
Differential Effects of Beta-Hydroxybutyrate Enantiomers on Induced Pluripotent Stem Derived Cardiac Myocyte Electrophysiology
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. iPS-CMs Generation and Purification
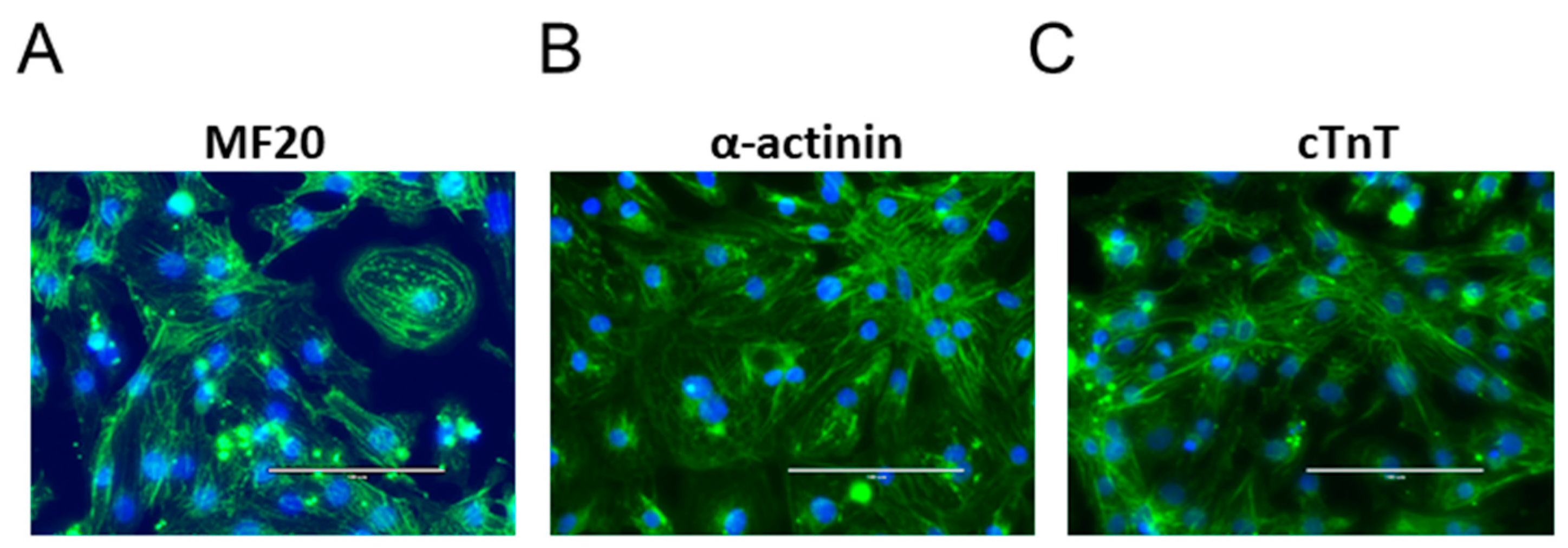
2.2. Immunofluorescence
2.3. βOHB Media
2.4. Voltage and Calcium Recordings
2.5. Data Analysis
2.6. Statistics
3. Results
3.1. βOHB Alters iPS-CMs Optical Action Potentials
3.2. βOHB Alters iPS-CMs Calcium Transients
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alberts, B. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002; p. 1464. [Google Scholar]
- Stubbs, B.J.; Cox, P.J.; Evans, R.D.; Santer, P.; Miller, J.J.; Faull, O.K.; Magor-Elliott, S.; Hiyama, S.; Stirling, M.; Clarke, K. On the Metabolism of Exogenous Ketones in Humans. Front. Physiol. 2017, 8, 848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, G.F., Jr. Fuel metabolism in starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.W.; Biton, M.; Haber, A.L.; Gunduz, N.; Eng, G.; Gaynor, L.T.; Tripathi, S.; Calibasi-Kocal, G.; Rickelt, S.; Butty, V.L.; et al. Ketone Body Signaling Mediates Intestinal Stem Cell Homeostasis and Adaptation to Diet. Cell 2019, 178, 1115–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.C.; Verdin, E. Beta-Hydroxybutyrate: A Signaling Metabolite. Annu. Rev. Nutr. 2017, 37, 51–76. [Google Scholar] [CrossRef]
- Cahill, G.F., Jr. Starvation in man. N. Engl. J. Med. 1970, 282, 668–675. [Google Scholar] [CrossRef]
- Henderson, S.T. Ketone bodies as a therapeutic for Alzheimer’s disease. Neurotherapeutics 2008, 5, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Taegtmeyer, H.; Young, M.E.; Lopaschuk, G.D.; Abel, E.D.; Brunengraber, H.; Darley-Usmar, V.; Des Rosiers, C.; Gerszten, R.; Glatz, J.F.; Griffin, J.L.; et al. Assessing Cardiac Metabolism: A Scientific Statement from the American Heart Association. Circ. Res. 2016, 118, 1659–1701. [Google Scholar] [CrossRef]
- Aubert, G.; Martin, O.J.; Horton, J.L.; Lai, L.; Vega, R.B.; Leone, T.C.; Koves, T.; Gardell, S.J.; Kruger, M.; Hoppel, C.L.; et al. The Failing Heart Relies on Ketone Bodies as a Fuel. Circulation 2016, 133, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, S.; Kelly, D.P.; Margulies, K.B. Implications of Altered Ketone Metabolism and Therapeutic Ketosis in Heart Failure. Circulation 2020, 141, 1800–1812. [Google Scholar] [CrossRef]
