
Preferential Expression of Ca2+-Stimulable Adenylyl Cyclase III in the Supraventricular Area, including Arrhythmogenic Pulmonary Vein of the Rat Heart

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Preparation
2.2. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.3. Immunohistochemistry
2.4. Cell Isolation
2.5. Immunocytochemistry
2.6. Patch-Clamp Electrophysiology
2.7. Drugs
2.8. Statistics
3. Results
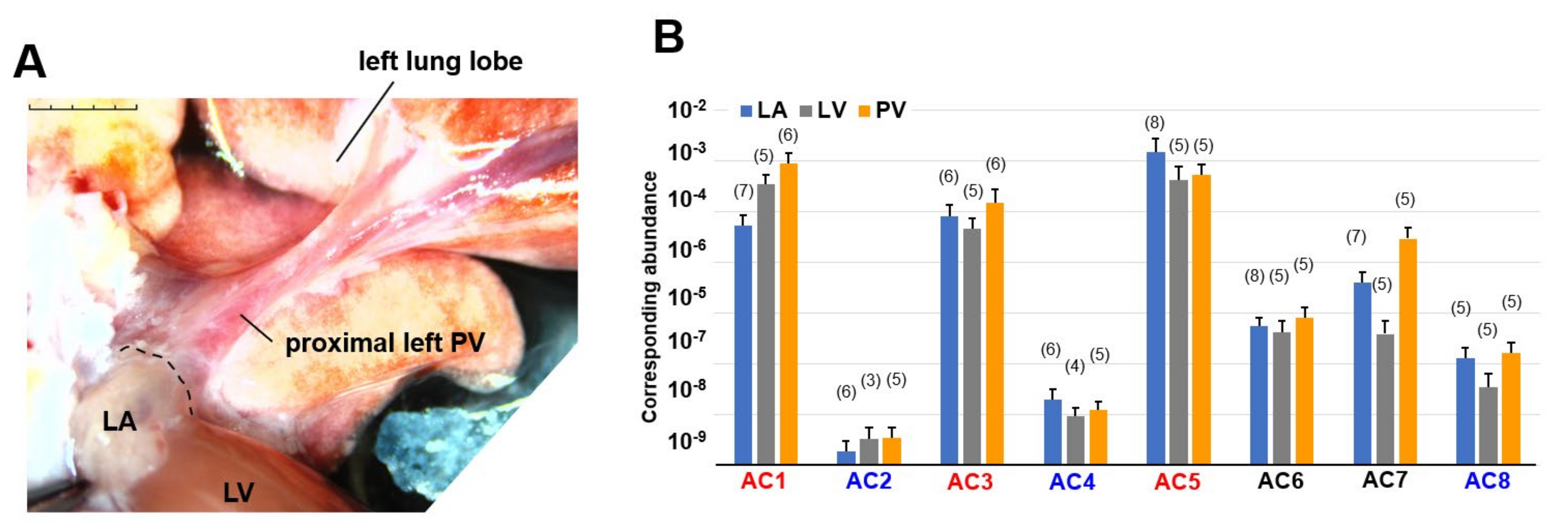
3.1. Genetic Screening of AC Family Members in Cardiopulmonary Organs by RT-PCR
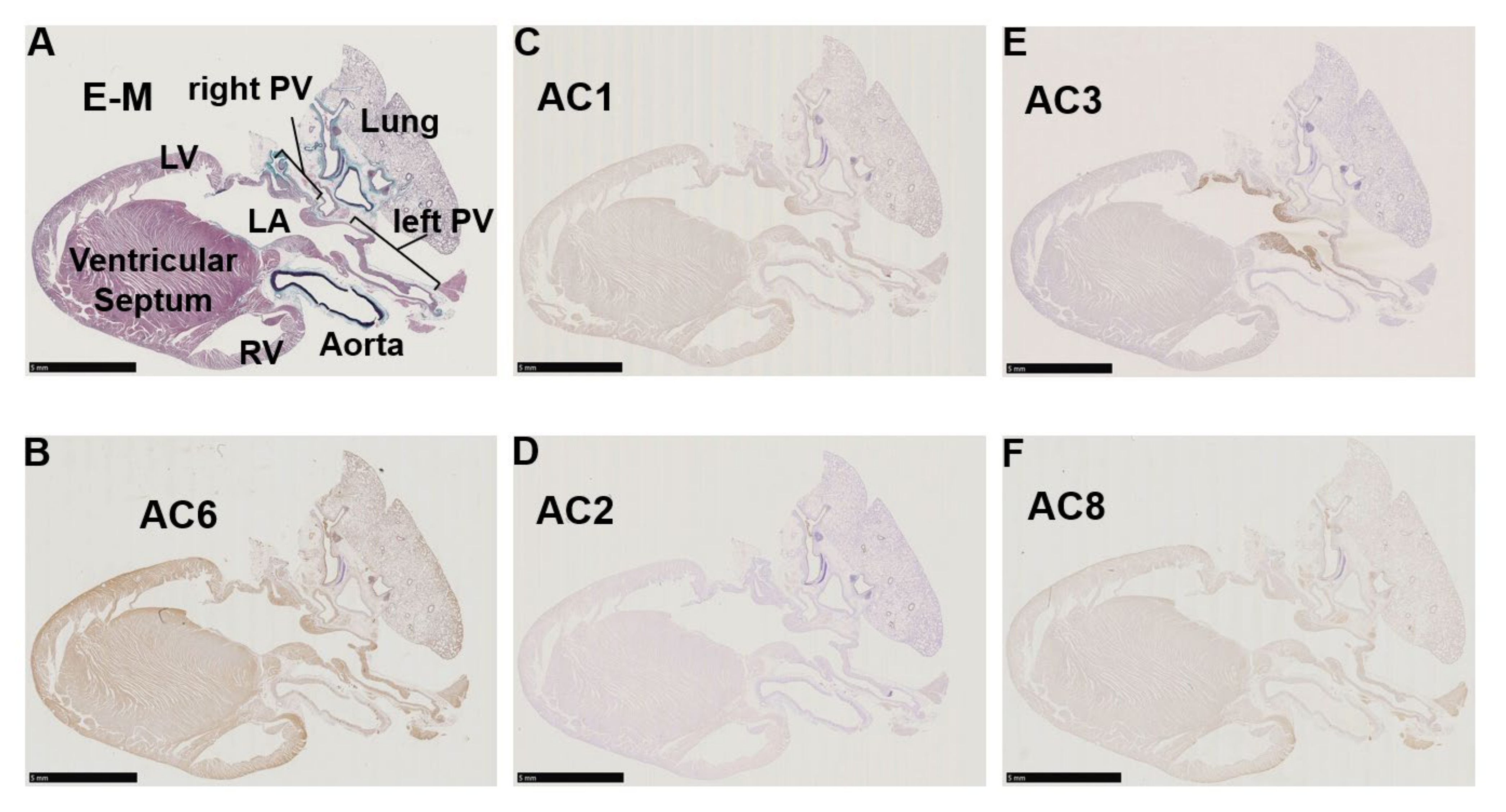
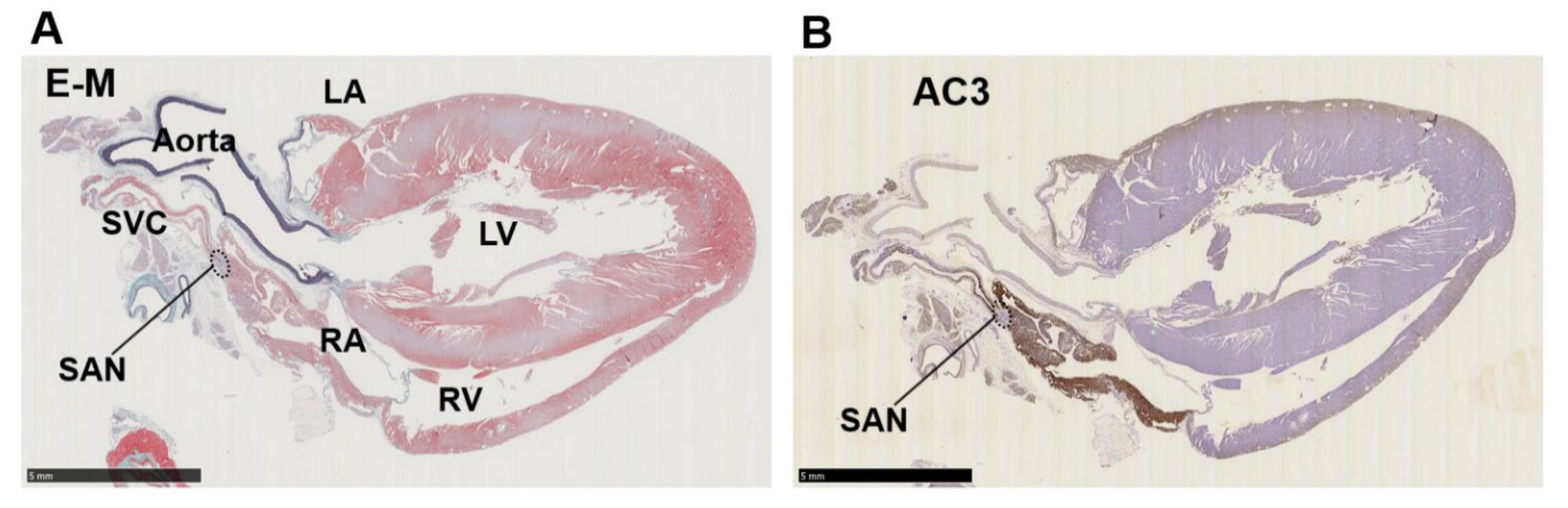
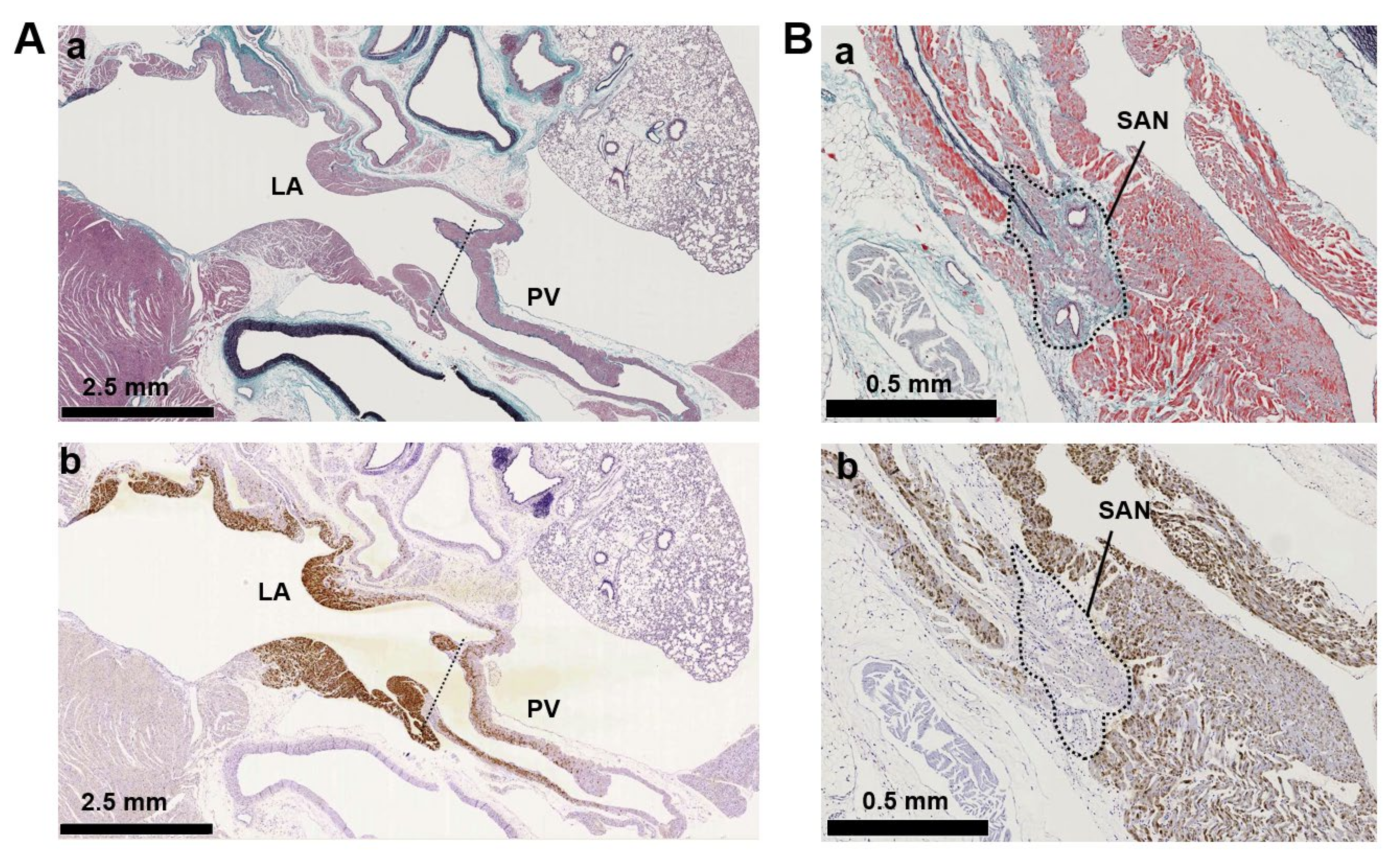
3.2. Immunohistochemistry of AC Family Members in Cardiopulmonary Regions
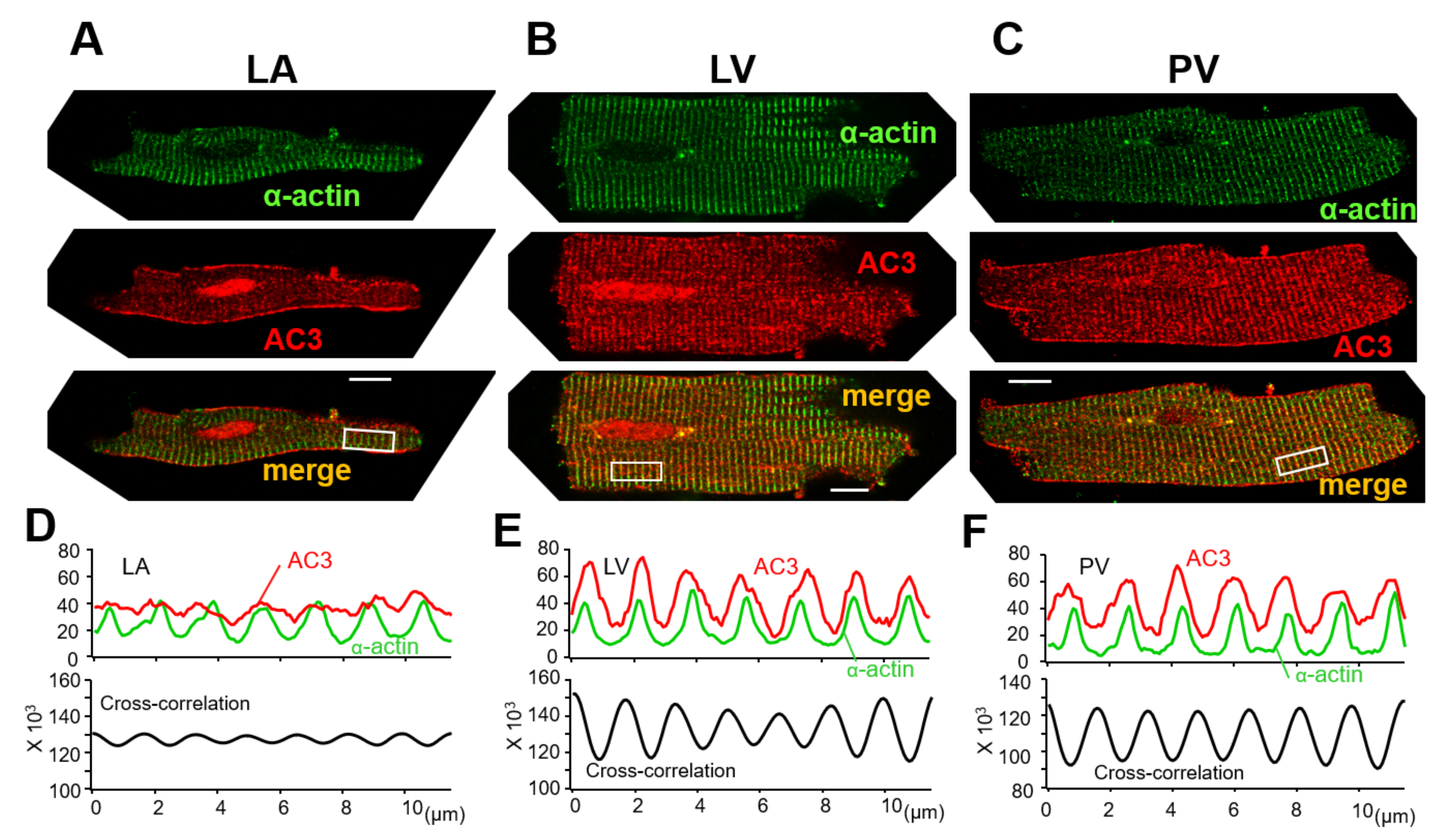
3.3. AC3 Is Localized in the t-Tubule Microdomains of PV Cardiomyocytes
