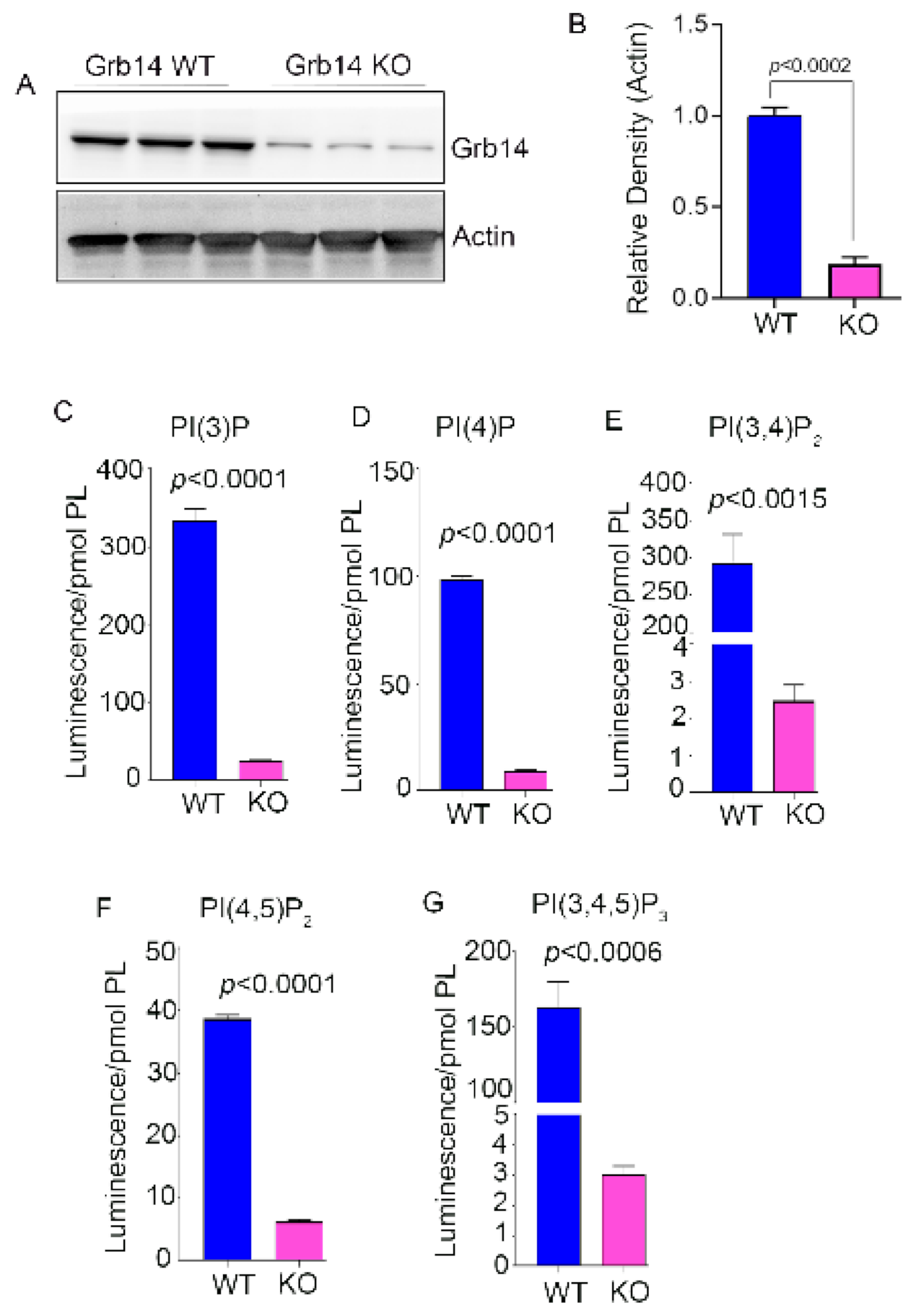
3.1. Phosphoinositide Levels in PTP1B and Grb14 KO Mouse Retina
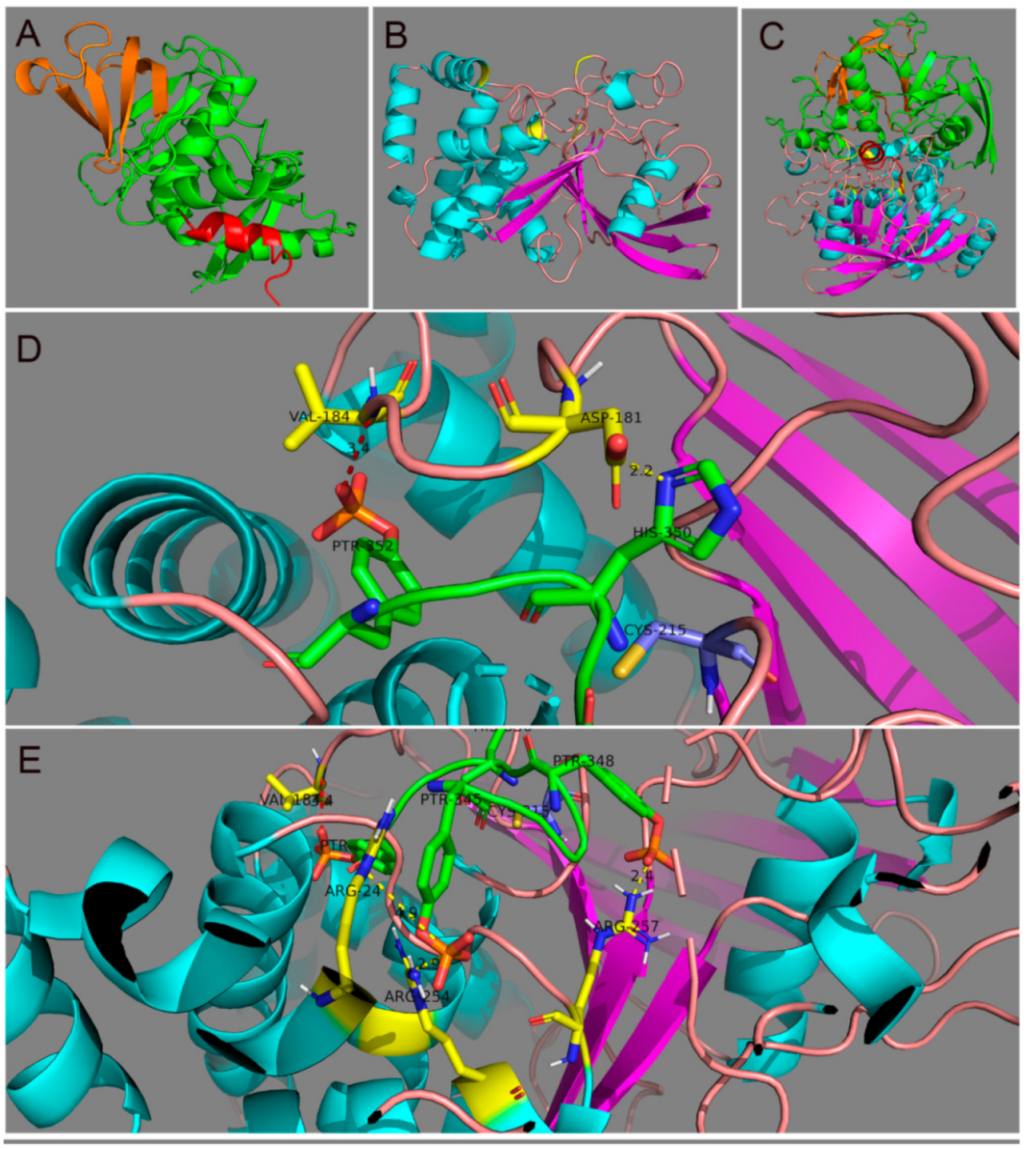
3.2. Identification of Putative Residues in PTP1B That Mediate the Interaction with Grb14
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Stuible, M.; Tremblay, M.L. In control at the ER: PTP1B and the down-regulation of RTKs by dephosphorylation and endocytosis. Trends Cell Biol. 2010, 20, 672–679. [Google Scholar] [CrossRef]
- Depetris, R.S.; Hu, J.; Gimpelevich, I.; Holt, L.J.; Daly, R.J.; Hubbard, S.R. Structural basis for inhibition of the insulin receptor by the adaptor protein Grb14. Mol. Cell 2005, 20, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Rajala, A.; Daly, R.J.; Tanito, M.; Allen, D.T.; Holt, L.J.; Lobanova, E.S.; Arshavsky, V.Y.; Rajala, R.V. Growth factor receptor-bound protein 14 undergoes light-dependent intracellular translocation in rod photoreceptors: Functional role in retinal insulin receptor activation. Biochemistry 2009, 48, 5563–5572. [Google Scholar] [CrossRef] [Green Version]
- Rajala, R.V.; Tanito, M.; Neel, B.G.; Rajala, A. Enhanced retinal insulin receptor-activated neuroprotective survival signal in mice lacking the protein-tyrosine phosphatase-1B gene. J. Biol. Chem. 2010, 285, 8894–8904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, R.V.; Rajala, A. Neuroprotective role of protein tyrosine phosphatase-1B in rod photoreceptor neurons. Protein Cell 2013, 4, 890–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, A.; Tanito, M.; Le, Y.Z.; Kahn, C.R.; Rajala, R.V. Loss of neuroprotective survival signal in mice lacking insulin receptor gene in rod photoreceptor cells. J. Biol. Chem. 2008, 283, 19781–19792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, A.; Wang, Y.; Rajala, R.V. Activation of oncogenic tyrosine kinase signaling promotes insulin receptor-mediated cone photoreceptor survival. Oncotarget 2016, 7, 46924–46942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooney, G.J.; Lyons, R.J.; Crew, A.J.; Jensen, T.E.; Molero, J.C.; Mitchell, C.J.; Biden, T.J.; Ormandy, C.J.; James, D.E.; Daly, R.J. Improved glucose homeostasis and enhanced insulin signalling in Grb14-deficient mice. EMBO J. 2004, 23, 582–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.C.; Weeks, K.L.; Gao, X.M.; Williams, R.B.; Bernardo, B.C.; Kiriazis, H.; Matthews, V.B.; Woodcock, E.A.; Bouwman, R.D.; Mollica, J.P.; et al. PI3K(p110 alpha) protects against myocardial infarction-induced heart failure: Identification of PI3K-regulated miRNA and mRNA. