
Sialic Acids as Receptors for Pathogens
by
 ,
,
Patrycja Burzyńska
, 
Łukasz F. Sobala
,
Krzysztof Mikołajczyk
,
Marlena Jodłowska
and
Ewa Jaśkiewicz
* Laboratory of Glycobiology, Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, R. Weigla 12, 53-114 Wroclaw, Poland
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(6), 831; https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060831
Submission received: 10 May 2021
/
Revised: 28 May 2021
/
Accepted: 29 May 2021
/
Published: 2 June 2021
(This article belongs to the Section Molecular Medicine)
Abstract
:Carbohydrates have long been known to mediate intracellular interactions, whether within one organism or between different organisms. Sialic acids (Sias) are carbohydrates that usually occupy the terminal positions in longer carbohydrate chains, which makes them common recognition targets mediating these interactions. In this review, we summarize the knowledge about animal disease-causing agents such as viruses, bacteria and protozoa (including the malaria parasite Plasmodium falciparum) in which Sias play a role in infection biology. While Sias may promote binding of, e.g., influenza viruses and SV40, they act as decoys for betacoronaviruses. The presence of two common forms of Sias, Neu5Ac and Neu5Gc, is species-specific, and in humans, the enzyme converting Neu5Ac to Neu5Gc (CMAH, CMP-Neu5Ac hydroxylase) is lost, most likely due to adaptation to pathogen regimes; we discuss the research about the influence of malaria on this trait. In addition, we present data suggesting the CMAH gene was probably present in the ancestor of animals, shedding light on its glycobiology. We predict that a better understanding of the role of Sias in disease vectors would lead to more effective clinical interventions.
1. Introduction
Sialic acids (Sias) are the terminal carbohydrate moieties that occur on cell glycocalyx. They have been found in almost all forms of life, including humans and other mammals, but also in fungi, algae, bacteria and viruses [1,2], creating a host-specific “glycocalyx and sialome” on the cell surface. Most Sias are linked to glycoproteins, but in human neuronal cells, sialic acids are linked to sphingolipids (gangliosides) [3]. Sias mediate a wide variety of physiological and pathological processes, serving as ligands for cell adhesion molecules [4,5]. In fact, Sias are considered “self-associated molecular patterns” (SAMPS) cooperating with “self-pattern recognition receptors” (SPRR) [6]. Due to their negative charge, hydrophobicity and terminal position in sugar chains, they play an important role in cell stability and direct interactions with pathogens and toxins [4,5].
1.1. Structure of Sialic Acids
Sias are sugar [7] derivatives of neuraminic acid (nonulosonic acid [3]) (Figure 1a) usually present in glycoconjugates [8]. Neuraminic acid is chemically named 5-amino-3,5-dideoxy-d-glycero-d-galacto-non-2-ulopyranosonic acid (according to IUPAC 2020) and belongs to the 9-carbon α-keto carboxylic acids [3]. Three functional groups can be distinguished in Sias: a carboxyl group linked with the anomeric C-2 carbon (which making them ketoses), a three-carbon glycerol chain and an amino group linked to carbon C-5.
There are three major forms of neuraminic acid: 5-N-acetylneuraminic acid (Neu5Ac) (Figure 1c), 5-N-glycolylneuraminic acid (Neu5Gc) (Figure 1d) and 2-keto-deoxynonulosonic acid (Kdn) (Figure 1b), having a hydroxyl group at the C-5 position. In vertebrates, the formation of Sias is catalyzed by a bifunctional enzyme, glucosamine UDP-GlcNAc-2-epimerase/N-acetylmannosamine kinase (encoded by GNE gene), which converts UDP-GlcNAc (uridine diphosphate N-acetylglucosamine) to ManNAc-6-P (N-acetyl-mannosamine 6-phosphate) and UDP (uridine diphosphate) [9]. ManNAc-6-P is condensed with phosphoenolpyruvate (PEP), forming N-acetylneuraminate 9-phosphate (Neu5Ac-9-P), by Neu5Ac-9-P synthase. Then, Neu5Ac-9-P is dephosphorylated by a specific phosphatase, resulting in the release of Neu5Ac to the cytoplasm. In prokaryotes the condensation of ManNAc (N-acetylmannosamine) and PEP is catalyzed by an aldolase. The same pathway can use Man-6-P instead ManNAc-6-P to generate Kdn [3].
Sias can be modified by methylation, phosphorylation, sulfation and, most commonly, O-acetylation at C-4, C-7 or C-9. O-acetylated forms of sialic acids have been observed in bacteria, as well as eukaryotes (including humans) [10]. A neuraminic acid molecule may contain one to three O-acetyl residues, but N-acetyl-9-O-acetylneuraminic acid (Neu5,9Ac2) is the predominant form [11] (Figure 1). N-acetyl-4-O-acetylneuraminic acid (Neu5,4Ac2) was not found in humans, but it has gathered more interest recently as a receptor for pathogens infecting vertebrates [12,13]. More than 80 Sias have been reported in the literature [3,14].
The linkage configurations of Sias are α-2,3 or α-2,6 with galactose or N-acetylgalactosamine and α-2,8 or α-2,9 in polysialic acids. These bonds are created by specific sialyltransferases and polysialyltransferases, respectively [15]. The α-glycosidic bonds linking Sias to glycans are hydrolyzed by glycoside hydrolases named sialidases or neuraminidases (EC 3.2.1.18 [16]). In mammals, four sialidases (NEU1-NEU4) exist that differ in localization, specificity and function [17,18]. Many viruses produce sialidases, which increases their virulence or pathogenicity. Sialidases cleave Sias to enable the virus to attach to the host cell surface or to recognize the receptor by changing its structure or to uncover it. Transsialidases relocate Sias residues from host to pathogen cell surfaces to prevent its recognition by the host immune system. They may vary in substrate specificity, affecting the immune system or facilitating a release of progeny virions from cells [19,20,21]. Bacterial sialidases are treated as drug development targets [22,23].
1.2. Neu5Gc vs. Neu5Ac
The transformation of Neu5Ac into Neu5Gc occurs by hydroxylation of the methyl group in the acetyl moiety. This reaction is catalyzed by cytidine monophosphate-N-acetylneuraminic acid hydroxylase (CMAH) [24]. The CMAH gene was reported to be present in chordates, including lancelets, tunicates, reptiles and amphibians; most fish and a number of mammals but, also, in algae and bacteria [2]. Only a few deuterostome lineages do not have a functional CMAH gene: among mammals, the CMAH gene is nonfunctional in humans, New World monkeys, the European hedgehog, musteloids, several bats, the sperm whale, white-tailed deer and the platypus. Thus, these organisms cannot synthesize Neu5Gc, which may influence their susceptibility to parasites. According to previous analyses [2,25], CMAH was acquired by a deuterostome via lateral gene transfer from a microalga. Notwithstanding the question of deuterostome monophyly [26], we performed BLAST searches using an extensive database of eukaryotic protein sequences and recovered a full sequence of putatively functional CMAH from a sponge Oscarella pearsei and partial sequences from sponges Oscarella carmela and Corticium candelabrum. All of these sponges belong in the class Homoscleromorpha; no CMAH sequences were detected in any of the three other sponge classes. Phylogenetic trees prepared in IQTree 2 using representative animal and nonanimal CMAH protein sequences showed that sponge CMAH belonged within a sister group to other animal CMAH sequences, while nonanimal CMAH sequences were basal to them (Figure 2). The placement of sponge CMAH within this sister clade had full bootstrap support. It follows that the alga-to-deuterostome transfer hypothesis should probably be revised; the gene seems to have appeared in animals earlier. We did not detect CMAH in any choanoflagellates (the sister group to animals) or other nonanimal holozoan species but only in protists even less closely related to animals (Figure 2). If we assume sponges are the earliest known branch of animals [27], CMAH might have been one of the genetic novelties present in the lineage that led to animals, present in the last common ancestor of animals but not in the (earlier) last unicellular ancestor of animals [28]. We posit that Neu5Gc presence might have shaped the relationships between animals and their pathogens since the Metazoa kingdom evolved.
In comparison with other mammals, the human CMAHP pseudogene lacks exon 3 [33] (Figure 3). Two to three million years ago, a deletion of 92 bp changed the open reading frame (ORF) to cause a premature STOP codon, resulting in a 72-aa-long truncated protein [34,35]. The longest current human CMAHP ORF encodes a 247-aa protein containing only the C-terminal β-lactamase-like domain (Figure 3). Even if this protein was produced, the lack of the N-terminal Rieske domain erases CMAH activity. The full ancestral human CMAH protein was 590 aa long, identically to the chimpanzee enzyme. Inactivation of the CMAH gene in animals occurred many times in independent lineages. For example, in New World monkeys, separate events caused an inversion of exons 4-13, a deletion of exons 4-8 and a deletion of exons 10-13. The platypus harbors a premature stop codon in exon 5, and musteloids lost nine coding exons. Species with a nonfunctional CMAH gene only express Neu5Ac on the surface of their cells [2,36].
According to the catastrophic selection hypothesis, a factor related to CMAH enzyme loss might be a lethal virus recognizing Neu5Gc [37]. One pathogen that could play such a role might be the malaria parasite Plasmodium, for which Neu5Gc was an ancestral receptor [38]. Others speculated that anti-Neu5Gc antibodies made by CMAH -/- females could eliminate sperms from CMAH-positive males, leading to a depletion of functional CMAH in the population [39].
Although Neu5Gc is not produced by humans, its trace amounts can be detected on the surface of endothelial and epithelial cells, as well as in fetal tissues. The source of Neu5Gc is a diet rich in red meat and dairy products [40]. It is absorbed by micropinocytosis and subsequently incorporated into cells [41]. Moreover, Neu5Gc was also found in Heamophilus influenzae, a human commensal that can cause respiratory infections and meningitis. H. influenzae is also unable to synthesize Neu5Gc, but it can absorb the molecule from the diet of its host and incorporate this Sia into its surface oligosaccharides [42].
1.3. Interactions with Pathogens
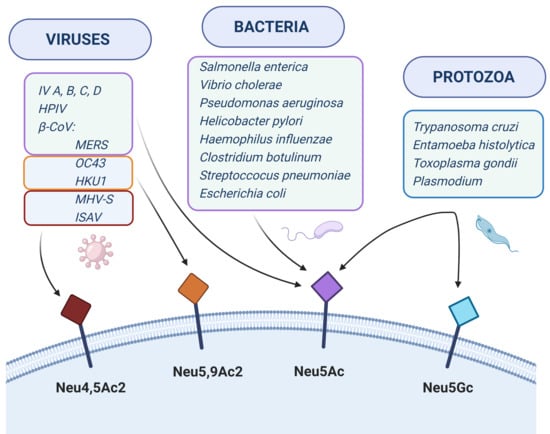
Sialic acids can act as receptors for various pathogens underlying many infectious diseases (Table 1). For example, viruses can use Sias to attach to and enter cells, bacteria produce adhesins or toxins interacting with Sias on host cells and protozoa can use Sias to evade host immunity. In this review, we describe the known molecular interactions of Sias with various pathogens. For more detailed information, we refer to recent reviews in these topics [43,44,45,46,47,48,49].
2. Viruses
2.1. Influenza Virus
There are a several types of influenza viruses, all belonging to Orthomyxoviridae family. Influenza virus type A (IAV) causes seasonal disease in humans, but it was also responsible for the 1918 Spanish flu and 2009 swine flu pandemics [89]. In addition to humans, it can infect other animal species, including pigs, horses and birds. Influenza virus type B (IBV) causes almost the same disease symptoms as IAV, but although it is able to colonize animals (dogs or grey seals), it infects mainly humans and does not have pandemic potential [50]. Influenza virus type C (ICV) causes mild flu symptoms. The main reservoir of ICV are humans, but it is able to infect pigs as well. Influenza virus type D (IDV), whose reservoir is cattle, is harmless for humans [53].
The antigenic proteins that participate in infection of influenza viruses comprise of hemagglutinins (HA), neuraminidases (NA) [90] and the hemagglutinin esterase fusion protein (HEF) [54]. Sialic acids are receptors for all types of influenza viruses, which recognize them using HA. Both the type and the linkage of a sialic acid affect the infectivity of influenza viruses in a species-specific manner [51]. Avian IAV binds to α-2,3-linked Neu5Ac, which is present in the human lower respiratory tract, while human IAV binds mainly α-2,6-linked Sias, present in the upper respiratory tract. Thus, the location of infection depends on the binding preferences of HA. The binding depends on the length of the surface glycans, as well as their density of sialylation [91]. IBV infects mainly human upper respiratory tract, but some strains are able to infect the lower respiratory tract. Thus, it seems IBV also prefers α-2,6-linked Sias. NAs remove sialic acids, promoting viral attachment to cell surface and then its internalization by endocytosis [92]. It was found that some IAV strains can bind to Neu5Gc [93] on human epithelial cells, which may be incorporated from diet, but it does not work as a functional receptor. It was suggested that Neu5Gc may act as a decoy towards IAV: it is not susceptible to the viral sialidase, which makes endocytosis impossible. As a result, this binding suppresses infection in humans.
The O-acetyl form of Neu5Ac is the preferred receptor for ICV. The major player in ICV infection is hemagglutinin-esterase-fusion protein (HEF), which binds to Neu5,9Ac2 (α-2,3 or α-2,6 linkage) and removes the 9-O-acetyl group from the receptor using esterase activity. HEF catalyzes subsequent viral fusion with host cell membrane [46,53]. ICV and IDV HEF, used for infection, are structurally very similar [53], as well as the Infectious Salmon Anemia Virus (ISAV) HEF protein, which is specific to Neu5,4Ac2 and is not contagious for humans [12,13,94].
2.2. SV40
Simian virus 40 (SV40) belongs to polyomaviruses, and its primary reservoir are monkeys. People can be infected through aerosols or ingestion. SV40 was found to contaminate the polio vaccines, which were produced in 1950s and 1960s in monkey cells [95]. The virus has long been known to cause cancer in some species, including humans, most often mesothelioma and brain tumors. Cells in these tissues are nonpermissive to SV40 replication, but the virus can integrate into them and cause malignant transformation instead of cell lysis [96].
