
The Role of COA6 in the Mitochondrial Copper Delivery Pathway to Cytochrome c Oxidase


Department of Biochemistry and Biophysics, Texas A&M University, College Station, TX 77843, USA
*
Author to whom correspondence should be addressed.
Biomolecules 2022, 12(1), 125; https://0-doi-org.brum.beds.ac.uk/10.3390/biom12010125
Submission received: 16 December 2021
/
Revised: 9 January 2022
/
Accepted: 10 January 2022
/
Published: 13 January 2022
(This article belongs to the Special Issue Insights in the Structure and Functions of Mitochondrial Proteins and Metalloproteins)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Copper is essential for the stability and activity of cytochrome c oxidase (CcO), the terminal enzyme of the mitochondrial respiratory chain. Copper is bound to COX1 and COX2, two core subunits of CcO, forming the CuB and CuA sites, respectively. Biogenesis of these two copper sites of CcO occurs separately and requires a number of evolutionarily conserved proteins that form the mitochondrial copper delivery pathway. Pathogenic mutations in some of the proteins of the copper delivery pathway, such as SCO1, SCO2, and COA6, have been shown to cause fatal infantile human disorders, highlighting the biomedical significance of understanding copper delivery mechanisms to CcO. While two decades of studies have provided a clearer picture regarding the biochemical roles of SCO1 and SCO2 proteins, some discrepancy exists regarding the function of COA6, the new member of this pathway. Initial genetic and biochemical studies have linked COA6 with copper delivery to COX2 and follow-up structural and functional studies have shown that it is specifically required for the biogenesis of the CuA site by acting as a disulfide reductase of SCO and COX2 proteins. Its role as a copper metallochaperone has also been proposed. Here, we critically review the recent literature regarding the molecular function of COA6 in CuA biogenesis.
Keywords:
mitochondria; copper; cytochrome c oxidase; COA6; COX2; CuA site; metallochaperone; disulfide reductase1. Cytochrome c Oxidase
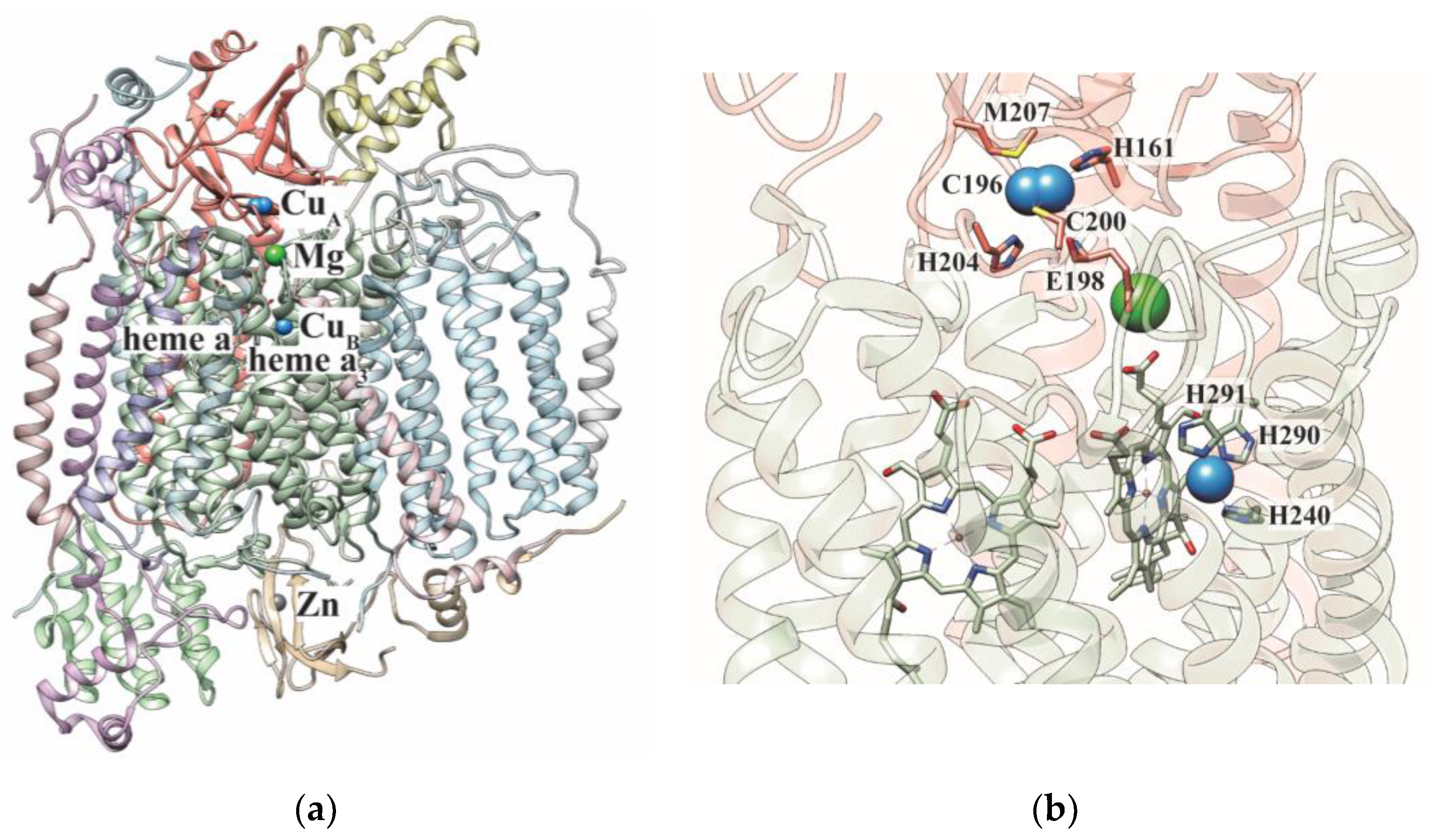
Cytochrome c oxidase (CcO) is the terminal enzyme of the mitochondrial respiratory chain (MRC) that catalyzes the reduction of molecular oxygen to water using electrons from cytochrome c [1]. CcO is a multimeric protein complex present in the inner mitochondrial membrane and is composed of three core subunits encoded by the mitochondrial DNA and numerous (up to 11) accessory subunits encoded by the nuclear DNA (Figure 1a) [2,3]. Two of the core subunits of CcO, COX1 and COX2, contain metal cofactors that participate in electron transfer and catalysis (Figure 1b) [4,5]. The COX2 subunit contains two copper ions forming the binuclear CuA site and the COX1 subunit contains heme a, heme a3, and a single copper ion, which together with heme a3 forms the CuB/heme a3 site (Figure 1b). Together, these cofactors form the “molecular wire” that transfers electrons from reduced cytochrome c to molecular oxygen. The process of electron transport is coupled to proton pumping from the matrix to the intermembrane space (IMS), an activity carried out by other complexes (I and III) of the MRC as well. The proton pumping activity of these MRC complexes generates a proton gradient across the inner mitochondrial membrane, which drives mitochondrial energy production in the form of adenosine triphosphate (ATP).
The copper requirement of CcO necessitates its delivery to the mitochondria [6]. With copper being a highly reactive metal, with its ability to generate deleterious reactive oxygen species and displace other metals from their native coordination sites [6,7,8,9,10], cells employ special strategies to transport and insert copper into target proteins. This involves small copper-binding molecules as well as proteins termed metallochaperones, which bind copper and insert it into its target proteins via specific protein–protein interactions [11].
