
Pseudomonas aeruginosa Bacterioferritin Is Assembled from FtnA and BfrB Subunits with the Relative Proportions Dependent on the Environmental Oxygen Availability



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Bacterial Strains, and Culture Media
2.2. Cell Culture under Aerobic or O2-Limiting Conditions
2.3. Purification of Bacterioferritin (Bfr)
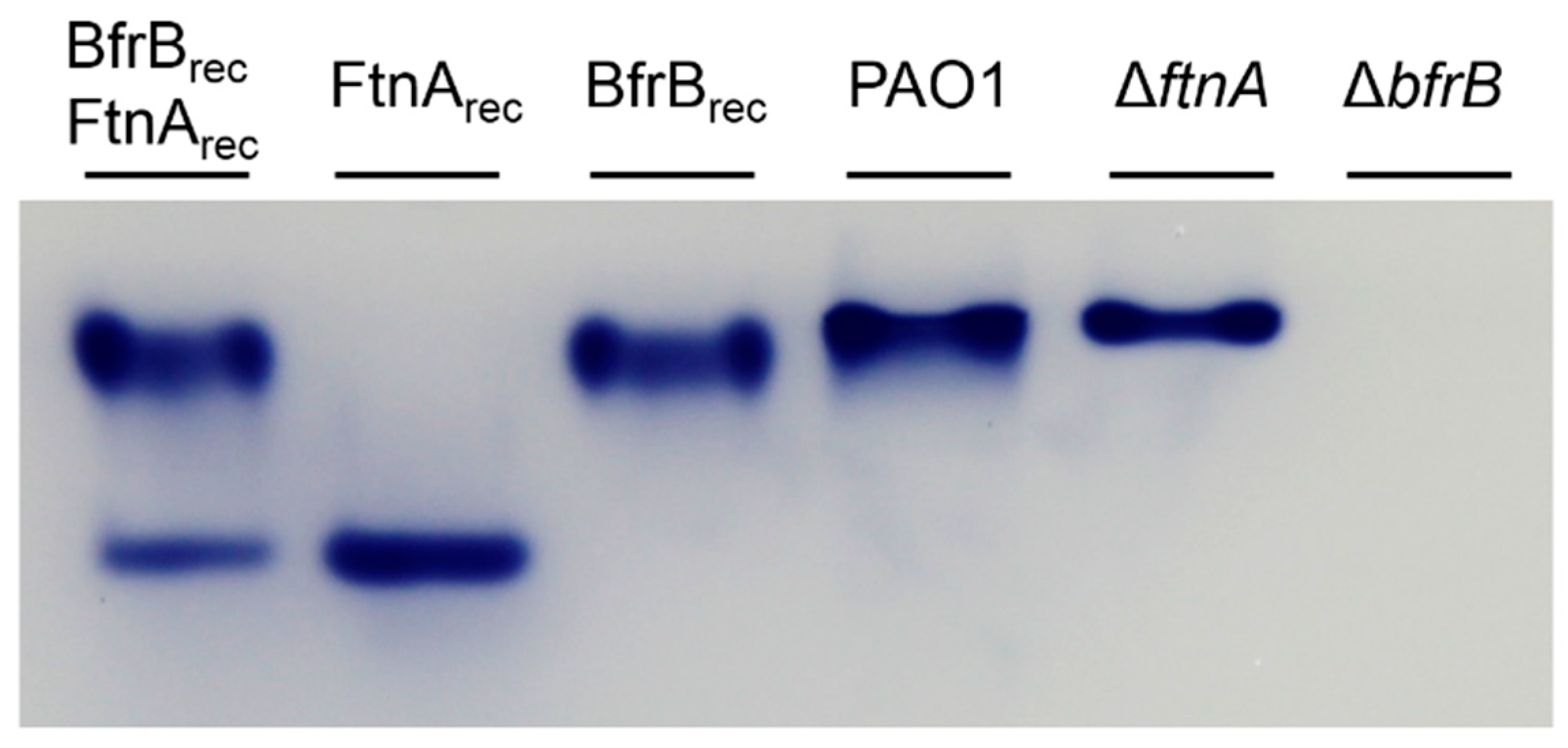
2.4. Determination of BfrB and FtnA Content in Bacterioferritin Samples Isolated from P. aeruginosa Cells
2.5. Native PAGE
2.6. In-Gel Digestion and Mass Spectrometric Characterization of Proteins
2.7. Measurement of Kd by Surface Plasmon Resonance (SPR)
3. Results and Discussion
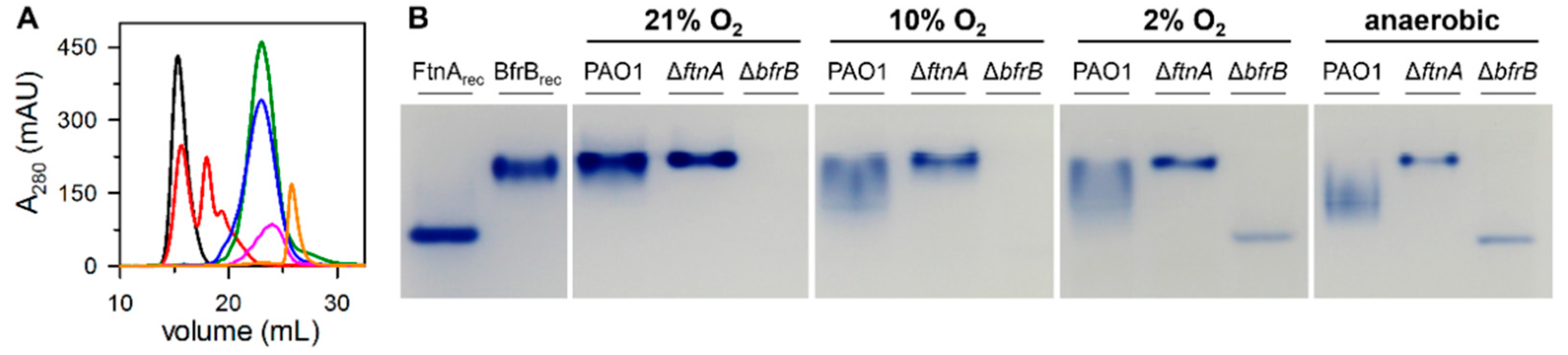