The natural SV40 receptor is GM1 ganglioside, a monosialotetrahexosyl glycosphingolipid. Upon binding of GM1 to the binding pockets formed by pentamers of viral capsid protein VP1, cholesterol-dependent endocytosis occurs [97]. It was found that SV40 binds to Neu5Gc-GM1 much tighter than to Neu5Ac-GM1 [98], which reduces its infectivity towards human cells [99].
2.3. Coronaviruses
Coronaviruses cause infections of respiratory and gastrointestinal tracts. They are classified into four genera: alpha-, beta-, gamma- and deltacoronaviruses [100]. All coronaviruses are able to infect mammals, but gamma- and deltacoronaviruses can also infect avian species. The coronaviruses most pathogenic for humans, the severe acute respiratory syndrome coronavirus (SARS-CoV) and Middle East respiratory syndrome coronavirus (MERS-CoV) are all betacoronaviruses. SARS-CoV-2, responsible for the 2019-21 pandemic [101], is more closely related to SARS-CoV than MERS-CoV and also belongs to the betacoronavirus genus. The main receptor for SARS-CoV viruses is angiotensin-converting enzyme 2 (ACE2), and for MERS-CoV, it is dipeptidyl peptidase 4 (DPP4 or CD26). In addition to DPP4, MERS-CoV recognizes sialic acids. It was shown that MERS-CoV Spike (S) protein preferentially binds to α-2,3-Neu5Ac, while its binding to α-2,6-Neu5Ac, Neu5,9Ac2 and Neu5Gc was low or nonexistent [56]. While sialic acid facilitates MERS-CoV infection, in the case of SARS-CoV and SARS-CoV-2, the presence of sialic acids seems to restrict the infection. It was shown that the NA treatment of lung epithelial cells significantly promoted the replication and infection of both viruses, wherein SARS-CoV-2 was affected to a lesser extent. These results suggested a precluding role of ACE2 receptor Sias for SARS-CoV -2 binding. The increased SARS-CoV-2 S protein binding to ACE2, treated by NA, indicates a different role of Sias on SARS-CoV-2 binding and infection [102]. Thus, viral attachment likely occurs in a two-step fashion; the first low affinity Sias binding is followed by the higher affinity protein receptor binding [103].
Other human betacoronaviruses, OC43 and HKU1, recognize 9-O-acetylated sialic acid [55,104]. These viruses produce the sialate-O-acetylesterase protein (HE), which contains an O-acetylated sialic acids-binding domain and sialate O-acetylesterase domain. It was suggested that the HE protein evolved from the ICV HEF protein, but its binding activity is lost. This enzyme derivatizes O-acetylated sialic acids at a later stage of infection to prevent irreversible binding of progeny virions to the 9-O-acetylated Sias decoys. Mouse Hepatitis Virus (MHV) is another betacoronavirus producing HE protein similar to ICV HEF, but like ISAV, it recognizes Neu5,4Ac2 and is not able to infect humans [12,13,105,106].
2.4. Human Parainfluenza Viruses
Human parainfluenza viruses (HPIVs) are common respiratory pathogens belonging to the Paramyxoviridae family. They are divided into four serotypes (HPIV-1–HPIV-4), the last of which has two major subtypes (HPIV-4A and HPIV-4B) [107]. HPIVs cause lower and upper respiratory tract infections, mainly in newborns and young children, but also in elderly and immunocompromised patients. The most common symptoms include cold-like symptoms, tracheobronchitis, bronchiolitis, laryngotracheobronchitis and pneumonia. HPIV infections are associated with a wide range of neurological and cardiovascular illnesses, i.e., febrile seizures, meningoencephalitis, myocarditis, pericarditis and bradycardia [108].
One of the six essential proteins encoded by HPIV RNA is a membrane-anchored hemagglutinin neuraminidase (HN), which uses sialic acid as a receptor in host cell entry. The HN glycoprotein is also involved in the budding and release of new virions from the infected cell by Sias residue cleavage [109]. The viral HN binds predominantly to oligosaccharides terminated with α-2,3-linked Neu5Ac, but it seems that HPIV-1 HN can cleave α-2,8-linked Sias from α-2,3-linked Sias without binding to it [57]. Moreover, HPIV-3 HN shows ability to bind to α-2,6-linked Neu5Ac and α-2,6-linked Neu5Gc [58,59].
3. Bacteria
3.1. Salmonella enterica
Salmonella enterica is a Gram-negative, rod-shaped bacterium. The species is divided into six subspecies and over 2600 serovars [110]. One of them, Salmonella Typhi, is a bacterium causing typhoid fever in humans with such symptoms as fever, headache, rash and abdominal pain. Its toxin belongs to the two-subunit AB5 six-component toxin type, unique seven-component structure (A2B5). The A subunits have enzymatic activity: deoxyribonuclease CdtB and ADP ribosyltransferase PltA, while the B subunits (PltB) bind to glycoprotein receptors with terminal Neu5Ac but not Neu5Gc [60]. The expression of the highly immunogenic S. Typhi toxin is substantially upregulated after bacterial entry into the host cells [111]. In species that express Neu5Gc, S. Typhi is unable to multiply. The only exception is the chimpanzee, in which S. Typhi was identified, but no disease symptoms have been observed.
Another example is Salmonella Typhimurium that causes bacteremias and acute gastroenteritis, with symptoms such as diarrhea [112,113]. Its toxin ArtAB, which is similar to S. Typhi toxin, is an important virulence factor causing symptoms, which are an effect of its immunogenicity that triggers the immune system response [114]. It was shown that Neu5Ac is a potential receptor for S. Typhimurium [61]. Moreover, S. Typhimurium LT2 sialidase prefers α-2,3-linked over α-2,6-linked sialic acids, and its ability to cleave Neu5Gc is low [115].
3.2. Vibrio cholerae
Cholera, caused by Vibrio cholerae, is a disease specific only to humans [116], whose symptoms include diarrhea, vomiting and muscle cramps. The bacterium has three cytotoxicity factors: TCP (toxin-coregulated pilus) responsible for the colonization of small intestine, CTx (cholera toxin) and VcN (V. cholerae neuraminidase) [62,117]. CTx belongs to the AB5 family and recognizes monosialylated Neu5Ac-GM1 or Neu5Gc-GM1. However, the majority of gangliosides on the surface of human epithelial cells are di- and trisialo-derivatives. V. cholerae uses a sialidase to hydrolyze α-2,3- and α-2,8-linked sialic acids, turning them into GM1. It was shown that Neu5Gc bound to GM1 is less susceptible to enzymatic hydrolysis in comparison to Neu5Ac. Thus, it seems that the species adapted to the human-specific loss of Neu5Gc, which underlies our susceptibility to this pathogen.
3.3. Pseudomonas aeruginosa
Pseudomonas aeruginosa is a Gram-negative, rod-shaped bacteria, often found in plants, soil and water environments and, sometimes, in human bodies. P. aeruginosa is an opportunistic pathogen with natural resistance to antibiotics [118,119], causing hospital-acquired infections, e.g., pneumonia, meningoencephalitis and sepsis [63]. It infects mostly the respiratory and urinary tracts, blood, burn wounds and the outer ear. Its ability to form biofilms, combined with quorum sensing mechanisms [120], is associated with extreme drug tolerance [121].
The main virulence factors are the single flagellum, pili and components of the outer plasma membrane that participate in adhesion to the host airway epithelia [122]. Moreover, P. aeruginosa produces toxins: pyocyanin [123], lipopolysaccharide and exotoxin A [124]. The flagellum interacts with Toll-like receptors TLR-5 and TLR-2 [125,126], mucins [127] and heparan sulfate proteoglycans [122,128], while the pili bind to N-glycans [129]. Furthermore, P. aeruginosa interacts with laminin [130,131] and integrins [132,133,134]. Both the pili and the flagellum recognize asialo-monogangliosides (GM1 or GM2), which expose a GalNAcβ1-4Gal moiety [64]. Such binding promotes an inflammatory response [126], which is especially threatening to individuals with cystic fibrosis because of an increase of asialo-GM1 expression on the surfaces of airway cells [135]. Another receptor of P. aeruginosa is the sialyl-Lewis x antigen with terminal α-2,3-Neu5Ac [65], and its importance in the context of cystic fibrosis is similar.
3.4. Helicobacter pylori
Helicobacter pylori is a helical-shaped Gram-negative bacterium able to survive in extreme acidic conditions; hence, its niche is usually the stomach lining. It is a common cause of ulcers and gastritis [136], which, untreated, can lead to gastric cancer [137]. H. pylori has flagella, which are necessary for bacterial motility in gastric mucus [138]. The bacterium produces Ni2+-dependent urease [139], which hydrolyzes urea into ammonia to neutralize the acidity and create an optimal environment to survive in the stomach. Other virulence factors of H. pylori are adhesins, some of which recognize sialic acids. Adhesin SabA prefers N-acetyllactosamine-based gangliosides with terminal α-2,3-linked Neu5Ac (including the sLexantigen) over α-2,6-linked Neu5Ac and terminal Neu5Gc [66,67,68,69]. Protein HpaA is an adhesin potentially capable of binding sialic acids [70,71], but its specificity is not known yet.
3.5. Haemophilus influenzae
Haemophilus influenzae is an opportunistic Gram-negative coccobacillus bacterium. Depending on its ability to form polysaccharide capsules, its strains may be divided to typeable (encapsulated) and non-typeable (unencapsulated) [140]. The encapsulated strains are categorized into six serotypes (from a to f), of which serotype b is considered the most virulent [141]. H. influenzae mainly causes respiratory tract infections, especially in children and the elderly, chronic obstructive pulmonary disease in cystic fibrosis patients [142,143], sinusitis [144] and pneumonia. It can also cause conjunctivitis [145] and otitis media (middle ear inflammation) [146].
H. influenzae strains have various factors participating in infection, playing a role in interactions with host cells [147,148,149,150,151]. They include adhesins [152,153] and other components anchored to its outer membrane: pili [154,155], IgA proteases [156] and lipooligosaccharide (LOS) [157]. It was shown that H. influenzae contains α-2,6-Neu5Ac [73], but the bacterium is not able to synthesize Sias and, instead, uses those absorbed from its environment [74]. Dietary Neu5Gc can be also incorporated into the LOS, but Neu5Ac is up to four times more favored [72]. It was suggested that H. influenzae mimics the sialylation pattern of host mucin [73].
3.6. Clostridium botulinum
Clostridium botulinum is a rod-shaped, Gram-positive bacterium with an ability to sporulate [158]. It is an obligate anaerobe thar lives in soil, dust and sediments, but it is also able to infect animals (including humans) through food. Botulism is a disease caused by botulinum neurotoxins. Its main symptom is a descending flaccid paralysis, which, if left untreated, can result in death.
Botulinum neurotoxins are metalloproteases that specifically cleave SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) proteins in neurons, which results in the inhibition of neurotransmission [159]. Based on its antigenic properties and amino acid sequence, the botulinum toxins are classified into seven subtypes (A–G). Subtypes A, B, E and (rarely) F cause botulism in humans [160], while subtypes C and G affect birds and other animals. It was shown that types A-D and G toxins associate in complexes with hemagglutinins (HA) [75,76,77], which enables them to bind to gangliosides with terminal sialic acid. In some cases, such binding is critical for infectivity [78,161]. It is known that complexes of botulinum toxin and HA recognize Neu5Ac, but whether this happens with Neu5Gc as well is not known.
3.7. Streptoccocus pneumoniae
Streptococcus pneumoniae, also known as pneumococcus, is a commensal, Gram-positive, spherical bacterium. It may, however, cause pneumonia and other infections such as otitis media, sinusitis, meningitis and bronchitis [162,163], especially in children, the elderly and in patients with immunodeficiency. S. pneumoaniae produces various virulence factors, including surface enzymes, polysaccharide capsules, pili, choline-binding proteins, lipoproteins and toxin pneumolysin [164], as well as sialidases. These glycoside hydrolases can cleave the T-antigen, a mucin disaccharide normally covered by a terminal sialic acid, and are specific to both α-2,3- and α-2,6-linked Neu5Ac. The T-antigen is processed to galactose and N-acetylgalactosamine [80], which elicits an IgM response, leading to pneumococcal hemolytic uremic syndrome (HUS). Additionally, it was shown that Neu5Ac is preferred over Neu5Gc [79].
3.8. Escherichia coli
Escherichia coli is a species of Gram-negative, rod-shaped, commensal bacteria naturally occurring in the colonic microbiota of humans and animals, transmitted by water and sediments [165]. Its vast genetic diversity causes differences in its pathogenicity [166]. The disease-causing strains mainly infect the gastrointestinal and the urinary tracts. The major virulence factors of E. coli are pili [167], K- and O-antigens [168], adhesins [169], lipopolysaccharide [170], hemolysin [171] and toxins. The subtilase cytotoxin (SubAB) of Shiga toxin-producing E. coli (STEC) can damage the internal organs, and the infection caused by it is characterized by acute kidney failure, thrombocytopenia and microangiopathic hemolytic anemia [82]. SubAB belongs to AB5 toxins and binds to receptors by pentameric B subunits. The receptors are sialylated glycolipids with terminal α-2,3-Neu5Gc, to which the affinity is 20 times higher than to α-2,3-Neu5Ac and 30 times higher than to α-2,6-Neu5Gc [81].
4. Protozoa
4.1. Trypanosoma cruzi
Trypanosoma cruzi is a parasite causing Chagas disease, affecting mainly Latin America but spreading worldwide due to human migration and, also, the specific way of pathogen transmission. The vectors of T. cruzi are certain species of three genera of blood-sucking triatomine insects: Triatoma, Panstrongylus and Rhadonius, also called “kissing bugs”. Other possible routes of infection are: mother-to-child transmission, blood and blood products or contaminated food and drink. The major disease symptoms involve cardiac and digestive manifestations [172]. A protein important in T. cruzi infection is gp85—a transsialidase (TcTS) [173] that hydrolyses α-linked Sias present on the host cell surface and transfers them to mucin molecules on the parasite cell surface. Such a mechanism prevents pathogen recognition by the host immune system [174]. TcTS is specific to α-2,3-linked Sias on the donor cells, and it recreates the same linkage on the pathogen cells, where the acceptor is a terminal β-galactopyranosyl group. TcTS is able to transfer both Neu5Ac and Neu5Gc [175].