2. The Mitochondrial Copper Delivery Pathway
Cytosolic copper is delivered to the mitochondrial matrix in a ligand-bound form, called copper-ligand (CuL), though the molecular identity of this ligand remains elusive [12]. Although copper insertion into the CcO subunits occurs in the mitochondrial IMS (Figure 2), cytosolic copper is first transported into the mitochondrial matrix via the mitochondrial phosphate carrier protein Pic2 in yeast and its mammalian homolog SLC25A3 [13,14]. Pic2-deleted yeast does not exhibit a robust copper-deficiency phenotype [13,15], suggesting alternate mechanism(s) of copper transport into the mitochondria. Indeed, it was later shown that mitochondrial iron transporter, Mrs3, could also transport copper to the mitochondrial matrix [16]. The mitochondrial matrix copper pool is the main source of copper ions that are inserted into the CcO subunits [12]. Thus, mobilization of copper from the mitochondrial matrix to the IMS for its delivery to CcO copper sites requires its efflux back to the IMS through a yet unidentified transporter [17]. Once in the IMS, a number of evolutionarily conserved proteins participate in its insertion into the COX1 and COX2 subunits of CcO [18] (Figure 2). Genetic and biochemical studies have shown that IMS-localized COX17 is the common source of copper for the COX1- and COX2-specific metallochaperones [18]. The next steps in copper delivery to CcO are a modular process, which involves the biogenesis of CuA and CuB as two separate modules on COX2 and COX1, respectively (Figure 2) [19,20]. Before copper insertion into the CuB site, heme cofactors are added on to the COX1 subunit, which is inserted into the inner mitochondrial membrane [18]. Similarly, copper insertion into the CuA site requires the membrane insertion of COX2 and the subsequent translocation of its copper-binding domain to the mitochondrial IMS [18]. This topology of COX1 and COX2 copper-binding domains necessitates the IMS localization of the CcO assembly factors involved in copper delivery (Figure 2). Within the IMS, SCO1, SCO2, and COA6 are involved in copper delivery to the COX2 module whereas COX11 and COX19 are involved in copper delivery to the COX1 module (Figure 2) [18,21,22,23,24,25,26,27,28,29,30]. The localization of the copper delivery proteins in the oxidizing environment of the IMS is reminiscent of prokaryotes, where copper delivery into the CcO subunits occurs in the periplasmic space.
The precise molecular functions of metallochaperones COX17, SCO1, and COX11, have been determined and they have been shown to transfer copper to CcO subunits in a bucket-brigade fashion [18,25,26,27,29,30]. Specifically, COX17 receives copper in the IMS via an unknown mechanism and transfers it to COX11 and SCO1, which then metallates copper sites on COX1 and COX2, respectively (Figure 2) [25,26,27,29,30]. In addition to these metallochaperones, SCO2, COA6, and COX19 have also been shown to be a part of the copper delivery pathway in the IMS [21,23,24,28,29]. SCO2, which is a paralog of SCO1, also receives copper from COX17 and acts as a copper-dependent disulfide reductase of COX2, a reaction necessary for its metallation by SCO1 [29]. COX19 has been shown to play a role in maintaining the redox state of COX11 and facilitating copper delivery to the COX1 subunit [28]. COA6 facilitates the assembly of the CuA site of COX2 [21,22,23,24], and its precise biochemical function in this process is the focus of this review.