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basavarajappa, D.K.; Gupta, V.K.; Dighe, R.; Rajala, A.; Rajala, R.V. Phosphorylated Grb14 Is an Endogenous Inhibitor of Retinal Protein Tyrosine Phosphatase 1B, and Light-Dependent Activation of Src Phosphorylates Grb14. Mol. Cell Biol. 2011, 31, 3975–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Rajala, R.V.S. Signaling roles of phosphoinositides in the retina. J. Lipid Res. 2021, 100041. [Google Scholar] [CrossRef] [PubMed]
- Rajala, A.; McCauley, A.; Brush, R.S.; Nguyen, K.; Rajala, R.V.S. Phosphoinositide Lipids in Ocular Tissues. Biology 2020, 9, 125. [Google Scholar] [CrossRef]
- Rouser, G.; Siakotos, A.N.; Fleischer, S. Quantitative analysis of phospholipids by thin-layer chromatography and phosphorus analysis of spots. Lipids 1966, 1, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Gortner, W.A. An evaluation of micromethods for phospholipids. J. Biol. Chem. 1945, 159, 97–100. [Google Scholar] [CrossRef]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, A.K.; Peters, G.G.; Møller, K.B.; Iversen, L.F.; Kastrup, J.S. Water-molecule network and active-site flexibility of apo protein tyrosine phosphatase 1B. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 1527–1534. [Google Scholar] [CrossRef]
- Qamra, R.; Hubbard, S.R. Structural basis for the interaction of the adaptor protein grb14 with activated ras. PLoS ONE 2013, 8, e72473. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Warnecke, A.; Sandalova, T.; Achour, A.; Harris, R.A. PyTMs: A useful PyMOL plugin for modeling common post-translational modifications. BMC Bioinform. 2014, 15, 370. [Google Scholar] [CrossRef]
- Andrew, C.D.; Warwicker, J.; Jones, G.R.; Doig, A.J. Effect of phosphorylation on alpha-helix stability as a function of position. Biochemistry 2002, 41, 1897–1905. [Google Scholar] [CrossRef]
- Gupta, V.K.; Rajala, A.; Daly, R.J.; Rajala, R.V. Growth factor receptor-bound protein 14: A new modulator of photoreceptor-specific cyclic-nucleotide-gated channel. EMBO Rep. 2010, 11, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Flint, A.J.; Tiganis, T.; Barford, D.; Tonks, N.K. Development of “substrate-trapping” mutants to identify physiological substrates of protein tyrosine phosphatases. Proc. Natl. Acad. Sci. USA 1997, 94, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Myers, M.P.; Andersen, J.N.; Cheng, A.; Tremblay, M.L.; Horvath, C.M.; Parisien, J.P.; Salmeen, A.; Barford, D.; Tonks, N.K. TYK2 and JAK2 are substrates of protein-tyrosine phosphatase 1B. J. Biol. Chem. 2001, 276, 47771–47774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, A.; Dilly, A.K.; Rajala, R.V. Protein tyrosine phosphatase-1B regulates the tyrosine phosphorylation of the adapter Grb2-associated binder 1 (Gab1) in the retina. Cell Commun. Signal. 