- Yurista, S.R.; Matsuura, T.R.; Sillje, H.H.W.; Nijholt, K.T.; McDaid, K.S.; Shewale, S.V.; Leone, T.C.; Newman, J.C.; Verdin, E.; van Veldhuisen, D.J.; et al. Ketone Ester Treatment Improves Cardiac Function and Reduces Pathologic Remodeling in Preclinical Models of Heart Failure. Circ. Heart Fail. 2021, 14, e007684. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.; Liu, J.; Li, R.; Lin, J.; Huang, Z.; Yang, Z.; Wu, X.; Huang, Z.; Zhu, Q.; Wu, X. Ketone Metabolite beta-Hydroxybutyrate Ameliorates Inflammation after Spinal Cord Injury by Inhibiting the NLRP3 Inflammasome. Neurochem. Res. 2021, 46, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Yao, A.; Li, Z.; Lyu, J.; Yu, L.; Wei, S.; Xue, L.; Wang, H.; Chen, G.Q. On the nutritional and therapeutic effects of ketone body D-beta-hydroxybutyrate. Appl. Microbiol. Biotechnol. 2021, 105, 6229–6243. [Google Scholar] [CrossRef] [PubMed]
- Puchalska, P.; Nelson, A.B.; Stagg, D.B.; Crawford, P.A. Determination of ketone bodies in biological samples via rapid UPLC-MS/MS. Talanta 2021, 225, 122048. [Google Scholar] [CrossRef]
- Neville, M.C.; Allen, J.C.; Archer, P.C.; Casey, C.E.; Seacat, J.; Keller, R.P.; Lutes, V.; Rasbach, J.; Neifert, M. Studies in human lactation: Milk volume and nutrient composition during weaning and lactogenesis. Am. J. Clin. Nutr. 1991, 54, 81–92. [Google Scholar] [CrossRef]
- Piquereau, J.; Ventura-Clapier, R. Maturation of Cardiac Energy Metabolism during Perinatal Development. Front. Physiol. 2018, 9, 959. [Google Scholar] [CrossRef] [Green Version]
- Rommel, C.; Hein, L. Four Dimensions of the Cardiac Myocyte Epigenome: From Fetal to Adult Heart. Curr. Cardiol. Rep. 2020, 22, 26. [Google Scholar] [CrossRef] [Green Version]
- Jalife, J. Cardiac Electrophysiology, 4th ed.; Saunders: Philadelphia, PA, USA, 2004. [Google Scholar]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Tohyama, S.; Hattori, F.; Sano, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Hashimoto, H.; Suzuki, T.; Yamashita, H.; Satoh, Y.; et al. Distinct metabolic flow enables large-scale purification of mouse and human pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell 2013, 12, 127–137. [Google Scholar] [CrossRef]
- Han, S.; Klos, M.; Morgenstern, S.; Ahmad, R.; Pua, I.; Suresh, S.; Hicks, K.; Devaney, E. Optical Imaging of Isolated Murine Ventricular Myocytes. J. Vis. Exp. 2020, 155, e60196. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Linders, A.; Yamak, A.; Correia, C.; Kijlstra, J.D.; Garakani, A.; Xiao, L.; Milan, D.J.; van der Meer, P.; Serra, M.; et al. Metabolic Maturation of Human Pluripotent Stem Cell-Derived Cardiomyocytes by Inhibition of HIF1alpha and LDHA. Circ. Res. 2018, 123, 1066–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klos, M.; Morgenstern, S.; Hicks, K.; Suresh, S.; Devaney, E.J. The effects of the ketone body beta-hydroxybutyrate on isolated rat ventricular myocyte excitation-contraction coupling. Arch. Biochem. Biophys. 2019, 662, 143–150. [Google Scholar] [CrossRef]