4.2. Entamoeba histolytica
Entamoeba histolytica is an anaerobic amoebozoan [176] belonging to the Entamoebidae family (Archamoebae). As a human pathogen, it causes amoebiasis with symptoms related to the gastrointestinal tract, with possible complications that include liver, lung or brain abscesses [177]. Foodborne infection occurs by ingestion food or water contaminated with E. histolytica cysts. Their cell wall structures allow them to survive in the acidic environment of the host stomach. In the small intestine, the cysts release motile trophozoites that colonize the colon and then differentiate into cysts that are excreted in the feces [178,179]. It was shown that trophozoites bind to host mucins in a sialic acid-dependent manner [87]. A transmembrane sialidase on the E. histolytica cell surface [180] uncovers the receptor before it can be recognized by the E. histolytica toxin. The sialidase substrates are α-2,3-Neu5Ac and colonic acid (a homopolymer of α-2,8-Neu5Ac), but α-2,6-Neu5Ac is not hydrolyzed [181].
4.3. Toxoplasma gondii
Toxoplasma gondii is a parasitic protozoan from phylum Apicomplexa that is able to infect humans and other warm-blooded animals. It is dangerous especially for immunocompromised individuals and developing fetuses. T. gondii encysts and establishes chronic infections in the brain, the heart and skeletal muscles [182,183]. While its life cycle contains both sexual and asexual phases, the only host where it can reproduce sexually is the cat. Inside the feline gastrointestinal tract, T. gondii differentiates into gametocytes that are then excreted in the feces. One of the major virulence factor of T. gondii are proteins produced by micronemes (apicomplexan parasites secretory organelles), and they recognize sugar chains on the host cell surface; thus, Sias are important molecules involved in infections. Toxoplasma microneme protein 1 (TgMIC1) has a high affinity for terminal α-2,3-linked Sias and a low affinity for α-2,6-linked Sias [184]. Another protein, Toxoplasma microneme protein 13 (TgMIC13), shows an affinity for 4-O-acetylated Sias and the less well-known α-2,9-linked Sias [185]. Recently, it was shown that TgTCP-1 chaperonin [186] and tachyzoite surface Sia-binding protein 1 (TgSABP1) interact with Sias on the host cell surface, and sialic acid ablation by sialidase affected the interaction [187].
4.4. Plasmodium
The malaria parasites from the genus Plasmodium also take advantage of the ubiquity of Sias on the surface on red blood cells to invade their hosts. Plasmodium falciparum is responsible for infecting humans, while African apes harbor at least six species of Plasmodium, three of which primarily infect chimpanzees (P. reichenowi, P. gaboni and P. billcollinsi), while the other three infect gorillas (P. praefalciparum, P. blacklocki and P. adleri) [188,189].
Several proteins expressed by Plasmodium merozoites are involved in the erythrocyte invasion, including two multigene families: erythrocyte-binding ligands (EBL) and reticulocyte-binding ligands (RBL) [190,191]. Four functional P. falciparum EBL proteins were identified so far [192]: erythrocyte-binding antigen-175 (EBA-175), erythrocyte-binding antigen-181 (EBA-181), erythrocyte-binding ligand-1 (EBL-1) and erythrocyte-binding antigen-140 (EBA-140). The well-studied P. falciparum EBA-175 merozoite ligand [83] recognizes α-2,3-linked Sias presented on clusters of O-linked glycans attached to erythrocyte surface glycophorin A (GPA) [193,194]. The EBA-140 homologous merozoite ligand [195,196,197] was shown to bind glycophorin C (GPC) [84,198,199,200], which is a minor erythrocyte sialoglycoprotein [201]. It was proposed that the receptor for EBA-140 ligand is a cluster of N- and O-linked sialylated glycans on the GPC molecule [202,203] (Figure 4). Thus, Sias seem to be necessary for the recognition of both EBA receptors [204,205,206,207].
Homologs of the PfEBL merozoite ligands were identified in ape parasites, including P. reichenowi [208]. Expression of P. falciparum and P. reichenowi EBA-175 and EBA-140 on the surface of African green monkey (COS7) cells showed some Sias host-specific preferences for erythrocyte binding [209]. The preferred ligand for P. falciparum EBA-175 and EBA-140 was Neu5Ac, while P. reichenowi proteins bound preferentially to Neu5Gc. Thus, it has been argued that the expression of different Sias is a primary factor responsible for the species-specific binding of human and ape parasites [39,209].
Further studies on the P. falciparum EBA-175 ligand specificity confirmed that it associates with GPA only when Neu5Ac linked to α-2,3-Gal is present [210]. However, the strong binding of EBA-175 to Neu5Gc monosaccharide was also observed.
In another approach, human ex vivo cultured erythrocytes (cRBCs) were modified by introducing Neu5Gc on the cell surface through the expression of chimpanzee CMAH [211]. The presence of Neu5Gc on human cRBCs significantly increased P. knowlesi merozoite invasion. However, in contrast to P. knowlesi, which is a parasite of macaques, the human parasite P. falciparum invaded normal and modified cRBCs at a similar level, which suggested that P. falciparum can utilize both Neu5Ac and Neu5Gc. This result supported the bespoke hypothesis that PfEBA-175, the major sialic acid-dependent invasion ligand, does not discriminate between Neu5Ac and Neu5Gc.
EBA-165, another homologous merozoite ligand dependent on Sias binding, is present in all ape-infective Plasmodium species, but due to a mutation causing a frameshift, it is not present in P. falciparum [212]. Consistent with the previous findings [211], it was confirmed that the PfEBA-175 ligand was able to bind to glycans containing Neu5Ac and Neu5Gc. By contrast, P. reichenowi EBA-165 and the full-length, corrected P. falciparum EBA-165 (cP.f. EBA-165) bound only to glycans containing Neu5Gc.
In summary, there is clear preference for Neu5Gc in all studied Plasmodium ape species (P. reichenowi and P. knowlesi), while the human parasite P. falciparum shows a specificity towards both Neu5Gc and Neu5Ac. Consequently, the gain/loss of function mechanism may serve as a host switch for P. falciparum, because P. falciparum has gained the ability to bind Neu5Ac without losing the binding specificity for Neu5Ac. It might have implications for the emergence of P. falciparum as a major deadly human pathogen. While the EBA Sias preference hypothesis is intriguing, it may not represent the complete story of Plasmodium merozoites specificity.
5. Conclusions
The glycocalyx of human cells differs from many other mammals by the lack of the Neu5Gc and the presence of its precursor Neu5Ac. Humans are not alone in this loss. Several other species of mammals have independently developed different loss-of-function mutation in the CMAH gene. This convergent evolution of the vertebrate-specific Neu5Gc-containing glycans represents an evolutionary change in the use of Sias as receptors for pathogens. A consequence of this change was the change in pathogen regimes; thus, pathogens that can bind Neu5Ac would have a preference for humans. Indeed, a number of human-specific pathogens evolved a specificity for Neu5Ac. These include most influenza viruses, bacteria and the human malaria parasite P. falciparum. Presumably, pathogens that require Neu5Ac could differentially expand in humans. Thus, a better understanding of human infectious disease mechanisms based on Sias would allow for new drug designs; for example, the Sias structural analogs currently used as specific inhibitors in influenza virus infection treatment.
Author Contributions
Conceptualization, E.J., P.B. and Ł.F.S.; writing—original draft preparation, P.B., E.J., and Ł.F.S.; writing—review and editing, P.B., Ł.F.S., K.M. and E.J.; figures and table preparation, P.B., K.M., M.J. and Ł.F.S.; supervision, E.J. and funding acquisition, E.J. and Ł.F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by National Science Centre of Poland Preludium project (A.Z.) 2016/23/N/NZ6/01482, Harmonia project (E.J.) 2018/30/M/NZ6/00653 and Sonatina project (Ł.F.S.) 2020/36/C/NZ8/00081.
Data Availability Statement
The data used to generate the phylogenetic trees, as well as all the CMAH protein sequences considered in this review, are available on Zenodo under DOI: https://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.4585495.
Acknowledgments
Ł.F.S. would like to thank the Multicellgenome lab for access to the protein sequence data. All the authors would like to thank Marcin Czerwiński for his valuable comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schauer, R. Sialic acids: Fascinating sugars in higher animals and man. Zoology 2004, 107, 49–64. [Google Scholar] [CrossRef]
- Peri, S.; Kulkarni, A.; Feyertag, F.; Berninsone, P.M.; Alvarez-Ponce, D. Phylogenetic Distribution of CMP-Neu5Ac Hydroxylase (CMAH), the Enzyme Synthetizing the Proinflammatory Human Xenoantigen Neu5Gc. Genome Biol. Evol. 2018, 10, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Schauer, R.; Kamerling, J.P. Exploration of the Sialic Acid World. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Curr. Opin. Struct. Biol. 2009, 19, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Padler-Karavani, V. Evolution of sialic acids: Implications in xenotransplant biology. Xenotransplantation 2018, 25, e12424. [Google Scholar] [CrossRef] [PubMed]
- Skarbek, K.; Milewska, M.J. Biosynthetic and synthetic access to amino sugars. Carbohydr. Res. 2016, 434, 44–71. [Google Scholar] [CrossRef]
- Wang, B.; Brand-Miller, J. The role and potential of sialic acid in human nutrition. Eur. J. Clin. Nutr. 2003, 57, 1351–1369. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Cummings, R.; Esko, J.; Freeze, H.; Stanley, P.; Bertozzi, C.; Hart, G.; Etzler, M. Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017. [Google Scholar]
- Nguyen, T.; Lee, S.; Yang, Y.-A.; Ahn, C.; Sim, J.H.; Kei, T.G.; Barnard, K.N.; Yu, H.; Millano, S.K.; Chen, X.; et al. The role of 9-O-acetylated glycan receptor moieties in the typhoid toxin binding and intoxication. PLoS Pathog. 2020, 16, e1008336. [Google Scholar] [CrossRef] [Green Version]
- Schauer, R.; Srinivasan, G.V.; Wipfler, D.; Kniep, B.; Schwartz-Albiez, R. O-Acetylated Sialic Acids and Their Role in Immune Defense. Adv. Exp. Med. Biol. 2011, 705, 525–548. [Google Scholar] [CrossRef]
- Park, S.S. Post-Glycosylation Modification of Sialic Acid and Its Role in Virus Pathogenesis. Vaccinces 2019, 7, 171. [Google Scholar] [CrossRef] [Green Version]
- Wasik, B.R.; Barnard, K.N.; Parrish, C.R. Effect of Sialic Acid Modifications on Virus Binding and Infection. Trends Microbiol. 2016, 24, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.; La Rocca, P.; Allevi, P.; Pappone, C.; Anastasia, L. Intramolecular Lactones of Sialic Acids. Int. J. Mol. Sci. 2020, 21, 8098. [Google Scholar] [CrossRef] [PubMed]
- Mikolajczyk, K.; Kaczmarek, R.; Czerwinski, M. How glycosylation affects glycosylation: The role of N-glycans in glycosyltransferase activity. Glycobiology 2020, 30, 941–969. [Google Scholar] [CrossRef] [PubMed]
- Carbohydrate-Active enZYmes Database. Available online: http://www.cazy.org (accessed on 1 February 2021).
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhorukov, V.N.; Karagodin, V.P.; Zakiev, E.R.; Grechko, A.V.; Orekhov, A.N. Sialidases: Therapeutic and Antiatherogenic Potential. Curr. Pharm. Des. 2017, 23, 4696–4701. [Google Scholar] [CrossRef]
- Kim, S.; Oh, D.B.; Kang, H.A.; Kwon, O. Features and applications of bacterial sialidases. Appl. Microbiol. Biotechnol. 2011, 91, 1–15. [Google Scholar] [CrossRef]
- Air, G.M. Influenza neuraminidase. Influ. Other Respir. Viruses 2012, 6, 245–256. [Google Scholar] [CrossRef]
- Lipnicanova, S.; Chmelova, D.; Ondrejovic, M.; Frecer, V.; Miertus, S. Diversity of sialidases found in the human body—A review. Int. J. Biol. Macromol. 2020, 148, 857–868. [Google Scholar] [CrossRef]
- Glanz, V.Y.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Inhibition of sialidase activity as a therapeutic approach. Drug Des. Dev. Ther. 2018, 12, 3431–3437. [Google Scholar] [CrossRef] [Green Version]
- Shie, J.J.; Fang, J.M. Development of effective anti-influenza drugs: Congeners and conjugates—A review. J. Biomed. Sci. 2019, 26. [Google Scholar] [CrossRef] [Green Version]
- Shaw, L.; Schauer, R. The Biosynthesis of N-Glycoloylneuraminic Acid Occurs by Hydroxylation of the CMP-Glycoside of N-Acetylneuraminic Acid. Biol. Chem. Hoppe-Seyler 1988, 369, 477–486. [Google Scholar] [CrossRef]
- Simakov, O.; Kawashima, T.; Marlétaz, F.; Jenkins, J.; Koyanagi, R.; Mitros, T.; Hisata, K.; Bredeson, J.; Shoguchi, E.; Gyoja, F.; et al. Hemichordate genomes and deuterostome origins. Nature 2015, 527, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Kapli, P.; Natsidis, P.; Leite, D.J.; Fursman, M.; Jeffrie, N.; Rahman, I.A.; Philippe, H.; Copley, R.R.; Telford, M.J. Lack of support for Deuterostomia prompts reinterpretation of the first Bilateria. Sci. Adv. 2021, 7, eabe2741. [Google Scholar] [CrossRef]
- Kapli, P.; Telford, M.J. Topology-dependent asymmetry in systematic errors affects phylogenetic placement of Ctenophora and Xenacoelomorpha. Sci. Adv. 2020, 6, eabc5162. [Google Scholar] [CrossRef]
- Ros-Rocher, N.; Pérez-Posada, A.; Leger, M.M.; Ruiz-Trillo, I. The origin of animals: An ancestral reconstruction of the unicellular-to-multicellular transition. Open Biol. 2021, 11, 200359. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geneious 8.1.9. Available online: https://www.geneious.com (accessed on 4 March 2021).