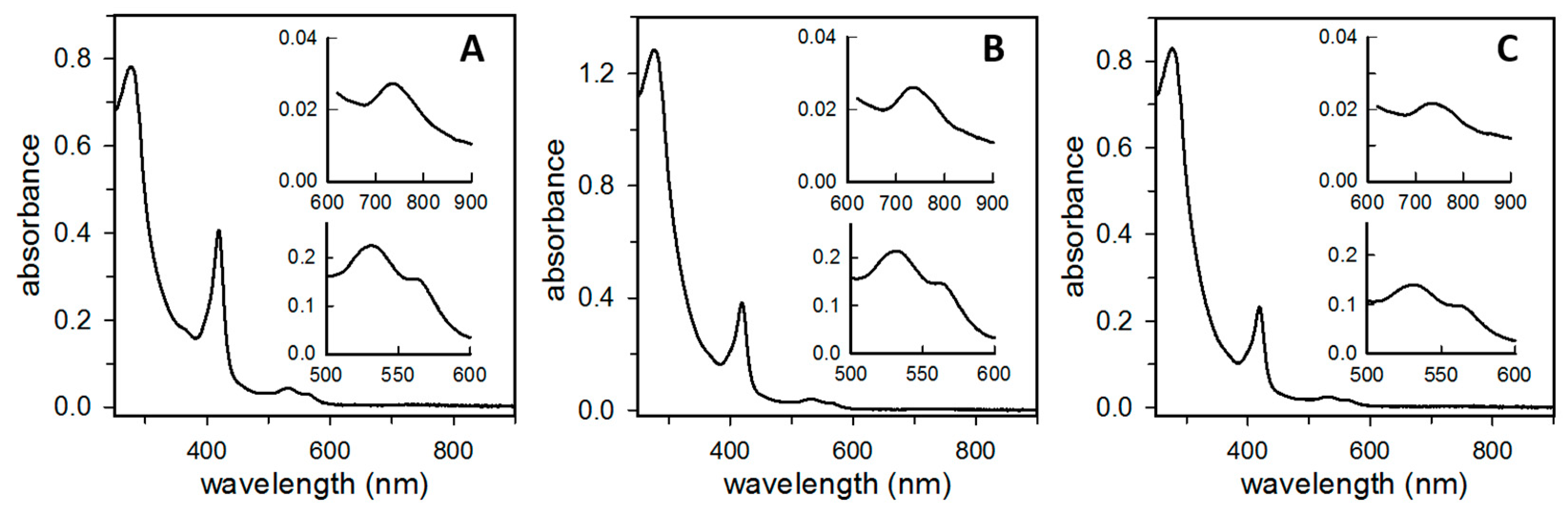
3.1. P. aeruginosa Bacterioferritin (Bfr) Is a Heterooligomeric Molecule Constituted by FtnA and BfrB Subunits
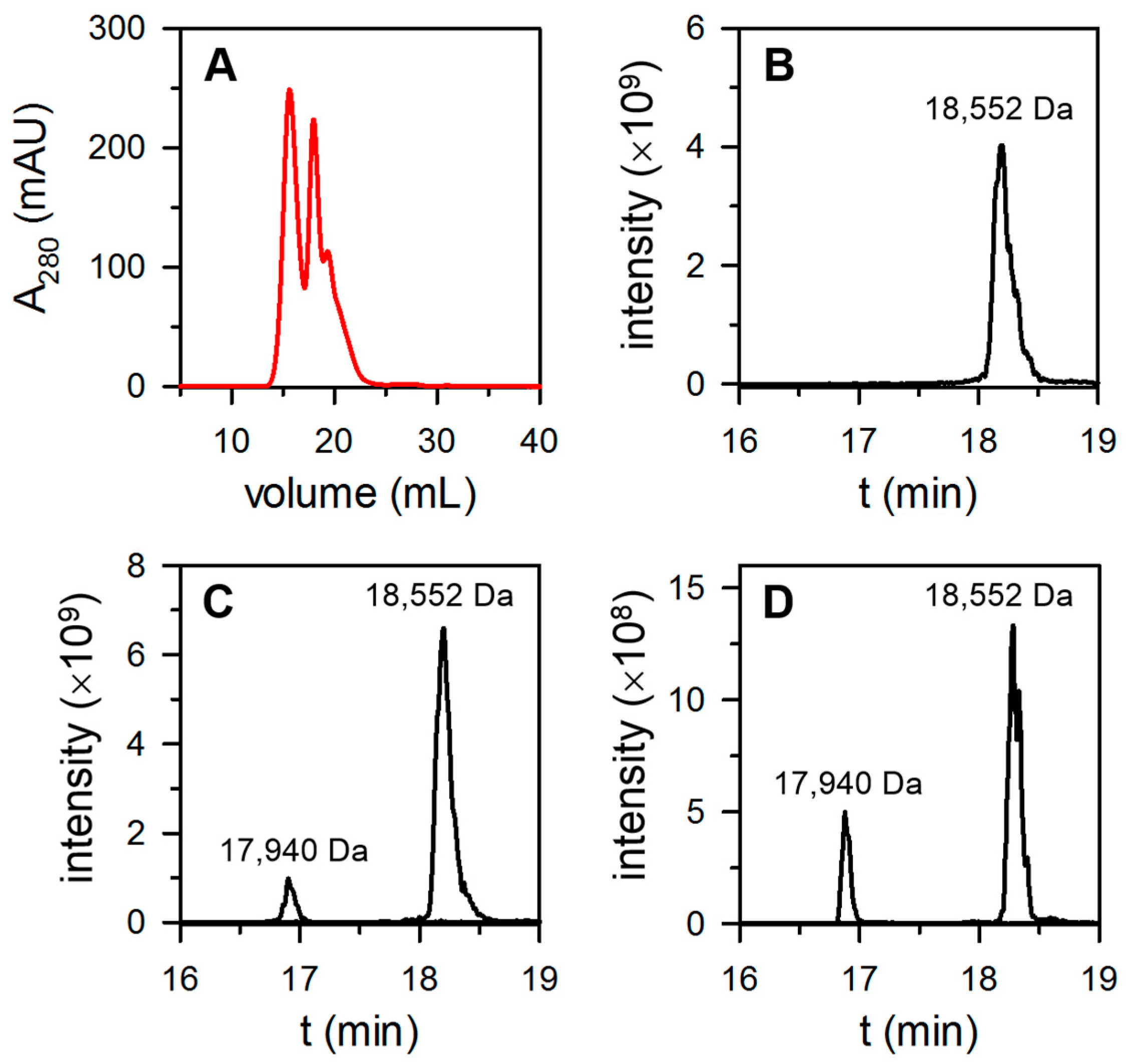
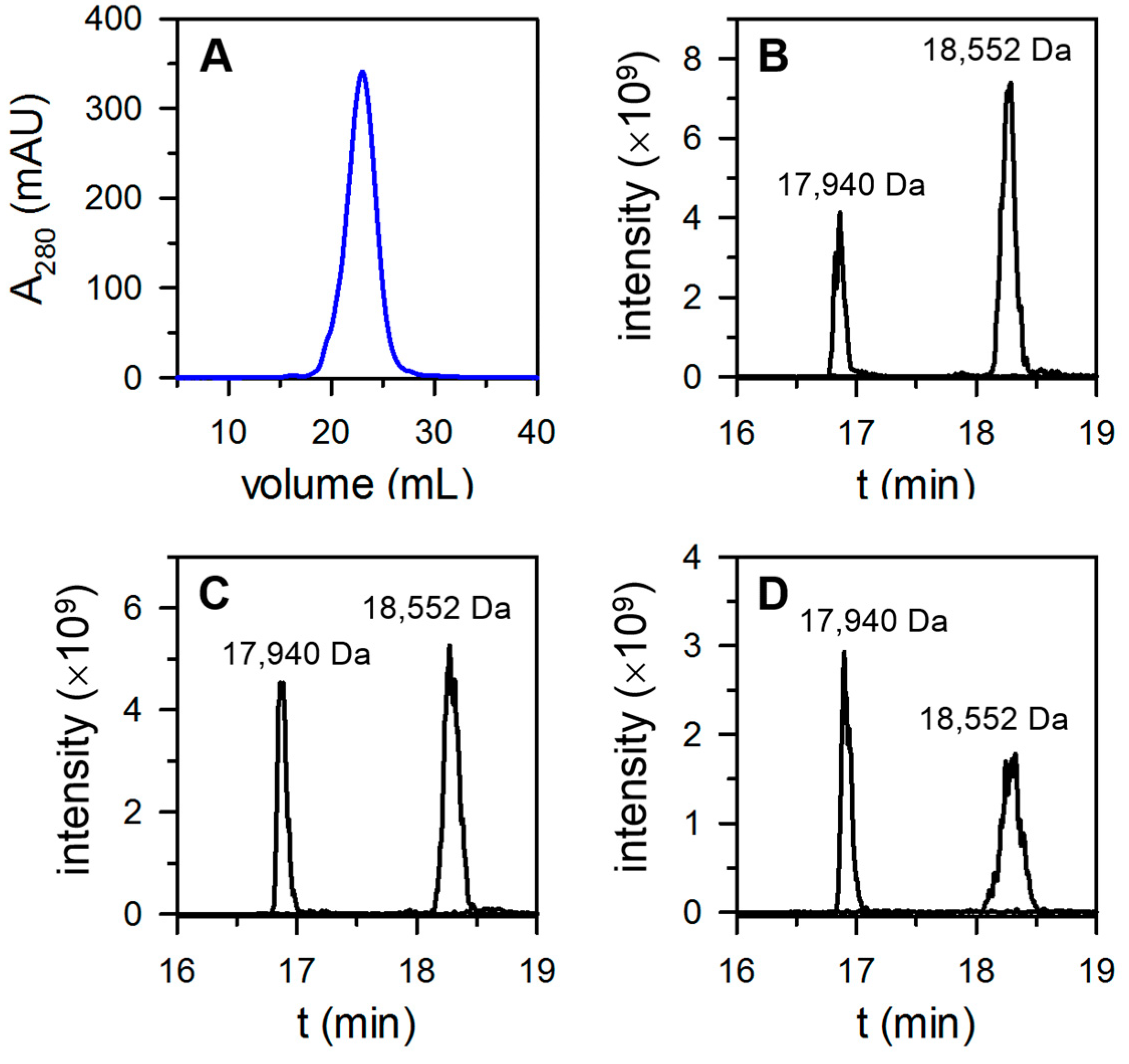
3.2. The Proportion of FtnA and BfrB Subunits in Heterooligomeric Bfr Depends on Environmental O2 Levels in the Bacterial Culture
