
Modular Assembly of Phosphite Dehydrogenase and Phenylacetone Monooxygenase for Tuning Cofactor Regeneration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Cloning
2.2. Heterologous Production and Purification of Fusion Enzymes
2.3. Formation of Multi-Enzyme Complexes
2.4. Thermostability Assay
2.5. Determination of Enzyme Activity
2.6. Biotransformation
3. Results and Discussion
3.1. Enzymes Production
3.2. Formation of Enzyme Complexes
3.3. Activity and Stability Analyses
3.4. Biotransformations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Muschiol, J.; Peters, C.; Oberleitner, N.; Mihovilovic, M.D.; Bornscheuer, U.T.; Rudroff, F. Cascade catalysis-strategies and challenges en route to preparative synthetic biology. Chem. Commun. 2015, 51, 5798–5811. [Google Scholar] [CrossRef]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-enzymatic cascade reactions: Overview and perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Denard, C.A.; Hartwig, J.F.; Zhao, H. Multistep one-pot reactions combining biocatalysts and chemical catalysts for asymmetric synthesis. ACS Catal. 2013, 3, 2856–2864. [Google Scholar] [CrossRef]
- Sperl, J.M.; Sieber, V. Multienzyme Cascade Reactions—Status and Recent Advances. ACS Catal. 2018, 8, 2385–2396. [Google Scholar] [CrossRef]
- Wang, X.; Saba, T.; Yiu, H.H.P.; Howe, R.F.; Anderson, J.A.; Shi, J. Cofactor NAD(P)H Regeneration Inspired by Heterogeneous Pathways. Chem 2017, 2, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Z.; Zhang, Y.H.; Ren, H.; Wang, Y.L.; Jiang, W.; Fang, B.S. Strategies and perspectives of assembling multi-enzyme systems. Crit. Rev. Biotechnol. 2017, 37, 1024–1037. [Google Scholar] [CrossRef]
- Elleuche, S. Bringing functions together with fusion enzymes—From nature’s inventions to biotechnological applications. Appl. Microbiol. Biotechnol. 2015, 99, 1545–1556. [Google Scholar] [CrossRef]
- Aalbers, F.S.; Fraaije, M.W. Enzyme Fusions in Biocatalysis: Coupling Reactions by Pairing Enzymes. ChemBioChem 2019, 20, 20–28. [Google Scholar] [CrossRef]
- Fürst, M.J.L.J.; Gran-Scheuch, A.; Aalbers, F.S.; Fraaije, M.W. Baeyer-Villiger Monooxygenases: Tunable Oxidative Biocatalysts. ACS Catal. 2019, 9, 11207–11241. [Google Scholar] [CrossRef] [Green Version]
- Torres Pazmiño, D.E.; Snajdrova, R.; Baas, B.J.; Ghobrial, M.; Mihovilovic, M.D.; Fraaije, M.W. Self-sufficient Baeyer-Villiger monooxygenases: Effective coenzyme regeneration for biooxygenation by fusion engineering. Angew. Chem. Int. Ed. 2008, 47, 2275–2278. [Google Scholar] [CrossRef] [Green Version]
- Aalbers, F.S.; Fraaije, M.W. Coupled reactions by coupled enzymes: Alcohol to lactone cascade with alcohol dehydrogenase–cyclohexanone monooxygenase fusions. Appl. Microbiol. Biotechnol. 2017, 101, 7557–7565. [Google Scholar] [CrossRef]