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, T.; Aki, I.; Varki, A.; Satta, Y.; Takahata, N. Fixation of the Human-Specific CMP-N-Acetylneuraminic Acid Hydroxylase Pseudogene and Implications of Haplotype Diversity for Human Evolution. Genetics 2006, 172, 1139–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, H.-H.; Takematsu, H.; Diaz, S.; Iber, J.; Nickerson, E.; Wright, K.L.; Muchmore, E.A.; Nelson, D.L.; Warren, S.T.; Varki, A. A mutation in human CMP-sialic acid hydroxylase occurred after the Homo-Pan divergence. Proc. Natl. Acad. Sci. USA 1998, 95, 11751–11756. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.-H.; Hayakawa, T.; Diaz, S.; Krings, M.; Indriati, E.; Leakey, M.; Paabo, S.; Satta, Y.; Takahata, N.; Varki, A. Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 11736–11741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, M.O.; Gagneux, P. Absence of Neu5Gc and Presence of Anti-Neu5Gc Antibodies in Humans—An Evolutionary Perspective. Front. Immunol. 2019, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagneux, P.; Varki, A. Evolutionary considerations in relating oligosaccharide diversity to biological function. Glycobiology 1999, 9, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Gagneux, P. Human-specific evolution of sialic acid targets: Explaining the malignant malaria mystery? Proc. Natl. Acad. Sci. USA 2009, 106, 14739–14740. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, D.; Springer, S.A.; Ma, F.; Cohen, M.; Secrest, P.; Taylor, R.E.; Varki, A.; Gagneux, P. Sexual selection by female immunity against paternal antigens can fix loss of function alleles. Proc. Natl. Acad. Sci. USA 2011, 108, 17743–17748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangvoranuntakul, P.; Gagneux, P.; Diaz, S.; Bardor, M.; Varki, N.; Varki, A.; Muchmore, E. Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc. Natl. Acad. Sci. USA 2003, 100, 12045–12050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardor, M.; Nguyen, D.H.; Diaz, S.; Varki, A. Mechanism of Uptake and Incorporation of the Non-human Sialic Acid N -Glycolylneuraminic Acid into Human Cells. J. Biol. Chem. 2005, 280, 4228–4237. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.E.; Gregg, C.J.; Padler-Karavani, V.; Ghaderi, D.; Yu, H.; Huang, S.; Sorensen, R.U.; Chen, X.; Inostroza, J.; Nizet, V.; et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid N-glycolylneuraminic acid. J. Exp. Med. 2010, 207, 1637–1646. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, N.; Matthews, S.; Soldati-Favre, D. Sialic acids: Key determinants for invasion by the Apicomplexa. Int. J. Parasitol. 2010, 40, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Neu, U.; Bauer, J.; Stehle, T. Viruses and sialic acids: Rules of engagement. Curr. Opin. Struct. Biol. 2011, 21, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.; Herrler, G.; Klenk, H.D. Sialic Acid Receptors of Viruses. Top. Curr. Chem. 2015, 367, 1–28. [Google Scholar] [CrossRef]
- Kato, K.; Ishiwa, A. The role of carbohydrates in infection strategies of enteric pathogens. Trop. Med. Health 2015, 43, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Haines-Menges, B.L.; Whitaker, W.B.; Lubin, J.B.; Boyd, E.F. Host Sialic Acids: A Delicacy for the Pathogen with Discerning Taste. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Juge, N.; Tailford, L.; Owen, C.D. Sialidases from gut bacteria: A mini-review. Biochem. Soc. Trans. 2016, 44, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Blaum, B.S.; Sthle, T. Sialic Acids in Nonenveloped Virus Infections. Adv. Carbohydr. Chem. Biochem. 2019, 76, 65–111. [Google Scholar] [CrossRef] [PubMed]
- van de Sandt, C.E.; Bodewes, R.; Rimmelzwaan, G.F.; de Vries, R.D. Influenza B viruses: Not to be discounted. Future Microbiol. 2015, 10, 1447–1465. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, M.; Fouchier, R.A.M. Role of receptor binding specificity in influenza A virus transmission and pathogenesis. EMBO J. 2014, 33, 823–841. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Veit, M. Hemagglutinin-esterase-fusion (HEF) protein of influenza C virus. Protein Cell 2016, 7, 28–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.V. Simian Virus 40 and Human Disease. J. Infect. Dis. 2004, 190, 2061–2064. [Google Scholar] [CrossRef] [Green Version]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; van Kuppeveld, F.J.M.; Boons, G.-J.; Bosch, B.-J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-Oacetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. USA 2018, 116, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Hulswit, R.J.G.; Widjaja, I.; Raj, V.S.; McBride, R.; Peng, W.; Widagdo, W.; Tortorici, M.A.; van Dieren, B.; Lang, Y.; et al. Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein. Proc. Natl. Acad. Sci. USA 2017, 114, E8508–E8517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amonsen, M.; Smith, D.F.; Cummings, R.D.; Air, G.M. Human parainfluenza viruses hPIV1 ad hPIV3 bind oligosaccharides with alpha2-3-linked sialic acids that are distinct from those bound by H5 avian influenza virus hemagglutinin. J. Virol. 2007, 81, 8341-5. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Portner, A.; Scroggs, R.A.; Uchikawa, M.; Koyama, N.; Matsu, K.; Suzuiki, Y.; Takimoto, T. Receptor Specificities of Human Respiroviruses. J. Virol. 2001, 75, 4604–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Bukreyev, A.; Thompson, C.I.; Watson, B.; Peeples, M.E.; Collins, P.L.; Pickles, R.J. Infection of ciliated cells by human parainfluenza virus type 3 in an in vitro model of human airway epithelium. J. Virol. 2005, 79, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Deng, L.; Stack, G.; Yu, H.; Chen, X.; Naito-Matsui, Y.; Varki, A.; Galán, J.E. Evolution of host adaptation in the Salmonella typhoid toxin. Nat. Microbiol. 2017, 2, 1592–1599. [Google Scholar] [CrossRef]
- Kappala, D.; Sarkhel, R.; Dixit, S.K.; Lalsangpuii; Mahawar, M.; Singh, M.; Ramakrishnan, S.; Goswami, T.K. Role of different receptors and actin filaments on Salmonella Typhimurium invasion in chicken macrophages. Immunobiology 2018, 223, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Muanprasat, C.; Chatsudthipong, V. Cholera: Pathophysiology and emerging therapeutic targets. Future Med. Chem. 2013, 5, 781–798. [Google Scholar] [CrossRef]
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to manage Pseudomonas aeruginosa infections. Drugs Context 2018, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Krivan, H.C.; Roberts, D.D.; Ginsburg, V. Many pulmonary pathogenic bacteria bind specifically to the carbohydrate sequence GalNAc beta 1-4Gal found in some glycolipids. Proc. Natl. Acad. Sci. USA 1988, 85, 6157–6161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, B.; Sachdev, G.P.; Cummings, R.D. Pseudomonas aeruginosa mucoid strain 8830 binds glycans containing the sialyl-Lewis x epitope. Glycoconj. J. 2006, 24, 87–95. [Google Scholar] [CrossRef]
- Evans, D.G.; Evans, D.J.; Moulds, J.J.; Graham, D.Y. N-acetylneuraminyllactose-binding fibrillar hemagglutinin of Campylobacter pylori: A putative colonization factor antigen. Infect. Immun. 1988, 56, 2896–2906. [Google Scholar] [CrossRef] [Green Version]
- Mahdavi, J. Helicobacter pylori SabA Adhesin in Persistent Infection and Chronic Inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, N.; Angstrom, J.; Hurtig, M.; Larsson, T.; Boren, T.; Teneberg, S. Helicobacter pylori and Complex Gangliosides. Infect. Immun. 2004, 72, 1519–1529. [Google Scholar] [CrossRef] [Green Version]
- Benktander, J.; Barone, A.; Johansson, M.M.; Teneberg, S. Helicobacter pylori SabA binding gangliosides of human stomach. Virulence 2018, 9, 738–751. [Google Scholar] [CrossRef] [Green Version]
- Bennett, H.J.; Roberts, I.S. Identification of a new sialic acid-binding protein in Helicobacter pylori. FEMS Immunol. Med. Microbiol. 2005, 44, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsohn, E.; Nystrom, J.; Bolin, I.; Nilsson, C.L.; Svennerholm, A.-M. HpaA is Essential for Helicobacter pylori Colonization in Mice. Infect. Immun. 2006, 74, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Ng, P.S.K.; Day, C.J.; Atack, J.M.; Hartley-Tassell, L.E.; Winter, L.E.; Marshanski, T.; Padler-Karavani, V.; Varki, A.; Barenkamp, S.J.; Apicella, M.A.; et al. Nontypeable Haemophilus influenzae Has Evolved Preferential Use of N- Acetylneuraminic Acid as a Host Adaptation. mBio 2019, 10, e00422-19. [Google Scholar] [CrossRef] [Green Version]
- Greiner, L.L.; Watanabe, H.; Phillips, N.J.; Shao, J.; Morgan, A.; Zaleski, A.; Gibson, B.W.; Apicella, M.A. Nontypeable Haemophilus influenzae Strain 2019 Produces a Biofilm Containing N-Acetylneuraminic Acid That May Mimic Sialylated O-Linked Glycans. Infect. Immun. 2004, 72, 4249–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, J.W.; Coussens, N.P.; Allen, S.; Houtman, J.C.D.; Turner, K.H.; Zaleski, A.; Ramaswamy, S.; Gibson, B.W.; Apicella, M.A. Characterization of the N -Acetyl-5-neuraminic Acid-binding Site of the Extracytoplasmic Solute Receptor (SiaP) of Nontypeable Haemophilus influenzae Strain 2019. J. Biol. Chem. 2008, 283, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Fujinaga, Y.; Inoue, K.; Nomura, T.; Sasaki, J.; Marvaud, J.C.; Popoff, M.R.; Kozaki, S.; Oguma, K. Identification and characterization of functional subunits of Clostridium botulinum type A progenitor toxin involved in binding to intestinal microvilli and erythrocytes. FEBS Lett. 2000, 467, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Rummel, A.; Eichner, T.; Weil, T.; Karnath, T.; Gutcaits, A.; Mahrhold, S.; Sandhoff, K.; Proia, R.L.; Acharya, K.R.; Bigalke, H.; et al. Identification of the protein receptor binding site of botulinum neurotoxins B and G proves the double-receptor concept. Proc. Natl. Acad. Sci. USA 2007, 104, 359–364. [Google Scholar] [CrossRef] [Green Version]
- Sagane, Y.; Hayashi, S.; Akiyama, T.; Matsumoto, T.; Hasegawa, K.; Yamano, A.; Suzuki, T.; Niwa, K.; Watanabe, T.; Yajima, S. Conformational divergence in the HA-33/HA-17 trimer of serotype C and D botulinum toxin complex. Biochem. Biophys. Res. Commun. 2016, 476, 280–285. [Google Scholar] [CrossRef]
- Sugawara, Y.; Iwamori, M.; Matsumura, T.; Yutani, M.; Amatsu, S.; Fujinaga, Y. Clostridium botulinum type C hemagglutinin affects the morphology and viability of cultured mammalian cells via binding to the ganglioside GM3. FEBS J. 2015, 282, 3334–3347. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, B.L.; Hale, J.Y.; Briles, D.E. Free Sialic Acid Acts as a Signal That Promotes Streptococcus pneumoniae Invasion of Nasal Tissue and Nonhematogenous Invasion of the Central Nervous System. Infect. Immun. 2016, 84, 2607–2615. [Google Scholar] [CrossRef] [Green Version]
- Coats, M.T.; Murphy, T.; Paton, J.C.; Gray, B.; Briles, D.E. Exposure of Thomsen-Friedenreich antigen in Streptococcus pneumoniae infection is dependent on pneumococcal neuraminidase A. Microb. Pathog. 2011, 50, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byres, E.; Paton, A.W.; Paton, J.C.; Löfling, J.C.; Smith, D.F.; Wilce, M.C.J.; Talbot, U.M.; Chong, D.C.; Yu, H.; Huang, S.; et al. Incorporation of a non-human glycan mediates human susceptibility to a bacterial toxin. Nature 2008, 456, 648–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyahian, E.A.; Oltra, G.; Ochoa, F.; Melendi, S.; Hermes, R.; Paton, J.C.; Paton, A.W.; Lago, N.; Castro Parodi, M.; Damiano, A.; et al. Systemic effects of Subtilase cytotoxin produced by Escherichia coli O113:H21. Toxicon 2017, 127, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Tolia, N.H.; Enemark, E.J.; Sim, B.K.L.; Joshua-Tor, L. Structural Basis for the EBA-175 Erythrocyte Invasion Pathway of the Malaria Parasite Plasmodium falciparum. Cell 2005, 122, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Rydzak, J.; Kaczmarek, R.; Czerwinski, M.; Lukasiewicz, J.; Tyborowska, J.; Szewczyk, B.; Jaskiewicz, E. The Baculovirus-Expressed Binding Region of Plasmodium falciparum EBA-140 Ligand and Its Glycophorin C Binding Specificity. PLoS ONE 2015, 10, e0115437. [Google Scholar] [CrossRef] [Green Version]
- Zerka, A.; Kaczmarek, R.; Czerwinski, M.; Jaskiewicz, E. Plasmodium reichenowi EBA-140 merozoite ligand binds to glycophorin D on chimpanzee red blood cells, shedding new light on origins of Plasmodium falciparum. Parasites Vectors 2017, 10, 554. [Google Scholar] [CrossRef] [Green Version]
- Freire-de-Lima, L.; Oliveira, I.A.; Neves, J.L.; Penha, L.L.; Alisson-Silva, F.; Dias, W.B.; Todeschini, A.R. Sialic acid: A sweet swing between mammalian host and Trypanosoma cruzi. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Takegawa, Y.; Ralston, K.S.; Gilchrist, C.A.; Hamano, S.; Petri, W.A.; Shinohara, Y. Sialic acid-dependent attachment of mucins from three mouse strains to Entamoeba histolytica. Biochem. Biophys. Res. Commun. 2013, 436, 252–258. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, Y.; Ogiso, A.; Kameyama, K.; Nishimura, M.; Xuan, X.; Ikehara, Y. α2-3 Sialic acid glycoconjugate loss and its effect on infection with Toxoplasma parasites. Exp. Parasitol. 2013, 135, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Jilani, T.N.; Jamil, R.T.; Siddiqui, A.H. H1N1 Influenza; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Gamblin, S.J.; Skehel, J.J. Influenza Hemagglutinin and Neuraminidase Membrane Glycoproteins. J. Biol. Chem. 2010, 285, 28403–28409. [Google Scholar] [CrossRef] [Green Version]