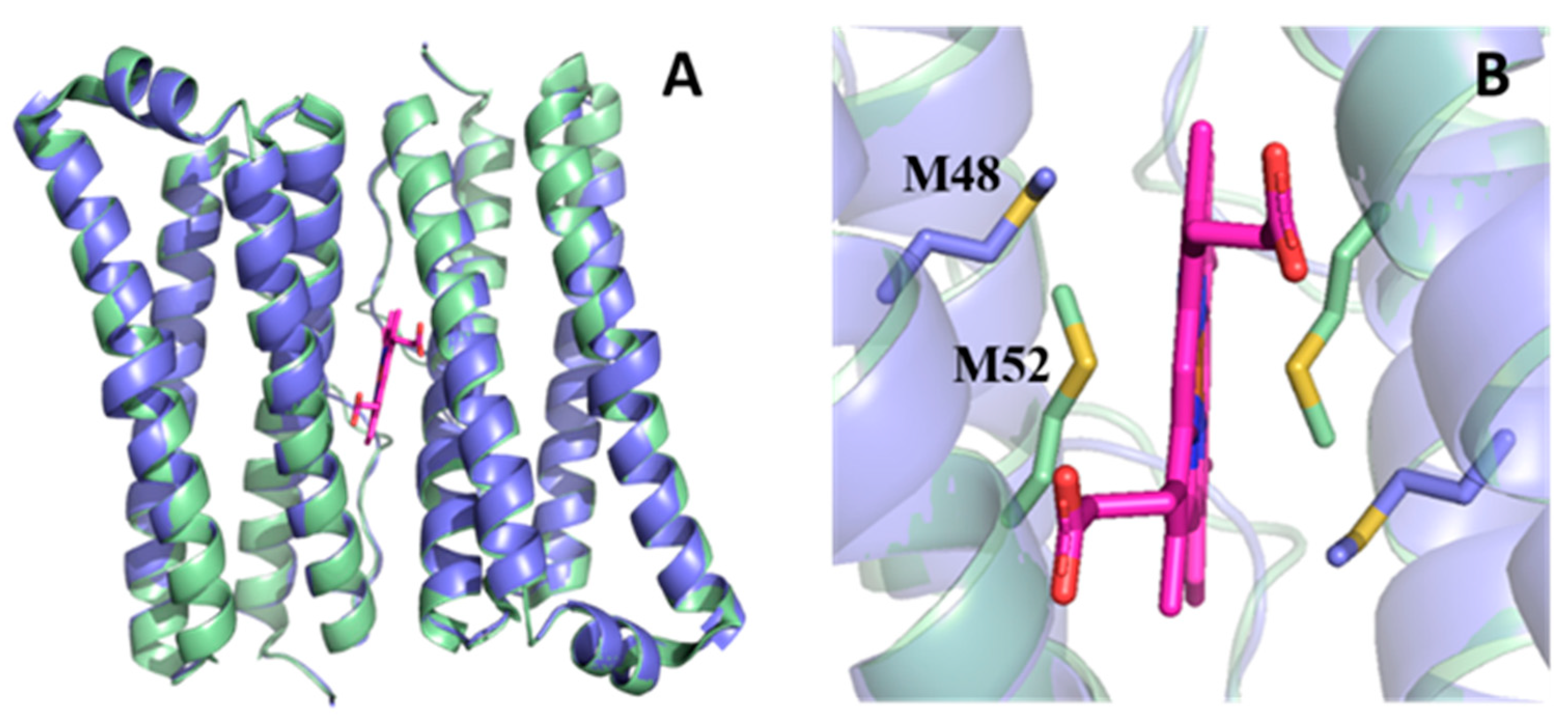
3.3. Do 24-mer FtnA Molecules Assemble in P. aeruginosa?
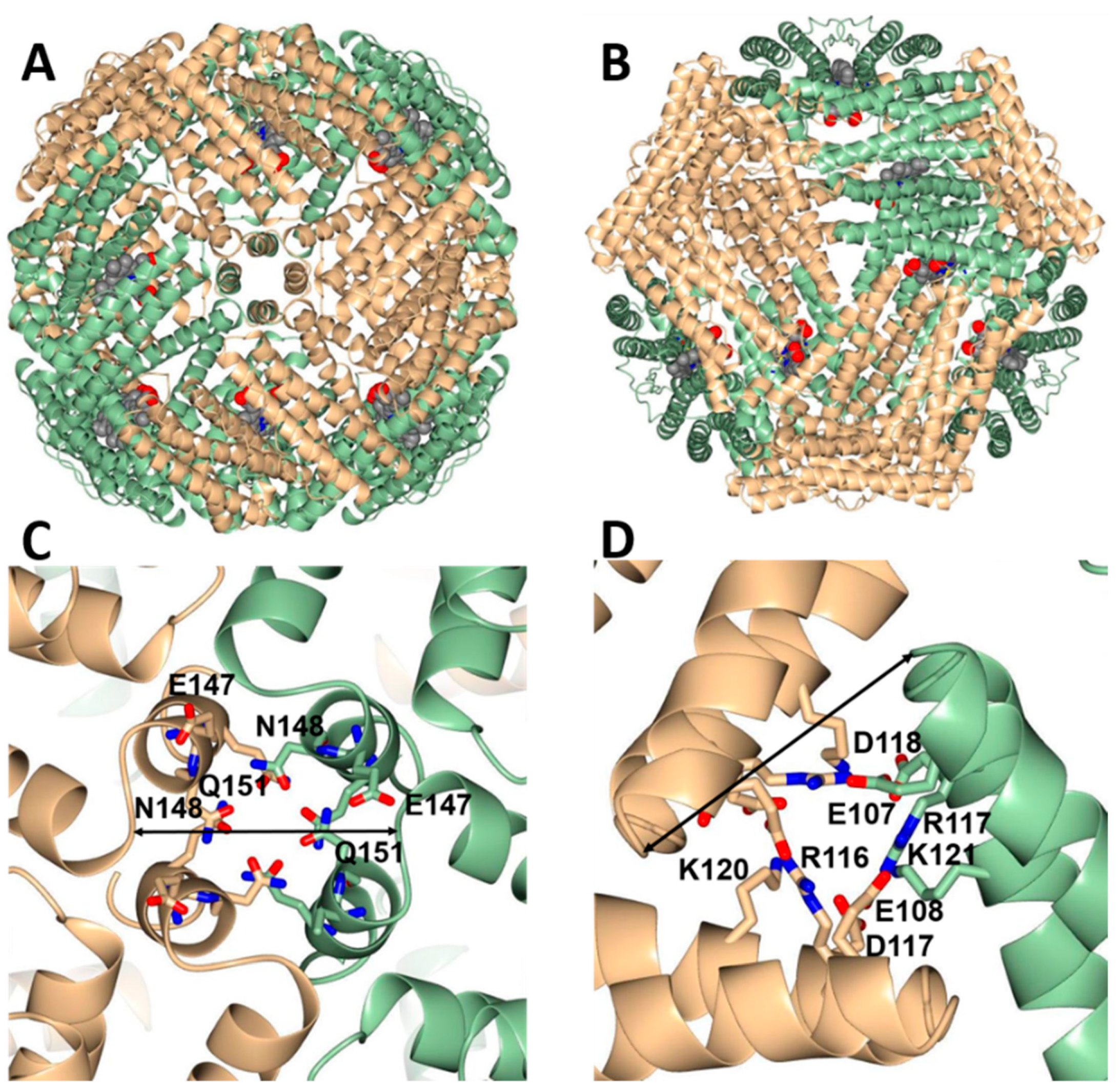
3.4. Bfr Is Assembled from FtnA and BfrB Inter-Subunit Dimers
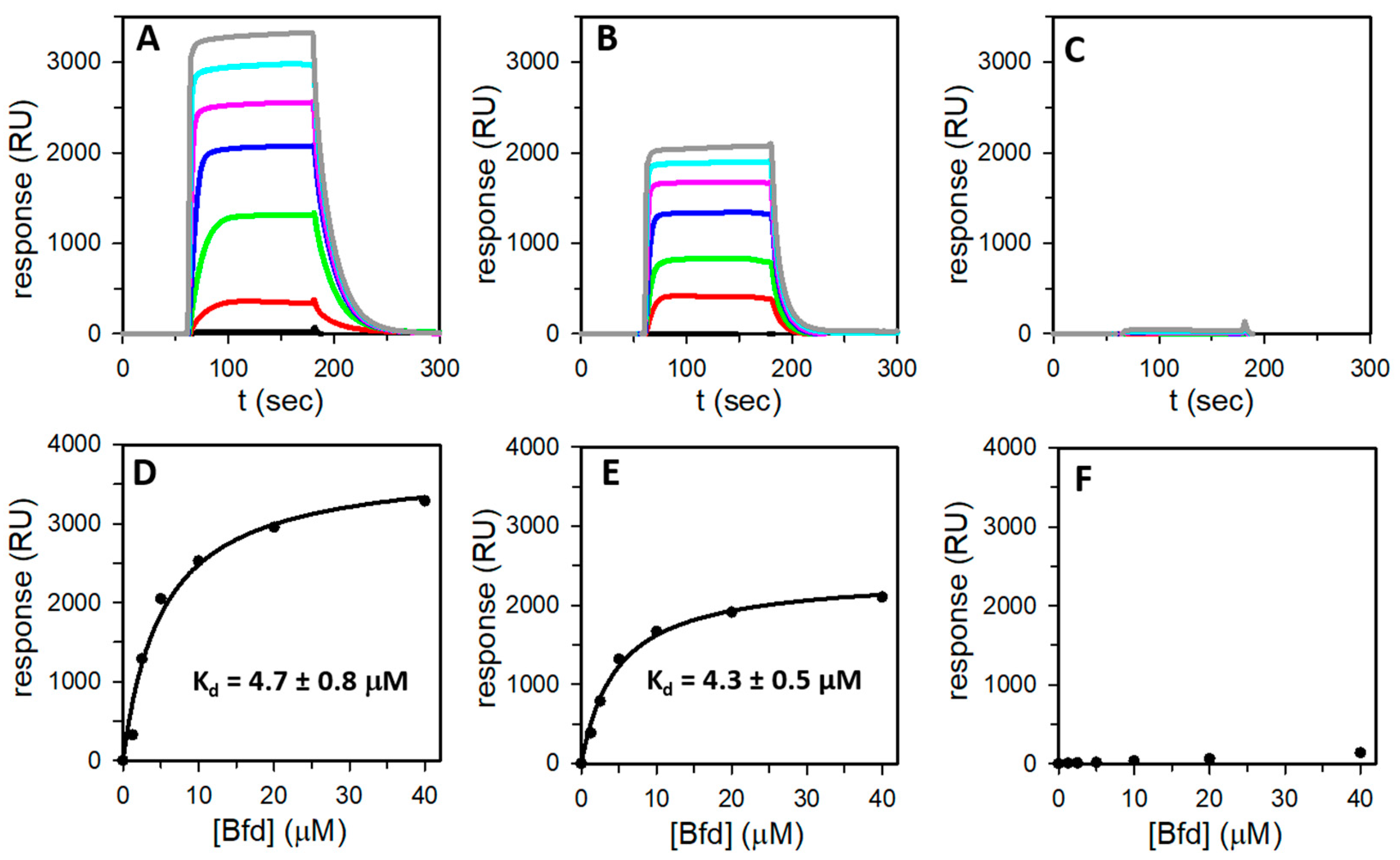
3.5. Bfd Binds Heterooligomeric Bfr at BfrB Inter-Subunit Dimers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bradley, J.M.; Svistunenko, D.A.; Wilson, M.T.; Hemmings, A.M.; Moore, G.R.; Le Brun, N.E. Bacterial iron detoxification at the molecular level. J. Biol. Chem. 2020, 295, 17602–17623. [Google Scholar] [CrossRef] [PubMed]
- Rivera, M. Bacterioferritin: Structure, Dynamics and Protein-Protein Interactions at Play in Iron Storage and Mobilization. Acc. Chem. Res. 2017, 50, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.M.; Le Brun, N.E.; Moore, G.R. Ferritins: Furnishing proteins with iron. J. Biol. Inorg. Chem. 2016, 21, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial Iron Homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Lundin, D.; Poole, A.M.; Sjoberg, B.M.; Hogbom, M. Use of structural phylogenetic networks for classification of the ferritin-like superfamily. J. Biol. Chem. 2012, 287, 20565–20575. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.C. The Ferritin-like Superfamily: Evolution of the Biological Iron Storeman from a Ruberythrin-Like Ancestor. Biochim. Biophys. Acta 2010, 1800, 691–705. [Google Scholar] [CrossRef]
- Frolow, F.; Kalb, A.J.; Yariv, J. Structure of a Unique Twofold Symmetric Haem-Binding Site. Nat. Struct. Biol. 1994, 1, 453–460. [Google Scholar] [CrossRef]
- Yao, H.; Wang, Y.; Lovell, S.; Kumar, R.; Ruvinsky, A.M.; Battaile, K.P.; Vakser, I.A.; Rivera, M. The Structure of the BfrB-Bfd Complex Reveals Protein-Protein Interactions Enabling Iron Release from Bacterioferritin. J. Am. Chem. Soc. 2012, 134, 13470–13481. [Google Scholar] [CrossRef]
- Eshelman, K.; Yao, H.; Punchi Hewage, A.N.D.; Deay, J.J.; Chandler, J.R.; Rivera, M. Inhibiting the BfrB:Bfd Interaction in Pseudomonas aeruginosa Causes Irreversible Iron Accumulation in Bacterioferritin and Iron Deficiency in the Bacterial Cell. Metallomics 2017, 9, 646–659. [Google Scholar] [CrossRef]
- Weeratunga, S.; Gee, C.E.; Lovell, S.; Zeng, Y.; Woodin, C.L.; Rivera, M. Binding of Pseudomonas aeruginosa Apobacterioferritin-Associated Ferredoxin to Bacterioferritin B Promotes Heme Mediation of Electron Delivery and Mobilization of Core Mineral Iron. Biochemistry 2009, 48, 7420–7431. [Google Scholar] [CrossRef] [Green Version]