- Chang, C.H.; Rossi, E.A.; Goldenberg, D.M. The dock and lock method: A novel platform technology for building multivalent, multifunctional structures of defined composition with retained bioactivity. Clin. Cancer Res. 2007, 13, 5586–5592. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.A.; Goldenberg, D.M.; Chang, C.H. The dock-and-lock method combines recombinant engineering with site-specific covalent conjugation to generate multifunctional structures. Bioconjug. Chem. 2012, 23, 309–323. [Google Scholar] [CrossRef]
- Newlon, M.G.; Roy, M.; Morikis, D.; Carr, D.W.; Westphal, R.; Scott, J.D.; Jennings, P.A. A novel mechanism of PKA anchoring revealed by solution structures of anchoring complexes. EMBO J. 2001, 20, 1651–1662. [Google Scholar] [CrossRef] [Green Version]
- Kinderman, F.S.; Kim, C.; von Daake, S.; Ma, Y.; Pham, B.Q.; Spraggon, G.; Xuong, N.H.; Jennings, P.A.; Taylor, S.S. A Dynamic Mechanism for AKAP Binding to RII Isoforms of cAMP-Dependent Protein Kinase. Mol. Cell 2006, 24, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Sivadas, P.; Dienes, J.M.; Maurice, M.S.; Meek, W.D.; Yang, P. A flagellar a-kinase anchoring protein with two amphipathic helices forms a structural scaffold in the radial spoke complex. J. Cell Biol. 2012, 199, 639–651. [Google Scholar] [CrossRef] [Green Version]
- Calejo, A.I.; Taskén, K. Targeting protein-protein interactions in complexes organized by A kinase anchoring proteins. Front. Pharmacol. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Blumenthal, R.D.; Hansen, H.J.; Goldenberg, D.M. Inhibition of adhesion, invasion, and metastasis by antibodies targeting CEACAM6 (NCA-90) and CEACAM5 (carcinoembryonic antigen). Cancer Res. 2005, 65, 8809–8817. [Google Scholar] [CrossRef] [Green Version]
- Stein, R.; Qu, Z.; Chen, S.; Rosario, A.; Shi, V.; Hayes, M.; Horak, I.D.; Hansen, H.J.; Goldenberg, D.M. Characterization of A New Humanized Anti-CD20 Monoclonal Antibody, IMMU-106, and Its Use in Combination with the Humanized Anti-CD22 Antibody, Epratuzumab, for the Therapy of Non-Hodgkin’s Lymphoma. Clin. Cancer Res. 2004, 10, 2868–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, E.A.; Goldenberg, D.M.; Cardillo, T.M.; Stein, R.; Chang, C.H. CD20-targeted tetrameric interferon-α, a novel and potent immunocytokine for the therapy of B-cell lymphomas. Blood 2009, 114, 3864–3871. [Google Scholar] [CrossRef]
- Rossi, E.A.; Rossi, D.L.; Stein, R.; Goldenberg, D.M.; Chang, C.H. A bispecific antibody-IFNα2b immunocytokine targeting CD20 and HLA-DR is highly toxic to human lymphoma and multiple myeloma cells. Cancer Res. 2010, 70, 7600–7609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Ma, T.; Liu, M.; Qu, J.; Liu, Z.; Zhang, H.; Shi, B.; Fu, S.; Ma, J.; Lai, L.T.F.; et al. Modular enzyme assembly for enhanced cascade biocatalysis and metabolic flux. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Torres Pazmiño, D.E.; Riebel, A.; De Lange, J.; Rudroff, F.; Mihovilovic, M.D.; Fraaije, M.W. Efficient biooxidations catalyzed by a new generation of self-sufficient Baeyer-Villiger monooxygenases. ChemBioChem 2009, 10, 2595–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forneris, F.; Orru, R.; Bonivento, D.; Chiarelli, L.R.; Mattevi, A. ThermoFAD, a Thermofluor®-adapted flavin ad hoc detection system for protein folding and ligand binding. FEBS J. 2009, 276, 2833–2840. [Google Scholar] [CrossRef] [PubMed]