2013, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, R.V.; Wiskur, B.; Tanito, M.; Callegan, M.; Rajala, A. Diabetes reduces autophosphorylation of retinal insulin receptor and increases protein-tyrosine phosphatase-1B activity. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.; Singh, H.; Srivastava, H.K.; Singh, S.; Kishore, G.; Raghava, G.P.S. Benchmarking of different molecular docking methods for protein-peptide docking. BMC Bioinform. 2019, 19, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, R.V.; Chan, M.D.; Rajala, A. Lipid-protein interactions of growth factor receptor-bound protein 14 in insulin receptor signaling. Biochemistry 2005, 44, 15461–15471. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.E.; van Veen, T.; Ekstrom, P.A. Differential Akt activation in the photoreceptors of normal and rd1 mice. Cell Tissue Res. 2005, 320, 213–222. [Google Scholar] [CrossRef]
- Li, G.; Anderson, R.E.; Tomita, H.; Adler, R.; Liu, X.; Zack, D.J.; Rajala, R.V. Nonredundant role of Akt2 for neuroprotection of rod photoreceptor cells from light-induced cell death. J. Neurosci. 2007, 27, 203–211. [Google Scholar] [CrossRef]
- Ivanovic, I.; Anderson, R.E.; Le, Y.Z.; Fliesler, S.J.; Sherry, D.M.; Rajala, R.V. Deletion of the p85alpha regulatory subunit of phosphoinositide 3-kinase in cone photoreceptor cells results in cone photoreceptor degeneration. Invest. Ophthalmol. Vis. Sci. 2011, 52, 3775–3783. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S.S.; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotox. Res. 2019, 35, 775–795. [Google Scholar] [CrossRef]
- Wensel, T.G. Phosphoinositides in Retinal Function and Disease. Cells 2020, 9, 866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, A.; He, F.; Anderson, R.E.; Wensel, T.G.; Rajala, R.V.S. Loss of Class III Phosphoinositide 3-Kinase Vps34 Results in Cone Degeneration. Biology 2020, 9, 384. [Google Scholar] [CrossRef]