- Overeem, N.J.; Hamming, P.H.E.; Grant, O.C.; Di Iorio, D.; Tieke, M.; Bertolino, M.C.; Li, Z.; Vos, G.; de Vries, R.P.; Woods, R.J.; et al. Hierarchical Multivalent Effects Control Influenza Host Specificity. ACS Cent. Sci. 2020, 6, 2311–2318. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Ross, T.M. H3N2 influenza viruses in humans: Viral mechanisms, evolution, and evaluation. Hum. Vaccines Immunother. 2018, 14, 1840–1847. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Takano, M.; Kurebayashi, Y.; Masuda, M.; Kawagishi, S.; Takaguchi, M.; Yamanaka, T.; Minami, A.; Otsubo, T.; Ikeda, K.; et al. N-Glycolylneuraminic Acid on Human Epithelial Cells Prevents Entry of Influenza A Viruses That Possess N-Glycolylneuraminic Acid Binding Ability. J. Virol. 2014, 88, 8445–8456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellebø, A.; Vilas, U.; Falk, K.; Vlasak, R. Infectious Salmon Anemia Virus Specifically Binds to and Hydrolyzes 4-O-Acetylated Sialic Acids. J. Virol. 2004, 78, 3055–3362. [Google Scholar] [CrossRef] [Green Version]
- Vilchez, R.A.; Butel, J.S. Emergent Human Pathogen Simian Virus 40 and Its Role in Cancer. Clin. Microbiol. Rev. 2004, 17, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, M.; Gazdar, A.; Butel, J.S. SV40 and human mesothelioma. Transl. Lung Cancer Res. 2020, 9, 47–59. [Google Scholar] [CrossRef]
- Neu, U.; Woellner, K.; Gauglitz, G.; Stehle, T. Structural basis of GM1 ganglioside recognition by simian virus 40. Proc. Natl. Acad. Sci. USA 2008, 105, 5219–5224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanero-Rhodes, M.A.; Smith, A.; Chai, W.; Sonnino, S.; Mauri, L.; Childs, R.A.; Zhang, Y.; Ewers, H.; Helenius, A.; Imberty, A.; et al. N-Glycolyl GM1 Ganglioside as a Receptor for Simian Virus 40. J. Virol. 2007, 81, 12846–12858. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, J.C.; Mazzoni, E.; Bononi, I.; Tognon, M.; Martini, F. Association Between Simian Virus 40 and Human Tumors. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Harapan, H.; Itoh, N.; Yufika, A.; Winardi, W.; Keam, S.; Te, H.; Megawati, D.; Hayati, Z.; Wagner, A.L.; Mudatsir, M. Coronavirus disease 2019 (COVID-19): A literature review. J. Infect. Public Health 2020, 13, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Hu, B.; Huang, X.; Chai, Y.; Zhou, D.; Wang, Y.; Shuai, H.; Yang, D.; Hou, Y.; Zhang, X.; et al. Host and viral determinants for efficient SARS-CoV-2 infection of the human lung. Nat. Commun. 2021, 12, 134. [Google Scholar] [CrossRef]
- Sun, X.-L. The role of cell surface sialic acids for SARS-CoV-2 infection. Glycobiology 2021. [Google Scholar] [CrossRef]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [Green Version]
- Regl, G.; Kaser, A.; Iwersen, M.; Schmid, H.; Kohla, G.; Strobl, B.; Vilas, U.; Schauer, R.; Vlasak, R. The Hemagglutinin-Esterase of Mouse Hepatitis Virus Strain S is a Sialate-4-O-Acetylesterase. J. Virol. 1999, 73, 4721–4727. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, K.; Kasai, M.; Kato, S.; Kasai, H.; Hatakeyama, K. Haemagglutinin-esterase protein (HE) of murine corona virus: DVIMD (diarrhea virus of infant mice). Arch. Virol. 1998, 143, 1523–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawełczyk, M.; Kowalski, M.L. The Role of Human Parainfluenza Virus Infections in the Immunopathology of teh Respiratory Tract. Curr. Allergy Asthma Rep. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Farahmans, M.; Malekshahi, S.S.; Jebbari, M.R.; Shayestehpour, M. The landscape of extrapulmonay manifestations of human parainfluenza viruses: A systematic narrative review. Microbiol. Immunol. 2020, 65, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Branche, A.E.; Falsey, A.R. Parainfluenza Virus Infection. Semin. Respir. Crit. Care Med. 2016, 37, 538–554. [Google Scholar] [CrossRef]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spano, S.; Ugalde, J.E.; Galan, J.E. Delivery of a Salmonella Typhi Exotoxin from a Host Intracellular Compartment. Cell Host Microbe 2008, 3, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Acheson, D.; Hohmann, E.L. Nontyphoidal Salmonellosis. Clin. Infect. Dis. 2001, 32, 263–269. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, M.; Tanaka, K.; Nishimori, K.; Makino, S.; Kanno, T.; Ishihara, R.; Hatama, S.; Kitano, R.; Kishima, M.; Sameshima, T.; et al. The artAB genes encode a putative ADP-ribosyltransferase toxin homologue associated with Salmonella enterica serovar Typhimurium DT104. Microbiology 2005, 151, 3089–3096. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Ishibashi, S.; Ikeda, K.; Ishitsubo, E.; Hori, T.; Tokiwa, H.; Taguchi, R.; Ieno, D.; Otsubo, T.; Matsuda, Y.; et al. Catalytic preference of Salmonella typhimurium LT2 sialidase for N -acetylneuraminic acid residues over N-glycolylneuraminic acid residues. FEBS Open Bio 2013, 3, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alisson-Silva, F.; Liu, J.Z.; Diaz, S.L.; Deng, L.; Gareau, M.G.; Marchelletta, R.; Chen, X.; Nizet, V.; Varki, N.; Barrett, K.E.; et al. Human evolutionary loss of epithelial Neu5Gc expression and species-specific susceptibility to cholera. PLoS Pathog. 2018, 14, e1007133. [Google Scholar] [CrossRef] [Green Version]
- Van den Broeck, D.; Horvath, C.; De Wolf, M.J.S. Vibrio cholerae: Cholera toxin. Int. J. Biochem. Cell Biol. 2007, 39, 1771–1775. [Google Scholar] [CrossRef]
- Alonso, A.; Rojo, F.; Martinez, J.L. Environmental and clinical isolates of Pseudomonas aeruginosa show pathogenic and biodegradative properties irrespective of their origin. Environ. Microbiol. 1999, 1, 421–430. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Chua, S.L.; Yam, J.K.H.; Hao, P.; Adav, S.S.; Salido, M.M.; Liu, Y.; Givskov, M.; Sze, S.K.; Tolker-Nielsen, T.; Yang, L. Selective labelling and eradication of antibiotic-tolerant bacterial populations in Pseudomonas aeruginosa biofilms. Nat. Commun. 2016, 7, 10750. [Google Scholar] [CrossRef] [Green Version]
- Bucior, I.; Pielage, J.F.; Engel, J.N. Pseudomonas aeruginosa Pili and Flagella Mediate Distinct Binding and Signaling Events at the Apical and Basolateral Surface of Airway Epithelium. PLoS Pathog. 2012, 8, e1002616. [Google Scholar] [CrossRef] [Green Version]
- Lau, G.W.; Hassett, D.J.; Ran, H.; Kong, F. The role of pyocyanin in Pseudomonas aeruginosa infection. Trends Microbiol. 2004, 10, 599–606. [Google Scholar] [CrossRef]
- Blier, A.-S.; Veron, W.; Bazire, A.; Gerault, E.; Taupin, L.; Vieillard, J.; Rehel, K.; Dufour, A.; Le Derf, F.; Orange, N.; et al. C-type natriuretic peptide modulates quorum sensing molecule and toxin production in Pseudomonas aeruginosa. Microbiology 2011, 157, 1929–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Sokol, S.; Soong, G.; Gomez, M.I.; Prince, A. Pseudomonas aeruginosa Flagella Activate Airway Epithelial Cells through asialoGM1 and Toll-Like Receptor 2 as well as Toll-Like Receptor 5. Am. J. Respir. Cell Mol. Biol. 2004, 30, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. The Pseudomonas aeruginosa Flagellar Cap Protein, FliD, is Responsible for Mucin Adhesion. Infect. Immun. 1998, 66, 1000–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotkowski, M.-C.; Costa, A.O.; Morandi, V.; Barbosa, H.S.; Nader, H.B.; Bentzmann, S.D.E.; Puchelle, E. Role of heparan sulphate proteoglycans as potential receptors for non-piliated Pseudomonas aeruginosa adherence to non-polarised airway epithelial cells. Infect. Immun. 2001, 50, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Bucior, I.; Mostov, K.; Engel, J.N. Pseudomonas Aeruginosa-Mediated Damage Requires Distinct Receptors at the Apical and Basolateral Surfaces of the Polarized Epithelium. Infect. Immun. 2010, 78, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Plotkowski, M.-C.; Tournier, J.-M.; Puchelle, E. Pseudomonas aeruginosa Strains Possess Specific Adhesins for Laminin. Infect. Immun. 1996, 64, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Paulsson, M.; Su, Y.-C.; Ringwood, T.; Uddén, F.; Riesbeck, K. Pseudomonas aeruginosa uses multiple receptors for adherence to laminin during infection of the respiratory tract and skin wounds. Sci. Rep. 2019, 9, 18168. [Google Scholar] [CrossRef]
- Leroy-Dudal, J.; Gagnière, H.; Cossard, E.; Carreiras, F.; Di Martino, P. Role of αvβ5 integrins and vitronectin in Pseudomonas aeruginosa PAK interaction with A549 respiratory cells. Microbes Infect. 2004, 6, 875–881. [Google Scholar] [CrossRef]
- Liu, W.; Sun, T.; Wang, Y. Integrin αvβ6 mediates epithelial-mesenchymal transition in human bronchial epithelial cells induced by lipopolysaccharides of Pseudomonas aeruginosa via TGF-β1-Smad2/3 signaling pathway. Folia Microbiol. 2020, 65, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Thuenauer, R.; Landi, A.; Trefzer, A.; Altmann, S.; Wehrum, S.; Eierhoff, T.; Diedrich, B.; Dengjel, J.; Nyström, A.; Imberty, A.; et al. The Pseudomonas aeruginosa Lectin LecB Causes Integrin Internalization and Inhibits Epithelial Wound Healing. mBio 2020, 11, e03260-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiman, L.; Prince, A. Pseudomonas aeruginosa pili bind to asialoGM1 which is increased on the surface of cystic fibrosis epithelial cells. J. Clin. Investig. 1993, 92, 1875–1880. [Google Scholar] [CrossRef] [Green Version]
- Kidd, M.; Modlin, I.M. A Century of Helicobacter pylori. Digestion 1998, 59, 1–15. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Bashir, A.H.H.; Aljarbou, A.N.; Ramadan, A.M.; Muddathir, A.K.; AlHoufie, S.T.S.; Hifny, A.; Elhassan, G.O.; Ibrahim, M.E.; Alqahtani, S.S.; et al. The Possible Role of Helicobacter pylori in Gastric Cancer and Its Management. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Gu, H. Role of Flagella in the Pathogenesis of Helicobacter pylori. Curr. Microbiol. 2017, 74, 863–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambelli, B.; Musiani, F.; Benini, S.; Ciurli, S. Chemistry of Ni 2+ in Urease: Sensing, Trafficking, and Catalysis. Acc. Chem. Res. 2011, 44, 520–530. [Google Scholar] [CrossRef] [PubMed]
- King, P. Haemophilus influenzae and the lung (Haemophilus and the lung). Clin. Transl. Med. 2012, 1. [Google Scholar] [CrossRef] [Green Version]
- Zarei, A.E.; Linjawi, M.H.; Redwan, E.M. Circulating innate and adaptive immunity against anti-Haemophilus influenzae type b. Hum. Antibodies 2019, 27, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Forstner, C.; Rohde, G.; Rupp, J.; Schuette, H.; Ott, S.R.; Hagel, S.; Harrison, N.; Thalhammer, F.; von Baum, H.; Suttorp, N.; et al. Community-acquired Haemophilus influenzae pneumonia—New insights from the CAPNETZ study. J. Infect. 2016, 72, 554–563. [Google Scholar] [CrossRef]
- Livorsi, D.J.; MacNeil, J.R.; Cohn, A.C.; Bareta, J.; Zansky, S.; Petit, S.; Gershman, K.; Harrison, L.H.; Lynfield, R.; Reingold, A.; et al. Invasive Haemophilus influenzae in the United States, 1999–2008: Epidemiology and outcomes. J. Infect. 2012, 65, 496–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderson, A.R.; Leid, J.G.; Hunsaker, D. Bacterial Biofilms on the Sinus Mucosa of Human Subjects with Chronic Rhinosinusitis. Laryngoscope 2006, 116, 1121–1126. [Google Scholar] [CrossRef]
- Epling, J. Bacterial conjuctivitis. BMJ Clin. Evid. 2012, 2, 1–21. [Google Scholar]
- Hall-Stoodley, L.; Hu, F.Z.; Gieseke, A.; Nistico, L.; Nguyen, D.; Hayes, J.; Forbes, M.; Greenberg, D.P.; Dice, B.; Burrows, A.; et al. Direct Detection of Bacterial Biofilms on the Middle-Ear Mucosa of Children with Chronic Otitis Media. JAMA 2006, 296, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, N.; Bakaletz, L.O. Selective adherence of non-typeable Haemophilus influenzae(NTHi) to mucus or epithelial cells in the chinchilla Eustachian tube and middle ear. Microb. Pathog. 1996, 21, 343–356. [Google Scholar] [CrossRef]
- Reddy, M.; Murphy, T.; Faden, H.; Bernstein, J. Middle ear mucin glycoprotein: Purification and interaction with nontypable and. Otolaryngol. Head Neck Surg. 1997, 116, 175–180. [Google Scholar] [CrossRef]
- Hallström, T.; Singh, B.; Resman, F.M.; Blom, A.; Mörgelin, M.; Riesbeck, K. Haemophilus influenzae Protein E Binds to the Extracellular Matrix by Concurrently Interacting with Laminin and Vitronectin. J. Infect. Dis. 2011, 204, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-C.; Halang, P.; Fleury, C.; Jalalvand, F.; Mörgelin, M.; Riesbeck, K. Haemophilus Protein F Orthologs of Pathogens Infecting the Airways: Exploiting Host Laminin at Heparin-Binding Sites for Maximal Adherence to Epithelial Cells. J. Infect. Dis. 2017, 216, 1303–1307. [Google Scholar] [CrossRef]
- Jalalvand, F.; Su, Y.-C.; Mörgelin, M.; Brant, M.; Hallgren, O.; Westergren-Thorsson, G.; Singh, B.; Riesbeck, K. Haemophilus influenzae Protein F Mediates Binding to Laminin and Human Pulmonary Epithelial Cells. J. Infect. Dis. 2013, 207, 803–813. [Google Scholar] [CrossRef] [Green Version]
- St Geme, J.W.; Falkow, S.; Barenkamp, S.J. High-molecular-weight proteins of nontypable Haemophilus influenzae mediate attachment to human epithelial cells. Proc. Natl. Acad. Sci. USA 1993, 90, 2875–2879. [Google Scholar] [CrossRef] [Green Version]
- Barenkamp, S.J.; St Geme, J.W. Identification of a second family of high-molecular-weight adhesion proteins expressed by non-typable Haemophilus influenzae. Mol. Microbiol. 1996, 19, 1215–1223. [Google Scholar] [CrossRef]
- Gilsdorf, J.R.; McCrea, K.W.; Marrs, C.F. Role of pili in Haemophilus influenzae adherence and colonization. Infect. Immun. 1997, 65, 2997–3002. [Google Scholar] [CrossRef] [Green Version]