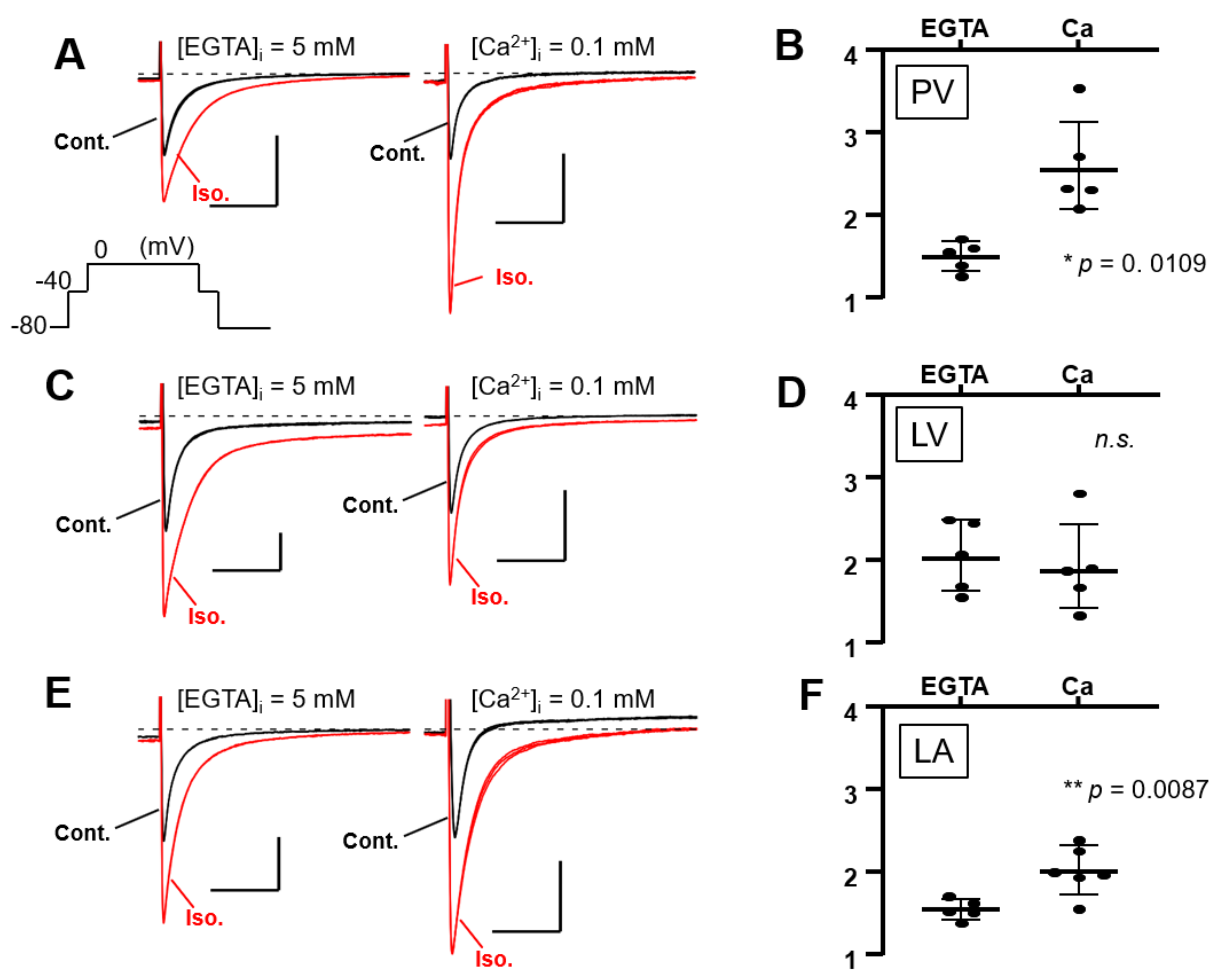
3.4. Full Activation of L-Type Ca2+ Current by β-Adrenergic Receptor Agonist Requires Intracellular Ca2+ in the Supraventricular Area, Unlike in the Ventricles
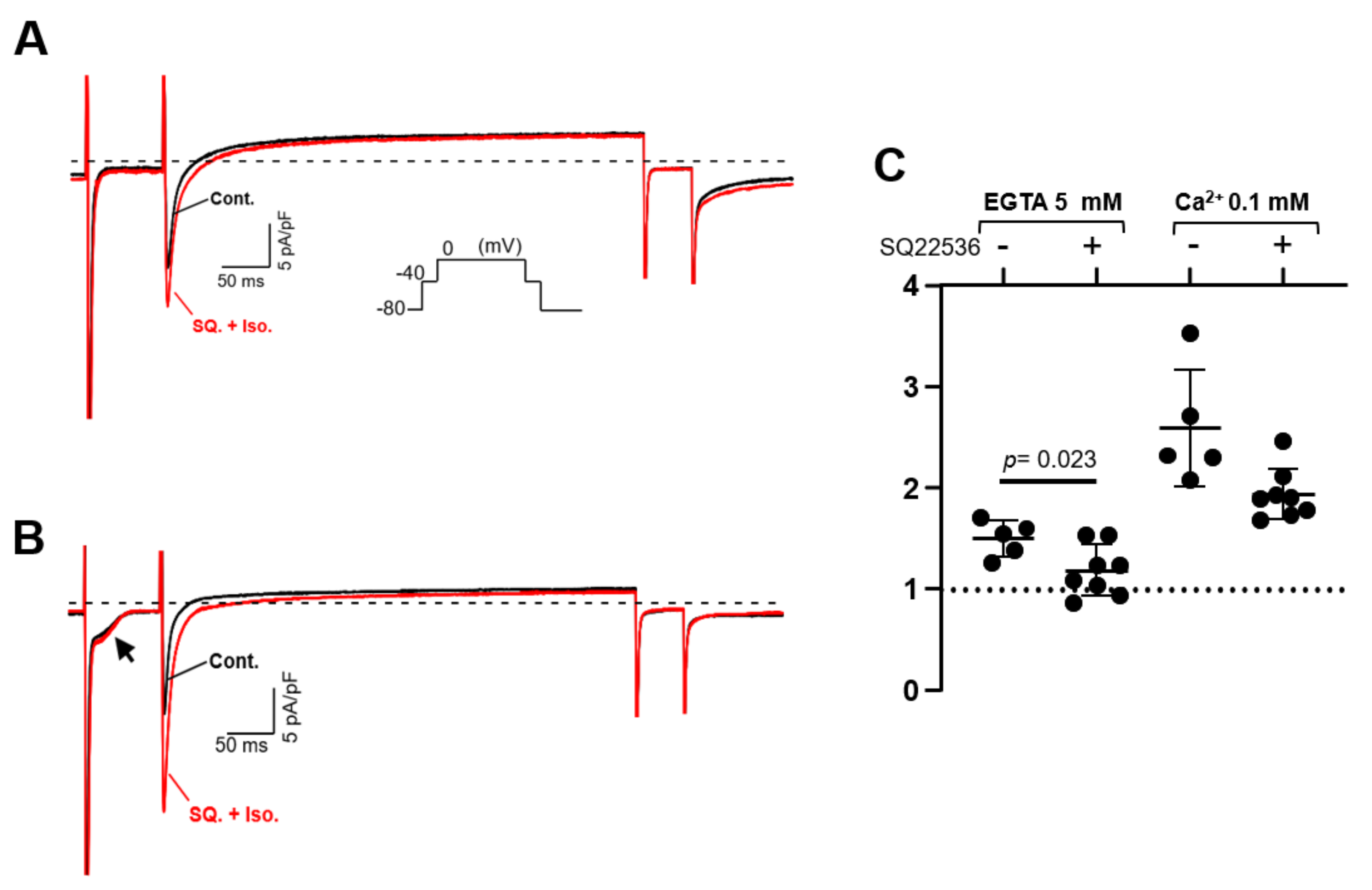
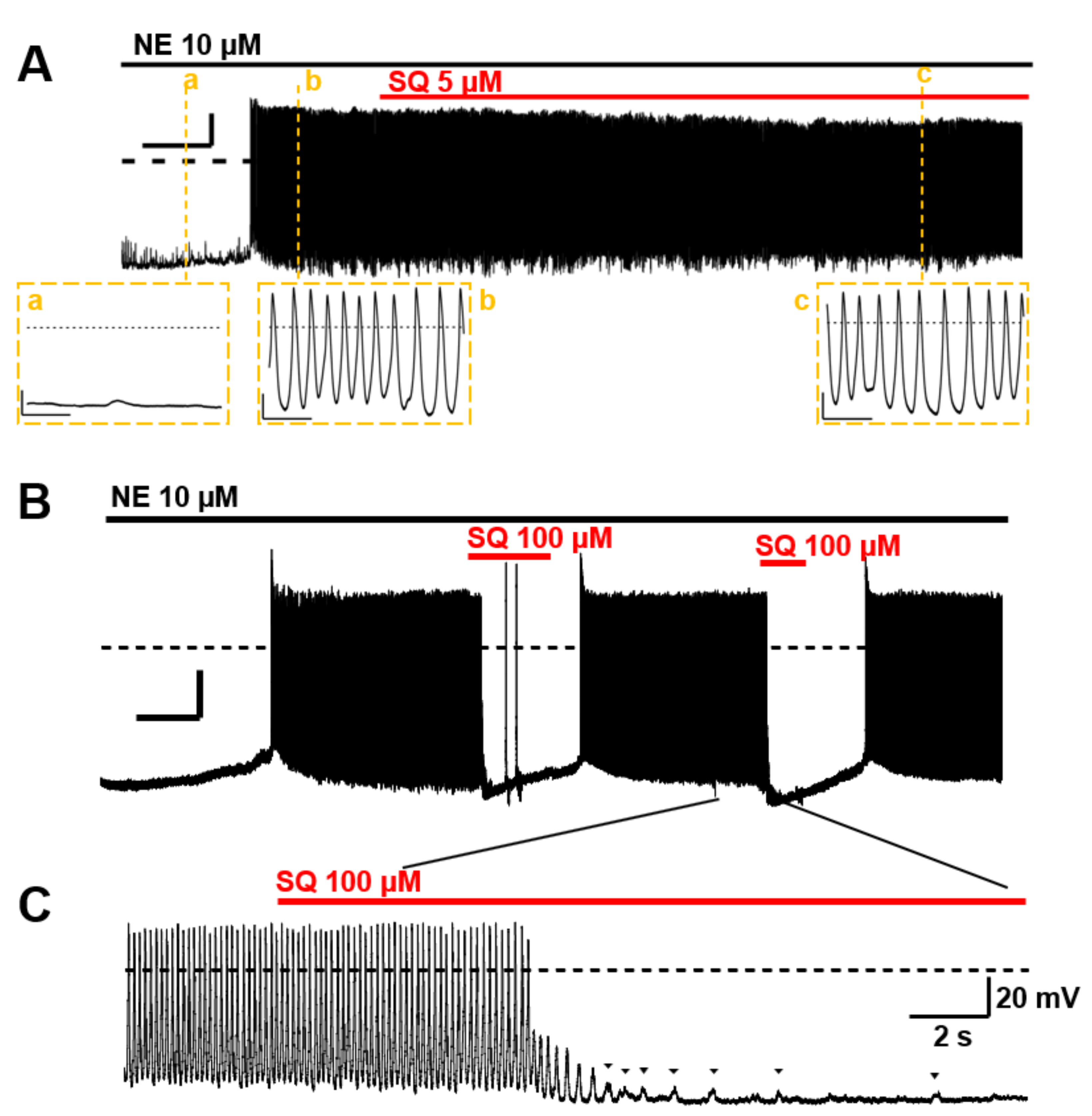
3.5. Suppression of Norepinephrine-Induced PV Automaticity by AC Inhibitor Treatment