3. History of Discovery of COA6
COA6, originally named C1ORF31 in humans and Ymr244C-A in yeast, was first identified as a putative CcO assembly factor based on an iterative orthology prediction [31]. Subsequently, it was found in a proteomic survey of mitochondrial IMS proteins in the yeast Saccharomyces cerevisiae and was renamed Coa6 (Cytochrome Oxidase Assembly factor 6) because of its requirement for CcO assembly [32]. Although these studies were the first to link Coa6 to CcO biogenesis, its precise role in the process remained unclear. Based on its sub-mitochondrial localization, human genetics, and protein sequence, we predicted the role of Coa6 in the copper delivery pathway to CcO [21]. Three key findings from our group confirmed this prediction. First, copper supplementation in the yeast growth media rescued respiratory growth and CcO assembly of coa6∆ cells, providing the first link between Coa6 and the mitochondrial copper delivery pathway to CcO [21]. Second, a comprehensive genetic interaction study uncovered a synthetic lethal interaction between yeast coa6∆ and sco2∆ mutants, a finding that placed Coa6 in the copper delivery to the CuA site [23]. Third, co-immunoprecipitation of Coa6 identified specific interaction of Coa6 with proteins involved in the CuA site biogenesis, including Sco1, Sco2, and Cox2 [23]. We further showed that heterologous expression of human COA6 was able to rescue the respiratory growth of yeast coa6∆ indicating that the function of Coa6 is evolutionarily conserved [23]. Indeed, a subsequent study with COA6 patient fibroblasts recapitulated copper-mediated rescue of COX2 levels [33]. Consistent with our studies with the yeast model system, two other groups independently demonstrated the role of COA6 in COX2 biogenesis in mammalian cells through protein–protein interaction studies [22,34]. Specifically, both groups found that human COA6 interacts with the newly synthesized COX2 [22,34], though there was some discrepancy between these two studies regarding the interaction of COA6 with SCO proteins, with one group reporting COA6:SCO1 interaction [22] and the other group reporting COA6:SCO2 interaction in vivo [34]. Our recent in vitro studies with purified human proteins have shown that COA6 can interact with both SCO1 and SCO2; however, when all three proteins are present in the reaction mixture, COA6 preferentially interacts with SCO1 [24]. These studies together, unequivocally place COA6 in the final steps of CuA site biogenesis.
4. Structure of COA6
The structure of the human recombinant COA6 protein has been determined by both X-ray crystallography and nuclear magnetic resonance (NMR) spectroscopy. Both the structures show COA6 as a helical protein with coiled-coil-helix-coiled-coil-helix (CHCH) fold domain [24,35,36]. COA6 contains three α-helices where the first two helices—helix 1 (H1) and helix 2 (H2)—form a CHCH domain stabilized by two disulfide bonds between the cysteine pairs of its CX9C-CX10C motif (Figure 3a,b) [24,36]. Despite these similarities between the solution and crystal structures, a number of differences exist, as described in detail in a recent review [35]. Briefly, the length of the H1 and H2 helices in the crystal structure is significantly longer than those in the solution structure (Figure 3a,b). In the solution structure, helix 3 (H3) is broken into a shorter fourth helix and the orientation of H3 is also different between these two structures (Figure 3a,b) [24,36]. In the crystal structure, the H3 of one monomer of COA6 interacts with the other, thereby stabilizing the dimeric state whereas in the solution structure this helix is present at the opposite face of the CHCH domain (Figure 3a,b) [24,36]. Notably, the structure of COA6 predicted through AlphaFold software is more similar to the crystal structure of COA6. We speculate that the indicated differences between the crystal structure and the solution structure could be due to the dynamic nature of the protein and its concentration dependent oligomerization involving H3 [24]. In support of this idea, NMR-based analysis of the interaction of COA6 with SCO1 reveals that the residues of H3 in COA6 are also involved in its interaction with SCO1 [24]. Therefore, it appears that this third helix is dynamic and plays a crucial role not only in the dimerization of COA6 but also in its interaction with its target proteins. Indeed, deletion of H3 abrogated COA6 function in vivo [24].
The proteins containing the CHCH domain have previously been shown to perform diverse functions in the cell [27,37,38]. For example, two well-characterized CHCH-domain containing proteins, COX17 and MIA40, function as metallochaperone and thiol-disulfide oxidoreductase, respectively. In the case of COX17, two cysteines at the N-terminal region outside the CHCH domain bind copper (Figure 3c), which is necessary for its role as a copper metallochaperone [27,39]. MIA40 is part of the protein import machinery of the mitochondrial IMS that functions as a thiol-disulfide oxidoreductase. It contains redox-active cysteines in its CPC motif present at the N-terminal region of the protein outside its CHCH domain (Figure 3d) [38]. Based on the known functions of these CHCH-domain containing proteins, COA6 can be predicted to act as a metallochaperone or a disulfide reductase.
5. Is COA6 a Copper Metallochaperone?
Metallochaperones are defined as proteins that bind metal ions and deliver them directly to target protein(s) via specific protein–protein interactions [11]. Based on this definition, if COA6 is a copper metallochaperone then it is expected to bind copper, physically interact with its target protein(s), and insert copper into the metal-binding sites on the target protein(s). Below, we critically evaluate these criteria for the potential metallochaperone role of COA6.