- Mu, X.-Q.; Egelman, E.H.; Bullitt, E. Structure and Function of Hib Pili from Haemophilus influenzae Type b. J. Bacteriol. 2002, 184, 4868–4874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clementi, C.F.; Håkansson, A.P.; Murphy, T.F. Internalization and Trafficking of Nontypeable Haemophilus influenzae in Human Respiratory Epithelial Cells and Roles of IgA1 Proteases for Optimal Invasion and Persistence. Infect. Immun. 2014, 82, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Morey, P.; Viadas, C.; Euba, B.; Hood, D.W.; Barberan, M.; Gil, C.; Grillo, M.J.; Bengoechea, J.A.; Garmendia, J. Relative Contributions of Lipooligosaccharide Inner and Outer Core Modifications to Nontypeable Haemophilus influenzae Pathogenesis. Infect. Immun. 2013, 81, 4100–4111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrillo-Marquez, M.A. Botulism. Pediatrics Rev. 2016, 37, 183–192. [Google Scholar] [CrossRef]
- Connan, C.; Popoff, M.R. Two-component systems and toxinogenesis regulation in Clostridium botulinum. Res. Microbiol. 2015, 166, 332–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosow, L.K.; Strober, J.B. Infant Botulism: Review and Clinical Update. Pediatric Neurol. 2015, 52, 487–492. [Google Scholar] [CrossRef]
- Yamashita, S.; Yoshida, H.; Uchiyama, N.; Nakakita, Y.; Nakakita, S.; Tonozuka, T.; Oguma, K.; Nishikawa, A.; Kamitori, S. Carbohydrate recognition mechanism of HA70 from Clostridium botulinum deduced from X-ray structures in complexes with sialylated oligosaccharides. FEBS Lett. 2012, 586, 2404–2410. [Google Scholar] [CrossRef] [Green Version]
- Henriques-Normark, B.; Tuomanen, E.I. The Pneumococcus: Epidemiology, Microbiology, and Pathogenesis. Cold Spring Harb. Perspect. Med. 2013, 3, a010215. [Google Scholar] [CrossRef]
- Engholm, D.H.; Kilian, M.; Goodsell, D.S.; Andersen, E.S.; Kjærgaard, R.S. A visual review of the human pathogen Streptococcus pneumoniae. FEMS Microbiol. Rev. 2017, 41, 854–879. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Mitchell, T.J. Streptococcus pneumoniae: Virulence factors and variation. Clin. Microbiol. Infect. 2010, 16, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B. Pathogenic Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 355–356. [Google Scholar] [CrossRef]
- Figler, H.M.; Dudley, E.G. The interplay of Escherichia coli O157:H7 and commensal E. coli: The importance of strain-level identification. Expert Rev. Gastroenterol. Hepatol. 2016, 10, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Capitani, G.; Eidam, O.; Glockshuber, R.; Grütter, M.G. Structural and functional insights into the assembly of type 1 pili from Escherichia coli. Microbes Infect. 2006, 8, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.W.; Kaijser, B. The local immune response to Escherichia coli O and K antigen in experimental pyelonephritis. J. Clin. Investig. 1976, 58, 276–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, J.L.; Martínez Ortiz, G.C.; Subedi, P.; Keerthikumar, S.; Mathivanan, S.; Paxman, J.J.; Heras, B. Autotransporter Adhesins in Escherichia coli Pathogenesis. Proteomics 2017, 17, 1600431. [Google Scholar] [CrossRef]
- Yu, F.; Mizushima, S. Roles of lipopolysaccharide and outer membrane protein OmpC of Escherichia coli K-12 in the receptor function for bacteriophage T4. J. Bacteriol. 1982, 151, 718–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nhu, N.T.K.; Phan, M.-D.; Forde, B.M.; Murthy, A.M.V.; Peters, K.M.; Day, C.J.; Poole, J.; Kidd, T.J.; Welch, R.A.; Jennings, M.P.; et al. Complex Multilevel Control of Hemolysin Production by Uropathogenic Escherichia coli. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Giorgi, M.E.; de Lederkremer, R.M. Trans-sialidase and mucins of Trypanosoma cruzi: An important interplay for the parasite. Carbohydr. Res. 2011, 346, 1389–1393. [Google Scholar] [CrossRef]
- Ruiz Díaz, P.; Mucci, J.; Meira, M.A.; Bogliotti, Y.; Musikant, D.; Leguizamón, M.S.; Campetella, O. Trypanosoma cruzi trans -Sialidase Prevents Elicitation of Th1 Cell Response via Interleukin 10 and Downregulates Th1 Effector Cells. Infect. Immun. 2015, 83, 2099–2108. [Google Scholar] [CrossRef] [Green Version]
- Agustí, R.; Giorgi, M.E.; de Lederkremer, R.M. The trans-sialidase from Trypanosoma cruzi efficiently transfers α-(2→3)-linked N-glycolylneuraminic acid to terminal β-galactosyl units. Carbohydr. Res. 2007, 342, 2465–2469. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Simpson, A.G.B.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The Revised Classification of Eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betanzos, A.; Bañuelos, C.; Orozco, E. Host Invasion by Pathogenic Amoebae: Epithelial Disruption by Parasite Proteins. Genes 2019, 10, 618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marie, C.; Petri, W.A. Regulation of Virulence of Entamoeba histolytica. Annu. Rev. Microbiol. 2014, 68, 493–520. [Google Scholar] [CrossRef]
- Rawat, A.; Singh, P.; Jyoti, A.; Kaushik, S.; Srivastava, V.K. Averting transmission: A pivotal target to manage amoebiasis. Chem. Biol. Drug Des. 2020, 96, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Udezulu, I.A.; Leitch, G.J. A membrane-associated neuraminidase in Entamoeba histolytica trophozoites. Infect. Immun. 1987, 55, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Nok, A.J.; Rivera, W. Characterization of sialidase from Entamoaeba hystolitica and possible pathogenic role in amebiasis. Parasitol. Res. 2003, 89, 302–307. [Google Scholar] [CrossRef]
- Kochanowsky, J.A.; Koshy, A.A. Toxoplasma gondii. Curr. Biol. 2018, 28, R770–R771. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.S.; Lodoen, M.B. Mechanisms of Human Innate Immune Evasion by Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Baba, M.; Sato, M.; Kitoh, K.; Takashima, Y. The distribution pattern of α2,3- and α2,6-linked sialic acids affects host cell preference in Toxoplasma gondii. Exp. Parasitol. 2015, 155, 74–81. [Google Scholar] [CrossRef]
- Friedrich, N.; Santos, J.M.; Liu, Y.; Palma, A.S.; Leon, E.; Saouros, S.; Kiso, M.; Blackman, M.J.; Matthews, S.; Feizi, T.; et al. Members of a Novel Protein Family Containing Microneme Adhesive Repeat Domains Act as Sialic Acid-binding Lectins during Host Cell Invasion by Apicomplexan Parasites. J. Biol. Chem. 2010, 285, 2064–2076. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Xing, M.; Ding, Y.; Wang, D.; Guo, X.; Sang, X.; Li, J.; Li, C.; Wang, Y.; Feng, Y.; et al. The Putative TCP-1 Chaperonin Is an Important Player Involved in Sialic Acid-Dependent Host Cell Invasion by Toxoplasma gondii. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Xing, M.; Yang, N.; Jiang, N.; Wang, D.; Sang, X.; Feng, Y.; Chen, R.; Wang, X.; Chen, Q. A Sialic Acid-Binding Protein SABP1 of Toxoplasma gondii Mediates Host Cell Attachment and Invasion. J. Infect. Dis. 2020, 222, 126–135. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Learn, G.H.; Rudicell, R.S.; Robertson, J.D.; Keele, B.F.; Ndjango, J.-B.N.; Sanz, C.M.; Morgan, D.B.; Locatelli, S.; et al. Origin of the human malaria parasite Plasmodium falciparum in gorillas. Nature 2010, 467, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Prugnolle, F.; Durand, P.; Neel, C.; Ollomo, B.; Ayala, F.J.; Arnathau, C.; Etienne, L.; Mpoudi-Ngole, E.; Nkoghe, D.; Leroy, E.; et al. African great apes are natural hosts of multiple related malaria species, including Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2010, 107, 1458–1463. [Google Scholar] [CrossRef] [Green Version]
- Gaur, D.; Chitnis, C.E. Molecular interactions and signaling mechanisms during erythrocyte invasion by malaria parasites. Curr. Opin. Microbiol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Tham, W.-H.; Healer, J.; Cowman, A.F. Erythrocyte and reticulocyte binding-like proteins of Plasmodium falciparum. Trends Parasitol. 2012, 28, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.H.; Blair, P.L.; Kaneko, O.; Peterson, D.S. An expanding ebl family of Plasmodium falciparum. Trends Parasitol. 2001, 17, 297–299. [Google Scholar] [CrossRef]
- Wanaguru, M.; Crosnier, C.; Johnson, S.; Rayner, J.C.; Wright, G.J. Biochemical Analysis of the Plasmodium falciparum Erythrocyte-binding Antigen-175 (EBA175)-Glycophorin-A Interaction. J. Biol. Chem. 2013, 288, 32106–32117. [Google Scholar] [CrossRef] [Green Version]
- Salinas, N.D.; Paing, M.M.; Tolia, N.H. Critical Glycosylated Residues in Exon Three of Erythrocyte Glycophorin a Engage Plasmodium falciparum EBA-175 and Define Receptor Specificity. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.K.; Triglia, T.; Reed, M.B.; Cowman, A.F. A novel ligand from Plasmodium falciparum that binds to a sialic acid-containing receptor on the surface of human erythrocytes. Mol. Microbiol. 2001, 41, 47–58. [Google Scholar] [CrossRef]
- Narum, D.L.; Fuhrmann, S.R.; Luu, T.; Sim, B.K.L. A novel Plasmodium falciparum erythrocyte binding protein-2 (EBP2/BAEBL) involved in erythrocyte receptor binding. Mol. Biochem. Parasitol. 2002, 119, 159–168. [Google Scholar] [CrossRef]
- Gilberger, T.-W.; Thompson, J.K.; Triglia, T.; Good, R.T.; Duraisingh, M.T.; Cowman, A.F. A Novel Erythrocyte Binding Antigen-175 Paralogue from Plasmodium falciparum Defines a New Trypsin-resistant Receptor on Human Erythrocytes. J. Biol. Chem. 2003, 278, 14480–14486. [Google Scholar] [CrossRef] [Green Version]
- Lobo, C.-A.; Rodriguez, M.; Reid, M.; Lustigman, S. Glycophorin C is the receptor for the Plasmodium falciparum erythrocyte binding ligand PfEBP-2 (baebl). Blood 2003, 101, 4628–4631. [Google Scholar] [CrossRef]
- Maier, A.G.; Duraisingh, M.T.; Reeder, J.C.; Patel, S.S.; Kazura, J.W.; Zimmerman, P.A.; Cowman, A.F. Plasmodium falciparum erythrocyte invasion through glycophorin C and selection for Gerbich negativity in human populations. Nat. Med. 2003, 9, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Duriseti, S.; Sun, P.; Miller, L.H. Molecular basis of binding of the Plasmodium falciparum receptor BAEBL to erythrocyte receptor glycophorin C. Mol. Biochem. Parasitol. 2009, 168, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Jaskiewicz, E.; Peyrard, T.; Kaczmarek, R.; Zerka, A.; Jodlowska, M.; Czerwinski, M. The Gerbich blood group system: Old knowledge, new importance. Transfus. Med. Rev. 2018, 32, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Mayer, D.C.G.; Jiang, L.; Achur, R.N.; Kakizaki, I.; Gowda, D.C.; Miller, L.H. The glycophorin C N-linked glycan is a critical component of the ligand for the Plasmodium falciparum erythrocyte receptor BAEBL. Proc. Natl. Acad. Sci. USA 2006, 103, 2358–2362. [Google Scholar] [CrossRef] [Green Version]
- Ashline, D.J.; Duk, M.; Lukasiewicz, J.; Reinhold, V.N.; Lisowska, E.; Jaskiewicz, E. The structures of glycophorin C N-glycans, a putative component of the GPC receptor site for Plasmodium falciparum EBA-140 ligand. Glycobiology 2015, 25, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.H.; Malpede, B.M.; Batchelor, J.D.; Tolia, N.H. Crystal and Solution Structures of Plasmodium falciparum Erythrocyte-binding Antigen 140 Reveal Determinants of Receptor Specificity during Erythrocyte Invasion. J. Biol. Chem. 2012, 287, 36830–36836. [Google Scholar] [CrossRef] [Green Version]
- Malpede, B.M.; Lin, D.H.; Tolia, N.H. Molecular Basis for Sialic Acid-dependent Receptor Recognition by the Plasmodium falciparum Invasion Protein Erythrocyte-binding Antigen-140/BAEBL. J. Biol. Chem. 2013, 288, 12406–12415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaskiewicz, E.; Jodłowska, M.; Kaczmarek, R.; Zerka, A. Erythrocyte glycophorins as receptors for Plasmodium merozoites. Parasites Vectors 2019, 12, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNicholas, S.; Potterton, E.; Wilson, K.S.; Noble, M.E. Presenting Your Structures: The CCP4mg Molecular-Graphics Software. Acta Crystallogr. Sect. D 2011, 67 Pt. 4, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Rayner, J.C.; Huber, C.S.; Barnwell, J.W. Conservation and divergence in erythrocyte invasion ligands: Plasmodium reichenowi EBL genes. Mol. Biochem. Parasitol. 2004, 138, 243–247. [Google Scholar] [CrossRef]
- Martin, M.J.; Rayner, J.C.; Gagneux, P.; Barnwell, J.W.; Varki, A. Evolution of human-chimpanzee differences in malaria susceptibility: Relationship to human genetic loss of N-glycolylneuraminic acid. Proc. Natl. Acad. Sci. USA 2005, 102, 12819–12824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanaguru, M.; Liu, W.; Hahn, B.H.; Rayner, J.C.; Wright, G.J. RH5-Basigin interaction plays a major role in the host tropism of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2013, 110, 20735–20740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dankwa, S.; Lim, C.; Bei, A.K.; Jiang, R.H.Y.; Abshire, J.R.; Patel, S.D.; Goldberg, J.M.; Moreno, Y.; Kono, M.; Niles, J.C.; et al. Ancient human sialic acid variant restricts an emerging zoonotic malaria parasite. Nat. Commun. 2016, 7, 11187. [Google Scholar] [CrossRef] [Green Version]
- Proto, W.R.; Siegel, S.V.; Dankwa, S.; Liu, W.; Kemp, A.; Marsden, S.; Zenonos, Z.A.; Unwin, S.; Sharp, P.M.; Wright, G.J.; et al. Adaptation of Plasmodium falciparum to humans involved the loss of an ape-specific erythrocyte invasion ligand. Nat. Commun. 2019, 10, 4512. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structure of sialic acids: (a) Neuraminic acid, (b) 2-keto-deoxynonulosonic acid, (c) N-acetylneuraminic acid, (d) N-glycolylneuraminic acid and (e) N-acetyl-9-O-acetylneuraminic acid.