- Gran-Scheuch, A.; Trajkovic, M.; Parra, L.; Fraaije, M.W. Mining the Genome of Streptomyces leeuwenhoekii: Two New Type I Baeyer–Villiger Monooxygenases From Atacama Desert. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Beyer, N.; Kulig, J.K.; Bartsch, A.; Hayes, M.A.; Janssen, D.B.; Fraaije, M.W. P450BM3fused to phosphite dehydrogenase allows phosphite-driven selective oxidations. Appl. Microbiol. Biotechnol. 2017, 101, 2319–2331. [Google Scholar] [CrossRef] [Green Version]
- Aalbers, F.S.; Fraaije, M.W. Design of Artificial Alcohol Oxidases: Alcohol Dehydrogenase–NADPH Oxidase Fusions for Continuous Oxidations. ChemBioChem 2019, 20, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Booth, W.T.; Schlachter, C.R.; Pote, S.; Ussin, N.; Mank, N.J.; Klapper, V.; Offermann, L.R.; Tang, C.; Hurlburt, B.K.; Chruszcz, M. Impact of an N-terminal polyhistidine tag on protein thermal stability. ACS Omega 2018, 3, 760–768. [Google Scholar] [CrossRef] [PubMed]




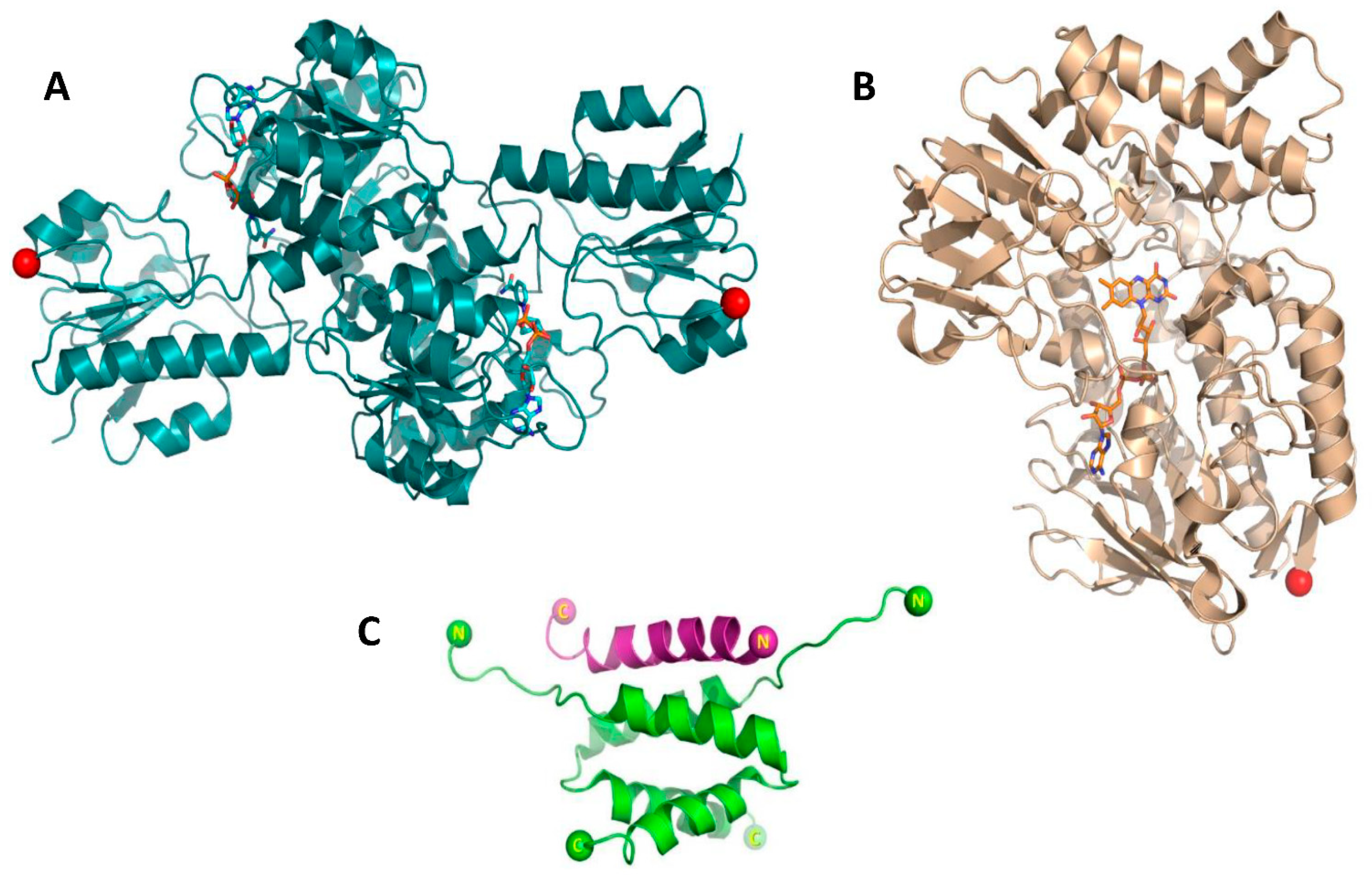{kind=link}
{kind=link}
{kind=link}
| Enzyme Variant a | Linker | C-Terminus | Mw (kDa) b |
|---|---|---|---|
| PAMOA | (GGGGS) × 3 | RIAD | 67 |
| PAMOA2 | (GGGGS) × 3 | RIAD–RIAD | 70 |
| PAMOI | (GGGGS) × 3 | RIDD | 70 |
| PTDHA | (GGGGS) × 3 | RIAD | 42 |
| PTDHA2 | (GGGGS) × 3 | RIAD–RIAD | 45 |
| PTDHI | (GGGGS) × 3 | RIDD | 45 |
| PAMO | PTDH | |||
|---|---|---|---|---|
| Enzyme | kobs (s−1) | Tmapp (°C) a | kobs (s−1) | Tmapp (°C) b |
| PTDHA–PAMOI | 0.85 ± 0.01 | 59.0 | 4.18 ± 0.16 | 53.0 |
| PTDHA2–PAMOI | 0.86 ± 0.04 | 59.0 | 2.79 ± 0.15 | 51.5 |
| PAMOA–PTDHI | 1.08 ± 0.01 | 59.0 | 2.07 ± 0.02 | 56.5 |
| PAMOA2–PTDHI | 1.06 ± 0.08 | 59.5 | 2.12 ± 0.02 | 56.5 |
| PTDH–PAMO | 1.13 ± 0.01 | 60.0 | 4.65 ± 0.04 | 59.0 |
| PAMO | 0.58 ± 0.01 | 60.5 | - | - |
| PTDH | - | - | 2.42 ± 0.04 | 64.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purwani, N.N.; Martin, C.; Savino, S.; Fraaije, M.W. Modular Assembly of Phosphite Dehydrogenase and Phenylacetone Monooxygenase for Tuning Cofactor Regeneration. Biomolecules 2021, 11, 905. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060905
Purwani NN, Martin C, Savino S, Fraaije MW. Modular Assembly of Phosphite Dehydrogenase and Phenylacetone Monooxygenase for Tuning Cofactor Regeneration. Biomolecules. 2021; 11(6):905. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060905
Chicago/Turabian StylePurwani, Ni Nyoman, Caterina Martin, Simone Savino, and Marco W. Fraaije. 2021. "Modular Assembly of Phosphite Dehydrogenase and Phenylacetone Monooxygenase for Tuning Cofactor Regeneration" Biomolecules 11, no. 6: 905. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11060905