There is discrepancy in the literature regarding the copper-binding ability of COA6, its stoichiometry of binding, and the physiological relevance of copper binding. One study showed that purified GST-Coa6 fusion protein could bind copper at a mass ratio of about 8.5 ng copper per µg of protein [34]. This would equate to copper:Coa6 molar stoichiometry of about 5:1. In the same study, GST-Sco1 and GST-Sco2, proteins known to bind copper at a 1:1 molar ratio, were used as controls and were shown to have copper: protein molar stoichiometries of about 6:1 and 5:1, respectively. Clearly, the experimental conditions used in this study were skewed towards excess copper binding. Another group performed a competitive in vitro copper-binding assay by incubating purified human COA6 with Cu(I)-bathocuproine disulfonic acid (BCS) complex and determined that COA6 binds Cu with a very high affinity (Kd of ~10−17 M) [22]. They further showed that COA6 binds copper in a 1:1 stoichiometry and mutagenesis of the first and fourth cysteines of COA6 completely eliminates copper binding [36]. In contrast to these studies, we determined that purified recombinant human COA6 did not contain any bound copper [24]. Nevertheless, the reconstitution of purified COA6 with copper under reducing conditions, as described in the previous studies [22,36], did result in copper binding [24]. However, the ability of COA6 to bind copper in vitro could be an experimental artifact since copper being thiophilic, is expected to bind to the cysteine residues of COA6 that are forcibly reduced. In fact, a similar proposal was made for Cmc1, another twin CX9C motif-containing mitochondrial IMS protein, which was shown to bind copper upon in vitro reconstitution [40]. Later, it was found to have a copper-independent role in the stabilization of the COX1-intermediate [37]. Thus, the Cmc1 work serves as a cautionary example of using an in vitro copper reconstitution assay as a metric for linking the function of twin CX9C motif-containing proteins to copper metabolism. Moreover, the concentration of free copper ions inside the cell is virtually zero since all copper is either protein bound or is sequestered by non-proteinaceous anionic ligands [12,41]. Therefore, the in vitro reconstitution studies do not mimic in vivo conditions. While demonstrating in vivo binding of copper to protein is challenging, our work using online tandem liquid chromatography-inductively coupled plasma-mass spectrometry (LC-ICP-MS)-based experiments suggested that Coa6 is not bound to copper in vivo [24].
Several additional observations call into question the metallochaperone role of COA6. First, COA6 lacks the canonical copper-binding residues present in other CHCH-domain containing copper metallochaperones such as COX17 (Figure 3a–c). Although it was proposed that copper binds through the first and fourth cysteines of the CX9C-CX10C motif of COA6 [36], the cysteines of the CX9C-CX9-10C motif have not been shown to bind copper in other well-characterized proteins such as COX17, COX6B, and MIA40. Notably, none of the structures reported for COA6 are in its copper-bound form [24,36]. Second, if COA6 acts as a copper metallochaperone in copper delivery to COX2, its deletion should phenocopy cox17∆ or sco1∆ yeast mutants because deletion mutants of all metallochaperones involved in copper delivery to CcO display strong respiratory deficient phenotypes. In contrast, coa6∆ exhibits much milder respiratory phenotypes including partially reduced respiratory growth, respiration, and CcO abundance [21,23]. Third, while protein–protein interaction studies in both yeast and mammalian cells have demonstrated interaction of COA6 with SCO1, SCO2, and COX2, currently no studies have shown the direct transfer of copper from COA6 to its interacting partners [22,23,34]. Without this direct evidence, it is too premature to call COA6 a copper metallochaperone. Taken together, these studies argue against a metallochaperone role for COA6 in vivo.
6. Is COA6 a Disulfide Reductase?
A prerequisite for copper binding by the cysteine ligands of the cuproproteins is that they must be in a reduced (sulfhydryl) state. The IMS of mitochondria has a reduction potential of −255 mV [42] and the reduction potential of copper-binding cysteines of SCO1 and COX2 were shown to be about −280 mV and −290 mV, respectively [29,43]. Now, assuming thermodynamic equilibrium, we would expect that these cysteines would be prone to oxidation in the IMS. This would pose a problem in copper transfer to the CuA site and would require the action of disulfide reductase(s) that can reduce these copper-binding cysteines. Bacteria face a similar problem because the copper-binding cysteine residues in the bacterial CuA site lie on the periplasmic side of the cytosolic membrane, which is also an oxidizing environment. These copper-coordinating cysteines are prone to form disulfide bonds, which must be reduced prior to copper insertion. The periplasmic thioredoxin, TlpA, was shown to perform this role by reducing the copper-binding cysteines of both SCOI and COXB, the bacterial homologs of SCO1 and COX2, respectively [44]. As described below, recent studies show that COA6 performs an analogous role in the eukaryotic mitochondria by reducing the disulfides of both SCO1 and COX2 [24].