Figure 1.
Structure of sialic acids: (a) Neuraminic acid, (b) 2-keto-deoxynonulosonic acid, (c) N-acetylneuraminic acid, (d) N-glycolylneuraminic acid and (e) N-acetyl-9-O-acetylneuraminic acid.

Figure 2.
Single protein maximum likelihood tree of CMAH protein sequences from various species. Sponge CMAH proteins form a clade and are grouped together with other metazoan sequences. Sequences were aligned using MUSCLE [29], trimmed in Geneious 8.1.9 (BioMatters, Ltd., Auckland, NZ) [30] and the tree was made in IQTree v2.1.2 [31] with the substitution model (LG+I+G4) selected using ModelFinder [32]. Nonparametric bootstrap branch supports are shown next to the nodes. As evident from the non-basal position of cnidarians, single protein trees only approximate true phylogenetic relationships.
Figure 2.
Single protein maximum likelihood tree of CMAH protein sequences from various species. Sponge CMAH proteins form a clade and are grouped together with other metazoan sequences. Sequences were aligned using MUSCLE [29], trimmed in Geneious 8.1.9 (BioMatters, Ltd., Auckland, NZ) [30] and the tree was made in IQTree v2.1.2 [31] with the substitution model (LG+I+G4) selected using ModelFinder [32]. Nonparametric bootstrap branch supports are shown next to the nodes. As evident from the non-basal position of cnidarians, single protein trees only approximate true phylogenetic relationships.

Figure 3.
Organization of the CMAH gene. The human gene lacks coding exon 3 in comparison to the mouse and the chimpanzee genes. The ticks show exons that are putatively still functional, and the crosses denote exons that were damaged in the human pseudogene, resulting in either premature STOP codons or inactivating mutations in the protein product. The exons are drawn to scale relative to each other, and the introns were rescaled to the same length.
Figure 3.
Organization of the CMAH gene. The human gene lacks coding exon 3 in comparison to the mouse and the chimpanzee genes. The ticks show exons that are putatively still functional, and the crosses denote exons that were damaged in the human pseudogene, resulting in either premature STOP codons or inactivating mutations in the protein product. The exons are drawn to scale relative to each other, and the introns were rescaled to the same length.

Figure 4.
The F2-binding pocket of the PfEBA-140 ligand with the bound N-acetylneuraminic acid. The amino acid residues and backbone atoms involved in sugar binding are shown (according to [205], modified). Figure made in ccp4mg [208] using PDB ID 4JNO.


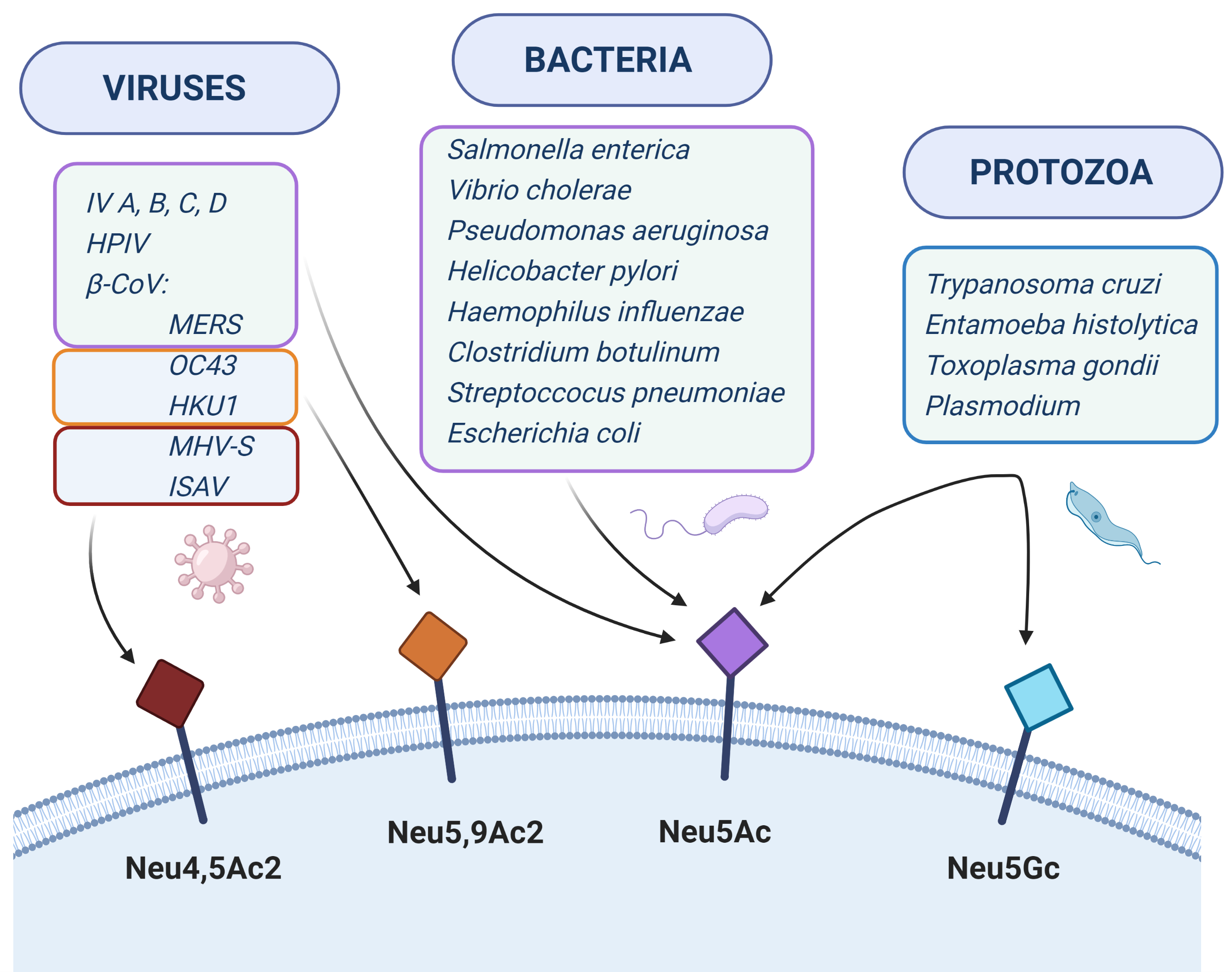{kind=link}
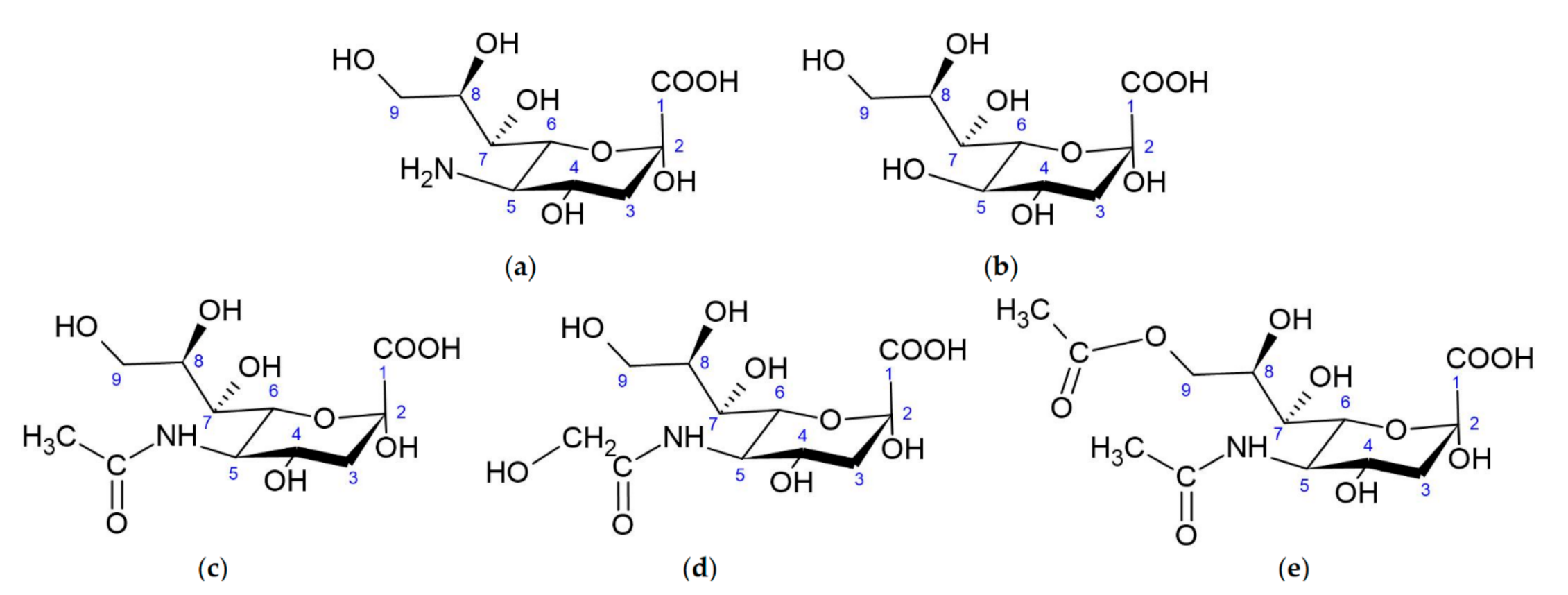{kind=link}
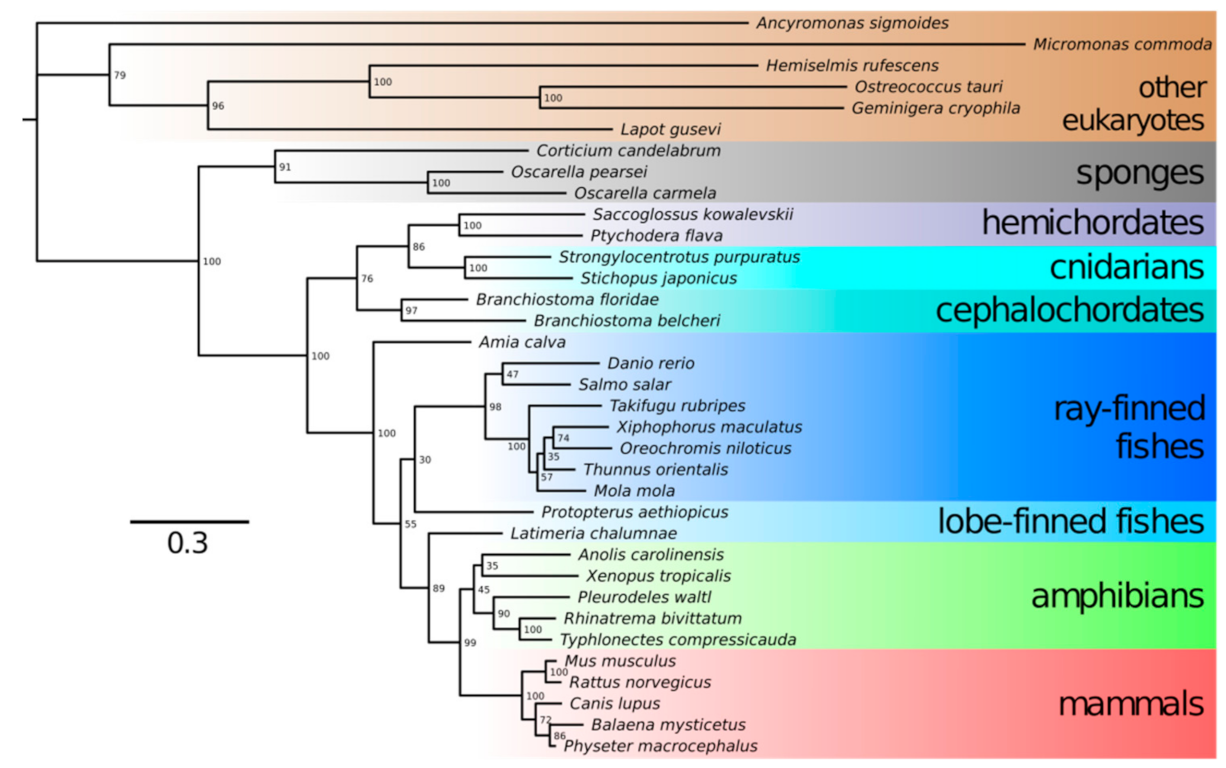{kind=link}
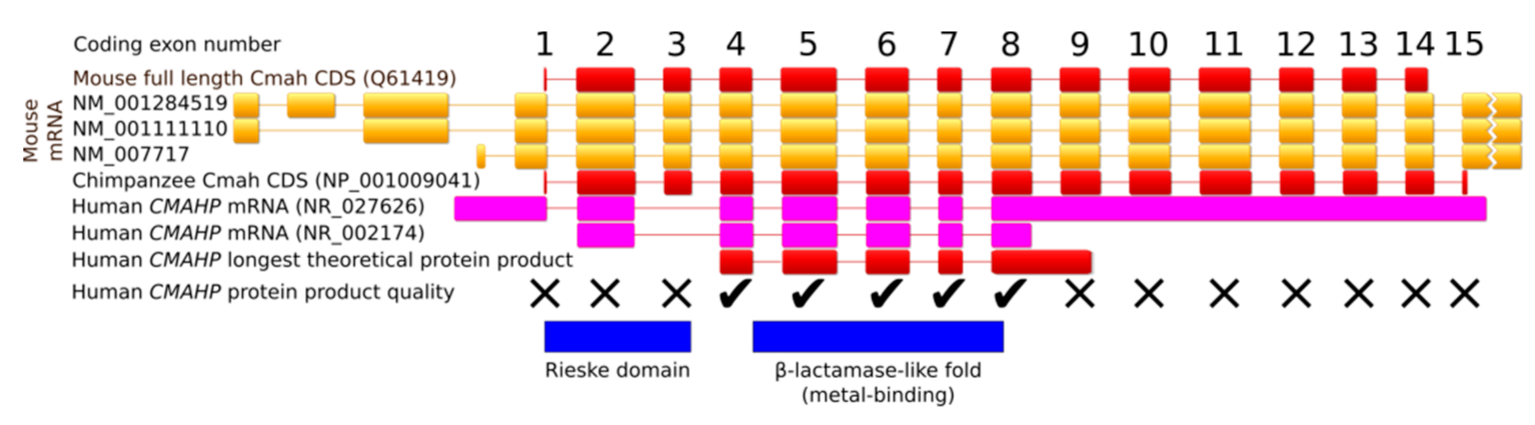{kind=link}
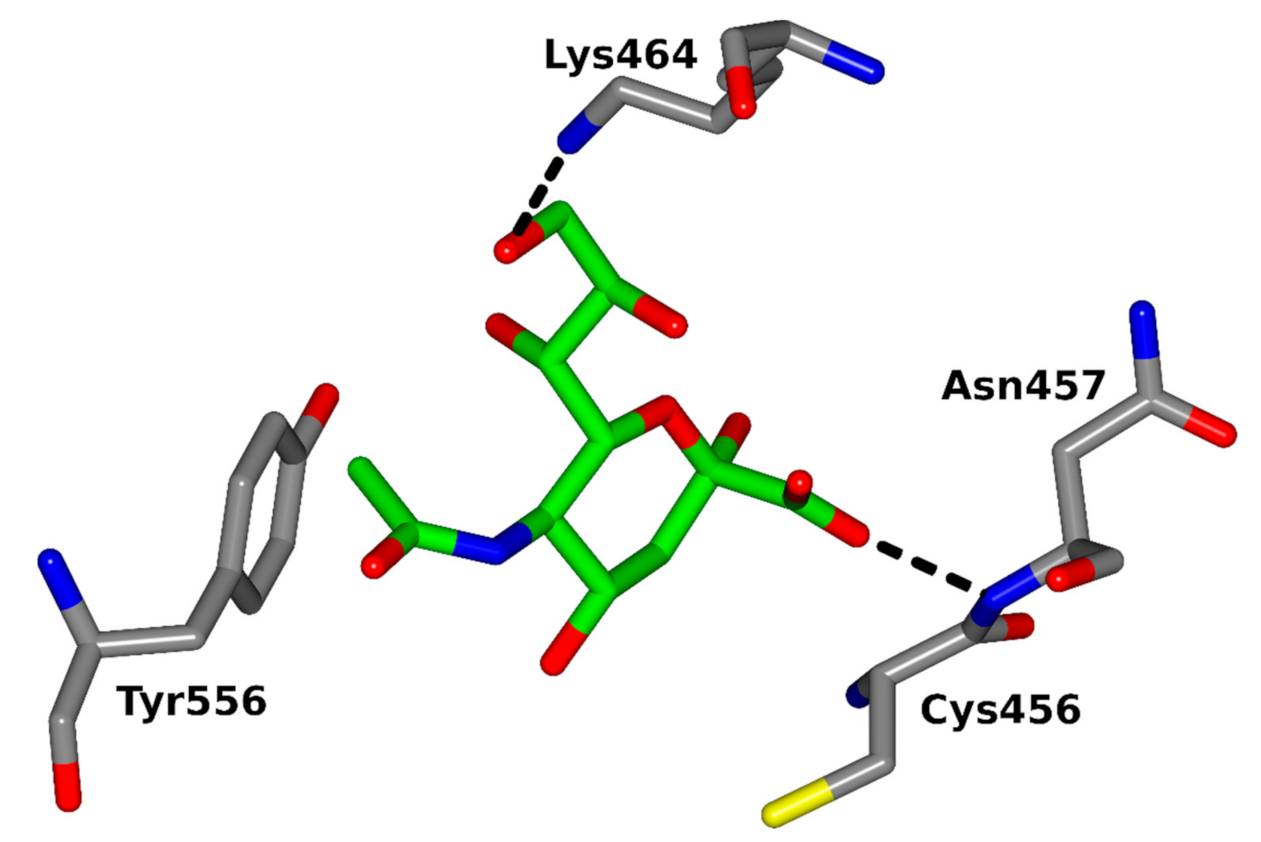{kind=link}
Table 1.
Pathogens exploiting sialic acids as receptors: viruses, bacteria and protozoa.
| Pathogen/Toxin | Disease | Ligand on Pathogen | Type of Sialic AcidBound/Incorporated | Symptoms in Humans | References |
|---|---|---|---|---|---|
| Avian influenza virus A | Flu | HA | α-2,3-Neu5Ac | Mild symptoms, fever, cough, sore throat, muscle anches | Van de Sandt et al., 2015 [50] |
| Human influenza virus A and B | Flu | HA | α-2,6-Neu5Ac/Neu5Gc 1 | Fever, cough, runny or stuffy nose, tiredness, sore throat, muscle anches | de Graaf and Fouchier 2014 [51] |
| Human influenza virus C | Flu | HEF | O-acetyl form of α-2,3- or α-2,6-Neu5Ac | Mild cough, fever, malaise | Wang and Veit 2016 [52] |
| Human influenza virus D | Flu in animals | HEF | O-acetyl form of α-2,3- or α-2,6-Neu5Ac | No symptoms detected in humans | Su et al., 2017 [53] |
| SV40 virus | Carcinogenic in humans | VP1 | NeuAc (weak binding in humans) and NeuGc (strong binding in simians) on GM1 glycolipid | Mesothelioma, osteosarcoma, pediatric and adult brain tumors and non-Hodgkin lymphomas | Shah 2004 [54] |
| Human betacoronaviruses - OC43 and HKU1 - MERS | Colds, human pulmonary infectious, lethal encephalitis (only OC43) Middle East respiratory syndrome | Spike protein | 9-O-acetylated Neu5Ac α-2,3-Neu5Ac | Fever, weakness, abdominal pain, rhinitis, and sore throat, sometimes severe pneumonia (HKU1), vomiting, diarrhea, abdominal pain (OC43) Fever, cough, breath shortness, pneumonia, sometimes diarrhea | Hulswit et al., 2018 [55] Li et al., 2017 [56] |
| Human parainfluenza viruses | Respiratory tract infections | HN | α-2,3-Neu5Ac α-2,6-Sias (HPIV-3) | cold-like, bronchiolitis, tracheobronchitis, laryngotracheobronchitis, pneumonia | Amonsen et al., 2007 [57]; Suzuki et al., 2001 [58]; Zhang et al., 2005 [59] |