- Veech, R.L.; Chance, B.; Kashiwaya, Y.; Lardy, H.A.; Cahill, G.F., Jr. Ketone bodies, potential therapeutic uses. IUBMB Life 2001, 51, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Puchalska, P.; Crawford, P.A. Metabolic and Signaling Roles of Ketone Bodies in Health and Disease. Annu. Rev. Nutr. 2021, 41, 49–77. [Google Scholar] [CrossRef]
- Greene, A.E.; Todorova, M.T.; Seyfried, T.N. Perspectives on the metabolic management of epilepsy through dietary reduction of glucose and elevation of ketone bodies. J. Neurochem. 2003, 86, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, J. Ketone Body Supplementation—A Potential New Approach for Heart Disease. JAMA 2021, 326, 17–18. [Google Scholar] [CrossRef]
- Storoni, M.; Plant, G.T. The Therapeutic Potential of the Ketogenic Diet in Treating Progressive Multiple Sclerosis. Mult. Scler. Int. 2015, 2015, 681289. [Google Scholar] [CrossRef] [Green Version]
- Pellinen, T.; Paivarinta, E.; Isotalo, J.; Lehtovirta, M.; Itkonen, S.T.; Korkalo, L.; Erkkola, M.; Pajari, A.M. Replacing dietary animal-source proteins with plant-source proteins changes dietary intake and status of vitamins and minerals in healthy adults: A 12-week randomized controlled trial. Eur. J. Nutr. 2021, 61, 1391–1404. [Google Scholar] [CrossRef]
- Correia, C.; Koshkin, A.; Duarte, P.; Hu, D.; Teixeira, A.; Domian, I.; Serra, M.; Alves, P.M. Distinct carbon sources affect structural and functional maturation of cardiomyocytes derived from human pluripotent stem cells. Sci. Rep. 2017, 7, 8590. [Google Scholar] [CrossRef]
- Brodarac, A.; Saric, T.; Oberwallner, B.; Mahmoodzadeh, S.; Neef, K.; Albrecht, J.; Burkert, K.; Oliverio, M.; Nguemo, F.; Choi, Y.H.; et al. Susceptibility of murine induced pluripotent stem cell-derived cardiomyocytes to hypoxia and nutrient deprivation. Stem Cell Res Ther 2015, 6, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.K.; Chen, Y.C.; Kao, Y.H.; Tsai, C.F.; Yeh, Y.H.; Huang, J.L.; Cheng, C.C.; Chen, S.A.; Chen, Y.J. A monounsaturated fatty acid (oleic acid) modulates electrical activity in atrial myocytes with calcium and sodium dysregulation. Int. J. Cardiol. 2014, 176, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Lin, X.; Stachel, M.; Wang, E.; Luo, Y.; Lader, J.; Sun, X.; Delmar, M.; Bu, L. Culture in Glucose-Depleted Medium Supplemented with Fatty Acid and 3,3’,5-Triiodo-l-Thyronine Facilitates Purification and Maturation of Human Pluripotent Stem Cell-Derived Cardiomyocytes. Front. Endocrinol. 2017, 8, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahara, S.; Soni, S.; Maayah, Z.H.; Ferdaoussi, M.; Dyck, J.R.B. Ketone therapy for heart failure: Current evidence for clinical use. Cardiovasc. Res. 2022, 118, 977–987. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Company | Number |