A number of observations hinted at the potential redox role of COA6. First, the respiratory growth phenotype of yeast coa6∆ mutant was suppressed under reducing conditions [24]. Second, yeast COA6 exhibited a strong synthetic lethal interaction with SCO2, which has already been shown to act as a disulfide reductase in vitro and in vivo [23,45]. Third, CHCH domain-containing proteins have been proposed to function as redox proteins [46]. In line with these observations, it was shown that COA6 can reduce the disulfides of SCO1, SCO2, and COX2, in vitro [24]. Importantly, this reaction is thermodynamically favored since the reduction potential values of COA6, variously reported as −330 mV [24] or −349 mV [36], is lower than that of SCO1 (−280 mV), SCO2 (<−300 mV), and COX2 (−290 mV) [24]. Consistent with this in vitro data, it was found that COA6 is required to maintain the cysteines of SCO1 and SCO2 in the reduced form in vivo [24,47]. Thus, there is strong experimental evidence in support of the disulfide reductase activity of COA6.
Despite this evidence, the molecular basis of disulfide reductase activity of COA6 is not known and some observations are not consistent with this role. For example, although COA6 has a CHCH domain similar to that seen in another thiol-disulfide oxidoreductase, MIA40, the redox-active CPC motif of MIA40 is absent in COA6. The cysteine residues of CX9C-CX10C motif of COA6 are likely candidates for the disulfide reductase activity. However, there is no evidence that the other CX9C-CX9-10C motif-containing proteins, such as MIA40 or COX17, exhibit redox activity using the cysteines of the CX9C-CX9-10C motif. Further, since COA6 has been shown to be a MIA40 substrate [32], at least one pair of its cysteines will be in the oxidized form upon its import into the mitochondrial IMS, making them incapable of redox cycling. Contradictory evidences exist regarding the identity of these potential redox-active cysteine residues. NMR studies on COA6 showed that the second and third cysteine pairs are capable of redox cycling [24]. However, a recent study showed that the first and fourth cysteine pairs of COA6 are redox active with a reduction potential of −349 mV [36]. In addition to this discrepancy, the NMR chemical shift perturbation data on the interaction between SCO1 and COA6 showed that only helix 3 of COA6 is involved in the intermolecular interaction with SCO1. None of the cysteines of COA6 or the copper-binding cysteines of SCO1 were shown to be involved in this interaction. Importantly, the source of electrons used by COA6 for its disulfide reductase activity is unknown. Taken together, the molecular mechanism by which COA6 influences the redox state of SCO1 and COX2 still remains unclear.
7. Concluding Remarks
The delivery and insertion of copper cofactors in the mitochondrial CcO requires a number of “assembly factors”. Elucidating the biochemical function of these CcO assembly factors has remained a major bottleneck in our understanding of the biogenesis of CcO, with the role of COA6 in CuA site formation being a case in point. A recent review article debated the potential role of COA6 as a metallochaperone or a thiol-disulfide oxidoreductase (disulfide reductase) in the copper delivery process [35]. Upon critical analysis of literature, we find that the current experimental data favors the disulfide reductase activity of COA6 and leads to a model where COA6 acts on its client proteins—COX2 and SCO1—by reducing the disulfide bond between their copper-coordinating cysteines to allow for copper binding (Figure 4). Future studies are required to determine the COA6 residues that participate in the disulfide reductase activity, the mechanism of its substrate specificity, and the structural basis of its activity. Addressing these issues could establish COA6 as a paradigm for understanding the biochemical roles of other CX9C-CX9-10C motif containing proteins in CcO biogenesis.
Author Contributions
The original draft of the manuscript was written by A.B.S., which was then jointly edited and revised by A.B.S., V.M.G. and V.M.G. was responsible for project administration and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institutes of Health grant R01GM111672 to V.M.G.
Acknowledgments
We thank Paul A. Lindahl, Jae-Hyun Cho, and Sagnika Ghosh for providing critical comments during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of Biochemistry: Life at the Molecular Level, 5th ed.; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Hartley, A.M.; Lukoyanova, N.; Zhang, Y.; Cabrera-Orefice, A.; Arnold, S.; Meunier, B.; Pinotsis, N.; Marechal, A. Structure of yeast cytochrome c oxidase in a supercomplex with cytochrome bc1. Nat. Struct. Mol. Biol. 2019, 26, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Zong, S.; Wu, M.; Gu, J.; Liu, T.; Guo, R.; Yang, M. Structure of the intact 14-subunit human cytochrome c oxidase. Cell Res. 2018, 28, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. Structures of metal sites of oxidized bovine heart cytochrome c oxidase at 2.8 A. Science 1995, 269, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.E.; Nevitt, T.; Thiele, D.J. Mechanisms for copper acquisition, distribution and regulation. Nat. Chem. Biol. 2008, 4, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, A.W.; Osman, D.; Robinson, N.J. Metal preferences and metallation. J. Biol. Chem. 2014, 289, 28095–28103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irving, H.; Williams, R.J.P. The Stability of Transition-Metal Complexes. J. Chem. Soc. 1953, 3192–3210. [Google Scholar] [CrossRef]