- He, F.; Agosto, M.A.; Anastassov, I.A.; Tse, D.Y.; Wu, S.M.; Wensel, T.G. Phosphatidylinositol-3-phosphate is light-regulated and essential for survival in retinal rods. Sci. Rep. 2016, 6, 26978. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Nichols, R.M.; Kailasam, L.; Wensel, T.G.; Agosto, M.A. Critical Role for Phosphatidylinositol-3 Kinase Vps34/PIK3C3 in ON-Bipolar Cells. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2861–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Agosto, M.A.; Nichols, R.M.; Wensel, T.G. Multiple phosphatidylinositol(3)phosphate roles in retinal pigment epithelium membrane recycling. bioRxiv 2020. [Google Scholar] [CrossRef]
- Van Epps, H.A.; Hayashi, M.; Lucast, L.; Stearns, G.W.; Hurley, J.B.; De Camilli, P.; Brockerhoff, S.E. The zebrafish nrc mutant reveals a role for the polyphosphoinositide phosphatase synaptojanin 1 in cone photoreceptor ribbon anchoring. J. Neurosci. 2004, 24, 8641–8650. [Google Scholar] [CrossRef] [Green Version]
- Prosseda, P.P.; Luo, N.; Wang, B.; Alvarado, J.A.; Hu, Y.; Sun, Y. Loss of OCRL increases ciliary PI(4,5)P(2) in Lowe oculocerebrorenal syndrome. J. Cell Sci. 2017, 130, 3447–3454. [Google Scholar] [CrossRef] [Green Version]
- Luo, N.; Conwell, M.D.; Chen, X.; Kettenhofen, C.I.; Westlake, C.J.; Cantor, L.B.; Wells, C.D.; Weinreb, R.N.; Corson, T.W.; Spandau, D.F.; et al. Primary cilia signaling mediates intraocular pressure sensation. Proc. Nat. Acad. Sci. USA 2014, 111, 12871–12876. [Google Scholar] [CrossRef] [Green Version]
- Travaglini, L.; Brancati, F.; Silhavy, J.; Iannicelli, M.; Nickerson, E.; Elkhartoufi, N.; Scott, E.; Spencer, E.; Gabriel, S.; Thomas, S.; et al. Phenotypic spectrum and prevalence of INPP5E mutations in Joubert syndrome and related disorders. Eur. J. Hum. Genet. 2013, 21, 1074–1078. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajala, R.V.S.; McCauley, A.; Rajala, R.; Teel, K.; Rajala, A. Regulation of Phosphoinositide Levels in the Retina by Protein Tyrosine Phosphatase 1B and Growth Factor Receptor-Bound Protein 14. Biomolecules 2021, 11, 602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11040602
Rajala RVS, McCauley A, Rajala R, Teel K, Rajala A. Regulation of Phosphoinositide Levels in the Retina by Protein Tyrosine Phosphatase 1B and Growth Factor Receptor-Bound Protein 14. Biomolecules. 2021; 11(4):602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11040602
Chicago/Turabian StyleRajala, Raju V. S., Austin McCauley, Rahul Rajala, Kenneth Teel, and Ammaji Rajala. 2021. "Regulation of Phosphoinositide Levels in the Retina by Protein Tyrosine Phosphatase 1B and Growth Factor Receptor-Bound Protein 14" Biomolecules 11, no. 4: 602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11040602