- Pullin, J.; Bradley, J.M.; Moore, G.R.; Le Brun, N.E.; Wilson, M.T.; Svistunenko, D.A. Electron Transfer from Haem to the Di-Iron Ferroxidase Centre in Bacterioferritin. Angew. Chem. (Int. Ed. Engl.) 2021, 60, 8376–8379. [Google Scholar] [CrossRef] [PubMed]
- Ruvinsky, A.M.; Vakser, I.A.; Rivera, M. Local packing modulates diversity of iron pathways and cooperative behavior in eukaryotic and prokaryotic ferritins. J. Chem. Phys. 2014, 140, 115104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honarmand Ebrahimi, K.; Hagedoorn, P.L.; Hagen, W.R. Unity in the biochemistry of the iron-storage proteins ferritin and bacterioferritin. Chem. Rev. 2015, 115, 295–326. [Google Scholar] [CrossRef]
- Chen, C.Y.; Morse, S.A. Neisseria gonorrhoeae bacterioferritin: Structural heterogeneity, involvement in iron storage and protection against oxidative stress. Microbiology 1999, 145 Pt 10, 2967–2975. [Google Scholar] [CrossRef] [Green Version]
- Bertani, L.E.; Huang, J.S.; Weir, B.A.; Kirschvink, J.L. Evidence for two types of subunits in the bacterioferritin of Magnetospirillum magnetotacticum. Gene 1997, 201, 31–36. [Google Scholar] [CrossRef]
- Uebe, R.; Ahrens, F.; Stang, J.; Jager, K.; Bottger, L.H.; Schmidt, C.; Matzanke, B.F.; Schuler, D. Bacterioferritin of Magnetospirillum gryphiswaldense Is a Heterotetraeicosameric Complex Composed of Functionally Distinct Subunits but Is Not Involved in Magnetite Biomineralization. mBio 2019, 10, e02795-18. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.R.; Kadir, H.A.; Al-Massad, K.; Le Brun, N.E.; Thomson, A.J.; Greenwood, C.; Keen, J.N.; Findlay, J.B.C. Structural Heterogeneity of Pseudomonas aeruginosa Bacterioferritin. Biochem. J. 1994, 304, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-F.; Ochsner, U.A.; Klotz, M.G.; Nanayakkara, V.K.; Howell, M.L.; Johnson, Z.; Posey, J.E.; Vasil, M.L.; Monaco, J.J.; Hassett, D.J. Bacterioferritin A Modulates Catalase A (KatA) Activity and Resistance to Hydrogen Peroxide in Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 3730–3742. [Google Scholar] [CrossRef] [Green Version]