- Rosenzweig, A.C. Metallochaperones: Bind and deliver. Chem. Biol. 2002, 9, 673–677. [Google Scholar] [CrossRef] [Green Version]
- Cobine, P.A.; Ojeda, L.D.; Rigby, K.M.; Winge, D.R. Yeast contain a non-proteinaceous pool of copper in the mitochondrial matrix. J. Biol. Chem. 2004, 279, 14447–14455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vest, K.E.; Leary, S.C.; Winge, D.R.; Cobine, P.A. Copper import into the mitochondrial matrix in Saccharomyces cerevisiae is mediated by Pic2, a mitochondrial carrier family protein. J. Biol. Chem. 2013, 288, 23884–23892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulet, A.; Vest, K.E.; Maynard, M.K.; Gammon, M.G.; Russell, A.C.; Mathews, A.T.; Cole, S.E.; Zhu, X.; Phillips, C.B.; Kwong, J.Q.; et al. The mammalian phosphate carrier SLC25A3 is a mitochondrial copper transporter required for cytochrome c oxidase biogenesis. J. Biol. Chem. 2018, 293, 1887–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soma, S.; Latimer, A.J.; Chun, H.; Vicary, A.C.; Timbalia, S.A.; Boulet, A.; Rahn, J.J.; Chan, S.S.L.; Leary, S.C.; Kim, B.E.; et al. Elesclomol restores mitochondrial function in genetic models of copper deficiency. Proc. Natl. Acad. Sci. USA 2018, 115, 8161–8166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vest, K.E.; Wang, J.; Gammon, M.G.; Maynard, M.K.; White, O.L.; Cobine, J.A.; Mahone, W.K.; Cobine, P.A. Overlap of copper and iron uptake systems in mitochondria in Saccharomyces cerevisiae. Open. Biol. 2016, 6, 150223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, Z.N.; Cobine, P.A.; Leary, S.C. The mitochondrion: A central architect of copper homeostasis. Metallomics 2017, 9, 1501–1512. [Google Scholar] [CrossRef]
- Timon-Gomez, A.; Nyvltova, E.; Abriata, L.A.; Vila, A.J.; Hosler, J.; Barrientos, A. Mitochondrial cytochrome c oxidase biogenesis: Recent developments. Semin. Cell Dev. Biol. 2018, 76, 163–178. [Google Scholar] [CrossRef]
- McStay, G.P.; Su, C.H.; Thomas, S.M.; Xu, J.T.; Tzagoloff, A. Characterization of assembly intermediates containing subunit 1 of yeast cytochrome oxidase. J. Biol. Chem. 2013, 288, 26546–26556. [Google Scholar] [CrossRef] [Green Version]
- Franco, L.V.R.; Su, C.H.; McStay, G.P.; Yu, G.J.; Tzagoloff, A. Cox2p of yeast cytochrome oxidase assembles as a stand-alone subunit with the Cox1p and Cox3p modules. J. Biol. Chem. 2018, 293, 16899–16911. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Trivedi, P.P.; Timbalia, S.A.; Griffin, A.T.; Rahn, J.J.; Chan, S.S.; Gohil, V.M. Copper supplementation restores cytochrome c oxidase assembly defect in a mitochondrial disease model of COA6 deficiency. Hum. Mol. Genet. 2014, 23, 3596–3606. [Google Scholar] [CrossRef]
- Stroud, D.A.; Maher, M.J.; Lindau, C.; Vogtle, F.N.; Frazier, A.E.; Surgenor, E.; Mountford, H.; Singh, A.P.; Bonas, M.; Oeljeklaus, S.; et al. COA6 is a mitochondrial complex IV assembly factor critical for biogenesis of mtDNA-encoded COX2. Hum. Mol. Genet. 2015, 24, 5404–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Pratt, A.T.; Soma, S.; Theriault, S.G.; Griffin, A.T.; Trivedi, P.P.; Gohil, V.M. Mitochondrial disease genes COA6, COX6B and SCO2 have overlapping roles in COX2 biogenesis. Hum. Mol. Genet. 2016, 25, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soma, S.; Morgada, M.N.; Naik, M.T.; Boulet, A.; Roesler, A.A.; Dziuba, N.; Ghosh, A.; Yu, Q.; Lindahl, P.A.; Ames, J.B.; et al. COA6 Is Structurally Tuned to Function as a Thiol-Disulfide Oxidoreductase in Copper Delivery to Mitochondrial Cytochrome c Oxidase. Cell Rep. 2019, 29, 4114–4126.e4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiser, L.; Di Valentin, M.; Hamer, A.G.; Hosler, J.P. Cox11p is required for stable formation of the Cu(B) and magnesium centers of cytochrome c oxidase. J. Biol. Chem. 2000, 275, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Carr, H.S.; George, G.N.; Winge, D.R. Yeast Cox11, a protein essential for cytochrome c oxidase assembly, is a Cu(I)-binding protein. J. Biol. Chem. 2002, 277, 31237–31242. [Google Scholar] [CrossRef] [Green Version]
- Horng, Y.C.; Cobine, P.A.; Maxfield, A.B.; Carr, H.S.; Winge, D.R. Specific copper transfer from the Cox17 metallochaperone to both Sco1 and Cox11 in the assembly of yeast cytochrome C oxidase. J. Biol. Chem. 2004, 279, 35334–35340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, M.; Woellhaf, M.W.; Bohnert, M.; van der Laan, M.; Sommer, F.; Jung, M.; Zimmermann, R.; Schroda, M.; Herrmann, J.M. Redox-regulated dynamic interplay between Cox19 and the copper-binding protein Cox11 in the intermembrane space of mitochondria facilitates biogenesis of cytochrome c oxidase. Mol. Biol. Cell 2015, 26, 2385–2401. [Google Scholar] [CrossRef]
- Morgada, M.N.; Abriata, L.A.; Cefaro, C.; Gajda, K.; Banci, L.; Vila, A.J. Loop recognition and copper-mediated disulfide reduction underpin metal site assembly of CuA in human cytochrome oxidase. Proc. Natl. Acad. Sci. USA 2015, 112, 11771–11776. [Google Scholar] [CrossRef] [Green Version]