|---|---|---|
| Anti-Sarcomeric Myosin | Developmental Studies Hybridoma Bank | MF20 |
| (R)-(−)-3-Hydroxybutyric acid sodium salt | MilliporeSigma | 298360-1G |
| (±)-Sodium 3-hydroxybutyrate | MilliporeSigma | 54965-10G-F |
| Sodium L-lactate | MilliporeSigma | L7022-5G |
| Oleic acid | MilliporeSigma | O1383-1G |
| Palmitic acid | MilliporeSigma | P0500-10G |
| D-(+)-Galactose | MilliporeSigma | G5388-100G |
| Anti-α-Actinin (Sarcomeric) | MilliporeSigma | A7732 |
| Anti-Troponin T (Cardiac Muscle) | MilliporeSigma | MABT368 |
| CHIR-99021 5 mg | Selleck Chemicals LLC | S1263 |
| Thiazovivin 5 mg | Selleck Chemicals LLC | S1459 |
| Wnt-C59 (C59) 5 mg | Selleck Chemicals LLC | S7037 |
| Essential 8 Medium | Thermo Fisher Scientific | A1517001 |
| Growth Factor Reduced Matrigel, 10 ml | Thermo Fisher Scientific | CB 40230 |
| B 27 Supplement | Thermo Fisher Scientific | 17 504 044 |
| Gibco RPMI 1640 Medium | Thermo Fisher Scientific | 11 875 093 |
| Gibco RPMI 1640 Medium, no glucose | Thermo Fisher Scientific | 11 879 020 |
| Fluo-4, AM | Thermo Fisher Scientific | F14201 |
| FluoVolt | Thermo Fisher Scientific | F10488 |
| Donkey anti-Mouse IgG Alexa Fluor 488 | Thermo Fisher Scientific | A-21202 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klos, M.L.; Hou, W.; Nsengimana, B.; Weng, S.; Yan, C.; Xu, S.; Devaney, E.; Han, S. Differential Effects of Beta-Hydroxybutyrate Enantiomers on Induced Pluripotent Stem Derived Cardiac Myocyte Electrophysiology. Biomolecules 2022, 12, 1500. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12101500
Klos ML, Hou W, Nsengimana B, Weng S, Yan C, Xu S, Devaney E, Han S. Differential Effects of Beta-Hydroxybutyrate Enantiomers on Induced Pluripotent Stem Derived Cardiac Myocyte Electrophysiology. Biomolecules. 2022; 12(10):1500. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12101500
Chicago/Turabian StyleKlos, Matthew L., Wanqing Hou, Bernard Nsengimana, Shiwang Weng, Chuyun Yan, Suowen Xu, Eric Devaney, and Shuxin Han. 2022. "Differential Effects of Beta-Hydroxybutyrate Enantiomers on Induced Pluripotent Stem Derived Cardiac Myocyte Electrophysiology" Biomolecules 12, no. 10: 1500. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12101500