- Weeratunga, S.; Lovell, S.; Yao, H.; Battaile, K.P.; Fischer, C.J.; Gee, C.E.; Rivera, M. Structural Studies of Bacterioferritin B (BfrB) from Pseudomonas aeruginosa Suggest a Gating Mechanism for Iron Uptake via the Ferroxidase Center. Biochemistry 2010, 49, 1160–1175. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Jepkorir, G.; Lovell, S.; Nama, P.V.; Weeratunga, S.K.; Battaille, K.P.; Rivera, M. Two Disctinct Ferritin-Like Molecules in P. aeruginosa: The Product of the bfrA Gene is a Bacterial Ferritin (FtnA) not a bacterioferritin (Bfr). Biochemistry 2011, 50, 5236–5248. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Schweizer, H.P. mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.C. A specific iron stain for iron-binding proteins in polyacrylamide gels: Application to transferrin and lactoferrin. Anal. Biochem. 1985, 148, 498–502. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, H.; Cheng, Y.; Lovell, S.; Battaile, K.P.; Middaugh, C.R.; Rivera, M. Characterization of the Bacterioferritin/Bacterioferritin Associated Ferredoxin Potein-Protein Interactions in Solution and Determination of Binding Energy Hot Spots. Biochemistry 2015, 54, 6162–6175. [Google Scholar] [CrossRef] [PubMed]
- Wijerathne, H.; Yao, H.; Wang, Y.; Lovell, S.; Battaile, K.P.; Rivera, M. Bfd, a New Class of [2Fe-2S] Protein That Functions in Bacterial Iron Homeostasis, Requires a Structural Anion Binding Site. Biochemistry 2018, 57, 5533–5543. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Wilderman, P.J.; Vasil, A.I.; Vasil, M.L. GeneChip Expression Analysis of the Iron Starvation Response in Pseudomonas Aeruginosa: Identification of Novel Pyoverdine Biosynthesis Genes. Mol. Microbiol. 2002, 45, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Soldano, A.; Yao, H.; Chandler, J.R.; Rivera, M. Inhibiting Iron Mobilization from Bacterioferritin in Pseudomonas aeruginosa Impairs Biofilm Formation Irrespective of Environmental Iron Availability. ACS Infect. Dis. 2020, 6, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Soldano, A.; Yao, H.; Punchi Hewage, A.N.D.; Meraz, K.; Annor-Gyamfi, J.K.; Bunce, R.A.; Battaile, K.P.; Lovell, S.; Rivera, M. Small Molecule Inhibitors of the Bacterioferritin (BfrB)-Ferredoxin (Bfd) Complex Kill Biofilm-Embedded Pseudomonas aeruginosa Cells. ACS Infect. Dis. 2021, 7, 123–140. [Google Scholar] [CrossRef]
- Hammond, J.H.; Dolben, E.F.; Smith, T.J.; Bhuju, S.; Hogan, D.A. Links between Anr and Quorum Sensing in Pseudomonas aeruginosa Biofilms. J. Bacteriol. 2015, 197, 2810–2820. [Google Scholar] [CrossRef] [Green Version]