- Leary, S.C.; Kaufman, B.A.; Pellecchia, G.; Guercin, G.H.; Mattman, A.; Jaksch, M.; Shoubridge, E.A. Human SCO1 and SCO2 have independent, cooperative functions in copper delivery to cytochrome c oxidase. Hum. Mol. Genet. 2004, 13, 1839–1848. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, R.; Wanschers, B.F.; Cuypers, T.D.; Esseling, J.J.; Riemersma, M.; van den Brand, M.A.; Gloerich, J.; Lasonder, E.; van den Heuvel, L.P.; Nijtmans, L.G.; et al. Iterative orthology prediction uncovers new mitochondrial proteins and identifies C12orf62 as the human ortholog of COX14, a protein involved in the assembly of cytochrome c oxidase. Genome Biol. 2012, 13, R12. [Google Scholar] [CrossRef]
- Vogtle, F.N.; Burkhart, J.M.; Rao, S.; Gerbeth, C.; Hinrichs, J.; Martinou, J.C.; Chacinska, A.; Sickmann, A.; Zahedi, R.P.; Meisinger, C. Intermembrane space proteome of yeast mitochondria. Mol. Cell Proteom. 2012, 11, 1840–1852. [Google Scholar] [CrossRef] [Green Version]
- Baertling, F.; van den Brand, M.A.M.; Hertecant, J.L.; Al-Shamsi, A.; van den Heuvel, L.P.; Distelmaier, F.; Mayatepek, E.; Smeitink, J.A.; Nijtmans, L.G.; Rodenburg, R.J. Mutations in COA6 cause cytochrome c oxidase deficiency and neonatal hypertrophic cardiomyopathy. Hum. Mutat. 2015, 36, 34–38. [Google Scholar] [CrossRef]
- Pacheu-Grau, D.; Bareth, B.; Dudek, J.; Juris, L.; Vogtle, F.N.; Wissel, M.; Leary, S.C.; Dennerlein, S.; Rehling, P.; Deckers, M. Cooperation between COA6 and SCO2 in COX2 maturation during cytochrome c oxidase assembly links two mitochondrial cardiomyopathies. Cell Metab. 2015, 21, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Maghool, S.; Ryan, M.T.; Maher, M.J. What Role Does COA6 Play in Cytochrome C Oxidase Biogenesis: A Metallochaperone or Thiol Oxidoreductase, or Both? Int. J. Mol. Sci. 2020, 21, 6983. [Google Scholar] [CrossRef] [PubMed]
- Maghool, S.; Cooray, N.D.G.; Stroud, D.A.; Aragao, D.; Ryan, M.T.; Maher, M.J. Structural and functional characterization of the mitochondrial complex IV assembly factor Coa6. Life Sci. Alliance 2019, 2. [Google Scholar] [CrossRef]
- Bourens, M.; Barrientos, A. A CMC1-knockout reveals translation-independent control of human mitochondrial complex IV biogenesis. EMBO Rep. 2017, 18, 477–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banci, L.; Bertini, I.; Cefaro, C.; Ciofi-Baffoni, S.; Gallo, A.; Martinelli, M.; Sideris, D.P.; Katrakili, N.; Tokatlidis, K. MIA40 is an oxidoreductase that catalyzes oxidative protein folding in mitochondria. Nat. Struct. Mol. Biol. 2009, 16, 198–206. [Google Scholar] [CrossRef]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Janicka, A.; Martinelli, M.; Kozlowski, H.; Palumaa, P. A structural-dynamical characterization of human Cox17. J. Biol. Chem. 2008, 283, 7912–7920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, D.; Al-Ali, H.; Barrientos, A. Cmc1p is a conserved mitochondrial twin CX9C protein involved in cytochrome c oxidase biogenesis. Mol. Cell Biol. 2008, 28, 4354–4364. [Google Scholar] [CrossRef] [Green Version]
- Rae, T.D.; Schmidt, P.J.; Pufahl, R.A.; Culotta, V.C.; O’Halloran, T.V. Undetectable intracellular free copper: The requirement of a copper chaperone for superoxide dismutase. Science 1999, 284, 805–808. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Dong, L.; Outten, C.E. The redox environment in the mitochondrial intermembrane space is maintained separately from the cytosol and matrix. J. Biol. Chem. 2008, 283, 29126–29134. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Leontari, I.; Martinelli, M.; Palumaa, P.; Sillard, R.; Wang, S. Human Sco1 functional studies and pathological implications of the P174L mutant. Proc. Natl. Acad. Sci. USA 2007, 104, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Abicht, H.K.; Scharer, M.A.; Quade, N.; Ledermann, R.; Mohorko, E.; Capitani, G.; Hennecke, H.; Glockshuber, R. How periplasmic thioredoxin TlpA reduces bacterial copper chaperone ScoI and cytochrome oxidase subunit II (CoxB) prior to metallation. J. Biol. Chem. 2014, 289, 32431–32444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leary, S.C.; Sasarman, F.; Nishimura, T.; Shoubridge, E.A. Human SCO2 is required for the synthesis of CO II and as a thiol-disulphide oxidoreductase for SCO1. Hum. Mol. Genet. 2009, 18, 2230–2240. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Tokatlidis, K. The coiled coil-helix-coiled coil-helix proteins may be redox proteins. FEBS Lett. 2009, 583, 1699–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheu-Grau, D.; Wasilewski, M.; Oeljeklaus, S.; Gibhardt, C.S.; Aich, A.; Chudenkova, M.; Dennerlein, S.; Deckers, M.; Bogeski, I.; Warscheid, B.; et al. COA6 Facilitates Cytochrome c Oxidase Biogenesis as Thiol-reductase for Copper Metallochaperones in Mitochondria. J. Mol. Biol. 2020, 432, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of human cytochrome c oxidase. (a) The structure of the 14-subunit human cytochrome c oxidase is shown (PDB: 5Z62). The CuA site is present on COX2 subunit (salmon) and heme a, heme a3/CuB sites are present on COX1 subunit (green). In addition to this, the enzyme also binds other metals such as Mg and Zn. (b) The copper-containing subunits are shown. The coordination ligands of the copper centers are depicted as sticks. The Cu, Mg, and Zn metals are shown as blue, green, and grey spheres, respectively.