| Salmonella enterica - S. Typhi - S. Typhimurium (LT2) | Typhoid fever Bacteremia and acute gastroenteritis | B subunit of toxin | α-2,3-Neu5Ac α-2,3-Neu5Ac > α-2,6-Neu5Ac and low ability to Neu5Gc | Fever, headache, dry cough, loss of appetite fever, nausea, vomiting, diarrhea diseases | Gao et al., 2017 [60] Kappala et al., 2018 [61] |
| Vibrio cholerae | Cholera | B subunit | Neu5Ac and Neu5Gc on GM1 | Watery diarrhea, vomiting, rapid heart rate, low blood pressure | Muanprasat and Chatsudthipong 2013 [62] |
| Pseudomonas aeruginosa | Pneumonia, meningoencephalitis and sepsis | Pili and flagellum Flagellum | asialo-GM1 and asialo-GM2 with GalNAcβ1-4Gal moiety sialyl-Lex epitope with terminal α-2,3-Neu5Ac | Pneumonia and urinary tracts-related symptoms, such as cough, fever, difficulty breathing, swelling | Bassetti et al., 2018 [63]; Krivan et al., 1988 [64]; Xia et al., 2006 [65] |
| Helicobacter pylori | Ulcer, gastritis and gastric cancer | SabA adhesion HpaA | N-acetyllactosamine-based gangliosides with terminal α-2,3-linked Neu5Ac > Neu5Gc and α-2,6-linked Neu5Ac Sialic acid | Ache or burning pain in abdomen, nausea, loss of appetite, bloating | Evans et al., 1988 [66]; Mahdavi 2002 [67]; Roche et al., 2004 [68]; Benktander et al., 2018 [69]; Bennett and Roberts 2005 [70]; Carlsohn et al., 2006 [71] |
| Haemophilus influenzae | Respiratory tract infections: pneumonia, sinusitis; conjunctivitis and otitis media | Lipooligosaccharide (LOS) | Incorporation of α-2,6-Neu5Ac (from human host) and/or dietary Neu5Gc into LOS | Fever, chills, diarrhea, anxiety, difficulty breathing, cough, muscle pain | Ng et al., 2019 [72]; Greiner et al., 2004 [73]; Johnston et al., 2008 [74] |
| Clostridium botulinum | Botulism | Botulin/HA complex | Gangliosides, prefer α-2,3-Neu5Ac > α-2,6-Neu5Ac | Double and blurred vision, dropping eyelids, difficulty swallowing, dry mouth, a thick-feeling tongue | Fujinaga et al., 2000 [75]; Rummel et al., 2007 [76]; Sagane et al., 2016 [77]; Sugawara et al., 2015 [78] |
| Streptococcus pneumoniae | pneumonia, otitis media, sinusitis, meningitis and bronchitis | Sialidases: NanA-C | Cleavage of α-2,3- and α-2,6-Neu5Ac | Fever, cough, shortness of breath, increased sensitivity to light, ear pain, hearing loss | Hatcher et al., 2016 [79]; Coats et al., 2011 [80] |
| SubAB toxin produced by Shiga toxin-produced Escherichia coli | Hemolytic-uremic syndrome (HUS) | B subunit | α-2,3-Neu5Gc > α-2,3-Neu5Ac > α-2,6-Neu5Gc | Extreme fatigue, decreased urination or/and blood in the urine, swelling of the legs, feet, ankles, high blood pressure | Byres et al., 2008 [81]; Seyahian et al., 2017 [82] |
| Plasmodium falciparum | Malaria | EBA-175, EBA-140 | α-2,3-Neu5Ac on GPA and GPC, respectively | Fever, headache, vomiting, tiredness | Tolia et al., 2005 [83]; Rydzak et al., 2015 [84] |
| Plasmodium reichenowi | Malaria in chimpanzee | EBA-140 | Neu5Gc on chimpanzee GPD | Malaria-like symptoms | Zerka et al., 2017 [85] |
| Trypanosoma cruzi | Chagas disease | No ligand, trans-sialidase | Trans-sialidase transfers α-2,3-linked Sias from host to surface mucin-like glycoproteins | Fever, malaise, headache, enlargement of the liver, spleen, lymph nodes | Freire-de-Lima et al., 2012 [86] |
| Entamoeba histolytica | Amoebiasis | Sialoglycoproteins | α-2,3-Neu5Ac | Stomach pain and cramping, loose feces, in severe form bloody feces and fever | Kato et al., 2013 [87] |
| Toxoplasma gondii | Toxoplasmosis | TgMIC1 TgMIC13 | α-2,3-linked Sias 4-O-acetylated Sias and α-2,9-linked Sias | Influenza-like symptoms with swollen lymph glands or muscle aches, damage of brain or eye, reduced vision | Nishikawa et al., 2013 [88] |
1 Human IAV may binds to dietary Neu5Gc.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Burzyńska, P.; Sobala, Ł.F.; Mikołajczyk, K.; Jodłowska, M.; Jaśkiewicz, E. Sialic Acids as Receptors for Pathogens. Biomolecules 2021, 11, 831. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060831
AMA Style
Burzyńska P, Sobala ŁF, Mikołajczyk K, Jodłowska M, Jaśkiewicz E. Sialic Acids as Receptors for Pathogens. Biomolecules. 2021; 11(6):831. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060831
Chicago/Turabian StyleBurzyńska, Patrycja, Łukasz F. Sobala, Krzysztof Mikołajczyk, Marlena Jodłowska, and Ewa Jaśkiewicz. 2021. "Sialic Acids as Receptors for Pathogens" Biomolecules 11, no. 6: 831. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060831
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.