- Trunk, K.; Benkert, B.; Quack, N.; Munch, R.; Scheer, M.; Garbe, J.; Jansch, L.; Trost, M.; Wehland, J.; Buer, J.; et al. Anaerobic adaptation in Pseudomonas aeruginosa: Definition of the Anr and Dnr regulons. Environ. Microbiol. 2010, 12, 1719–1733. [Google Scholar] [CrossRef]
- Clay, M.E.; Hammond, J.H.; Zhong, F.; Chen, X.; Kowalski, C.H.; Lee, A.J.; Porter, M.S.; Hampton, T.H.; Greene, C.S.; Pletneva, E.V.; et al. Pseudomonas aeruginosa lasR mutant fitness in microoxia is supported by an Anr-regulated oxygen-binding hemerythrin. Proc. Natl. Acad. Sci. USA 2020, 117, 3167–3173. [Google Scholar] [CrossRef]
- Sabra, W.; Kim, E.J.; Zeng, A.P. Physiological responses of Pseudomonas aeruginosa PAO1 to oxidative stress in controlled microaerobic and aerobic cultures. Microbiology 2002, 148, 3195–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yariv, J.; Kalb, A.J.; Sperling, R.; Bauminger, E.R.; Cohen, S.G.; Ofer, S. The Composition and Structure of Bacterioferritin of Escherichia coli. Biochem. J. 1981, 197, 171–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringeling, P.L.; Davy, S.L.; Monkara, F.A.; Hunt, C.; Dickson, D.P.; McEwan, A.G.; Moore, G.R. Iron metabolism in Rhodobacter capsulatus. Characterisation of bacterioferritin and formation of non-haem iron particles in intact cells. Eur. J. Biochem. 1994, 223, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, A.; Parida, A.; Subhadarshanee, B.; Behera, N.; Subudhi, T.; Koochana, P.K.; Behera, R.K. Alteration of Coaxial Heme Ligands Reveals the Role of Heme in Bacterioferritin from Mycobacterium tuberculosis. Inorg. Chem. 2021, 60, 16937–16952. [Google Scholar] [CrossRef]
- Punchi Hewage, A.N.D.; Fontenot, L.; Guidry, J.; Weldeghiorghis, T.; Mehta, A.K.; Donnarumma, F.; Rivera, M. Mobilization of Iron Stored in Bacterioferritin Is Required for Metabolic Homeostasis in Pseudomonas aeruginosa. Pathogens 2020, 9, 980. [Google Scholar] [CrossRef]