Figure 1.
Structure of human cytochrome c oxidase. (a) The structure of the 14-subunit human cytochrome c oxidase is shown (PDB: 5Z62). The CuA site is present on COX2 subunit (salmon) and heme a, heme a3/CuB sites are present on COX1 subunit (green). In addition to this, the enzyme also binds other metals such as Mg and Zn. (b) The copper-containing subunits are shown. The coordination ligands of the copper centers are depicted as sticks. The Cu, Mg, and Zn metals are shown as blue, green, and grey spheres, respectively.

Figure 2.
A schematic diagram of the mitochondrial copper delivery pathway to cytochrome c oxidase. The copper insertion into CcO is a modular process where COX1 and COX2 metallation occurs separately using a dedicated set of proteins. COX17 is the primary donor of copper to both COX1 and COX2, directly delivering copper to COX11 and SCO proteins. SCO1 is the metallochaperone that inserts copper into COX2, with SCO2 and COA6 facilitating the copper transfer. COX11 is the metallochaperone that inserts copper into the COX1 subunit and requires COX19 for its functionality.
Figure 2.
A schematic diagram of the mitochondrial copper delivery pathway to cytochrome c oxidase. The copper insertion into CcO is a modular process where COX1 and COX2 metallation occurs separately using a dedicated set of proteins. COX17 is the primary donor of copper to both COX1 and COX2, directly delivering copper to COX11 and SCO proteins. SCO1 is the metallochaperone that inserts copper into COX2, with SCO2 and COA6 facilitating the copper transfer. COX11 is the metallochaperone that inserts copper into the COX1 subunit and requires COX19 for its functionality.

Figure 3.
Comparison of the structure of COA6 with other human CHCH domain-containing proteins. The solution structure of COA6 (PDB 6NL3) (a) and a monomer from the crystal structure of dimeric COA6 (PDB 6PCE chain B) are shown (b). Solution structure of COX17 (PDB 2RNB) (c). Solution structure of human MIA40 (PDB 2K3J) (d). The cysteines that form the disulfide bonds in the CX9C-CX9-10C motif have been shown in sticks. The copper binding cysteines of COX17 (c) and the redox-active cysteines of MIA40 (d) have also been highlighted in sticks. Copper is shown as a blue sphere.
Figure 3.
Comparison of the structure of COA6 with other human CHCH domain-containing proteins. The solution structure of COA6 (PDB 6NL3) (a) and a monomer from the crystal structure of dimeric COA6 (PDB 6PCE chain B) are shown (b). Solution structure of COX17 (PDB 2RNB) (c). Solution structure of human MIA40 (PDB 2K3J) (d). The cysteines that form the disulfide bonds in the CX9C-CX9-10C motif have been shown in sticks. The copper binding cysteines of COX17 (c) and the redox-active cysteines of MIA40 (d) have also been highlighted in sticks. Copper is shown as a blue sphere.

Figure 4.
A schematic diagram of the biochemical role of COA6 in the metallation of COX2. The metallation of SCO1 and COX2 requires that their copper-binding cysteines are in the reduced form. COA6 acts as a disulfide reductase in this process, reducing the cysteines of both oxidized SCO1 and COX2, enabling their copper metallation by Cu-COX17 and Cu-SCO1, respectively.
Figure 4.
A schematic diagram of the biochemical role of COA6 in the metallation of COX2. The metallation of SCO1 and COX2 requires that their copper-binding cysteines are in the reduced form. COA6 acts as a disulfide reductase in this process, reducing the cysteines of both oxidized SCO1 and COX2, enabling their copper metallation by Cu-COX17 and Cu-SCO1, respectively.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Swaminathan, A.B.; Gohil, V.M. The Role of COA6 in the Mitochondrial Copper Delivery Pathway to Cytochrome c Oxidase. Biomolecules 2022, 12, 125. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12010125
AMA Style
Swaminathan AB, Gohil VM. The Role of COA6 in the Mitochondrial Copper Delivery Pathway to Cytochrome c Oxidase. Biomolecules. 2022; 12(1):125. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12010125
Chicago/Turabian StyleSwaminathan, Abhinav B., and Vishal M. Gohil. 2022. "The Role of COA6 in the Mitochondrial Copper Delivery Pathway to Cytochrome c Oxidase" Biomolecules 12, no. 1: 125. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12010125
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.