- Punchi Hewage, A.N.D.; Yao, H.; Nammalwar, B.; Gnanasekaran, K.K.; Lovell, S.; Bunce, R.A.; Eshelman, K.; Phaniraj, S.M.; Lee, M.M.; Peterson, B.R.; et al. Small Molecule Inhibitors of the BfrB-Bfd Interaction Decrease Pseudomonas aeruginosa Fitness and Potentiate Fluoroquinolone Activity. J. Am. Chem. Soc. 2019, 141, 8171–8184. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Ortega, C.; Harwood, C.S. Responses of Pseudomonas aeruginosa to low oxygen indicate that growth in the cystic fibrosis lung is by aerobic respiration. Mol. Microbiol. 2007, 65, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Pullin, J.; Wilson, M.T.; Clemancey, M.; Blondin, G.; Bradley, J.M.; Moore, G.R.; Le Brun, N.E.; Lucic, M.; Worrall, J.A.R.; Svistunenko, D.A. Iron Oxidation in Escherichia coli Bacterioferritin Ferroxidase Centre, a Site Designed to React Rapidly with H2O2 but Slowly with O2. Angew. Chem. (Int. Ed. Engl.) 2021, 60, 8361–8369. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction from SourceTM 15Q | %FtnA in Bfr | |||
|---|---|---|---|---|
| 21% O2 | 10% O2 | 2% O2 | Anaerobic | |
| 1st peak a | 0 | |||
| 2nd peak a | 10 ± 1 | |||
| shoulder a | 21 ± 0.6 | |||
| left b | 28 ± 1 | 30 ± 4 | 29 ± 1 | |
| center b | 38 ± 2 | 45 ± 6 | 47 ± 2 | |
| right b | 49 ± 3 | 59 ± 1 | 69 ± 2 | |
| %FtnA in Bfr | Kd (μM) | Immobilization Level (RU) | Rmax | Ref. |
|---|---|---|---|---|
| 0 (21% O2) | 4.7 ± 0.8 | 14,003 | 3430 | This work |
| 45 (2% O2) | 4.3 ± 0.5 | 14,059 | 2169 | This work |
| 100 (FtnArec) | - | 9497 | - | This work |
| 0 (BfrBrec) | 3.3 ± 0.5 | - | - | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, H.; Soldano, A.; Fontenot, L.; Donnarumma, F.; Lovell, S.; Chandler, J.R.; Rivera, M. Pseudomonas aeruginosa Bacterioferritin Is Assembled from FtnA and BfrB Subunits with the Relative Proportions Dependent on the Environmental Oxygen Availability. Biomolecules 2022, 12, 366. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12030366
Yao H, Soldano A, Fontenot L, Donnarumma F, Lovell S, Chandler JR, Rivera M. Pseudomonas aeruginosa Bacterioferritin Is Assembled from FtnA and BfrB Subunits with the Relative Proportions Dependent on the Environmental Oxygen Availability. Biomolecules. 2022; 12(3):366. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12030366
Chicago/Turabian StyleYao, Huili, Anabel Soldano, Leo Fontenot, Fabrizio Donnarumma, Scott Lovell, Josephine R. Chandler, and Mario Rivera. 2022. "Pseudomonas aeruginosa Bacterioferritin Is Assembled from FtnA and BfrB Subunits with the Relative Proportions Dependent on the Environmental Oxygen Availability" Biomolecules 12, no. 3: 366. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12030366