4. Discussion
4.1. Major Findings
4.2. Potential Role of AC3 in the Rat Heart, among Other ACs
4.3. Interpretation of Region-Dependent AC3 Expression
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.H.; McAnulty, J.H.; Zheng, Z.J.; et al. Worldwide epidemiology of atrial fibrillation: A global burden of disease 2010 study. Circulation 2014, 129, 837–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, D.W. Electrical activity of the pulmonary vein and its interaction with the right atrium in the guinea-pig. J. Physiol. 1981, 314, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doisne, N.; Maupoil, V.; Cosnay, P.; Findlay, I. Catecholaminergic automatic activity in the rat pulmonary vein: Electrophysiological differences between cardiac muscle in the left atrium and pulmonary vein. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H102–H108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namekata, I.; Tsuneoka, Y.; Takahara, A.; Shimada, H.; Sugimoto, T. Short Communication Involvement of the Na+/Ca 2+ Exchanger in the Automaticity of Guinea-Pig Pulmonary Vein Myocardium as Revealed by SEA0400. J. Pharmacol. Sci. 2009, 116, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, Y.; Takano, M.; Ohba, T.; Ono, K. Arrhythmogenic coupling between the Na+–Ca2+ exchanger and inositol 1,4,5-triphosphate receptor in rat pulmonary vein cardiomyocytes. J. Mol. Cell. Cardiol. 2012, 52, 988–997. [Google Scholar] [CrossRef]
- Bredeloux, P.; Pasqualin, C.; Bordy, R.; Maupoil, V.; Findlay, I. Automatic activity arising in cardiac muscle sleeves of the pulmonary vein. Biomolecules 2022, 12, 23. [Google Scholar] [CrossRef]
- Cheung, D.W. Pulmonary vein as an ectopic focus in digitalis-induced arrhythmia. Nature 1981, 294, 582–584. [Google Scholar] [CrossRef]
- Honjo, H.; Boyett, M.R.; Niwa, R.; Inada, S.; Yamamoto, M.; Mitsui, K.; Horiuchi, T.; Shibata, N.; Kamiya, K.; Kodama, I. Pacing-induced spontaneous activity in myocardial sleeves of pulmonary veins after treatment with ryanodine. Circulation 2003, 107, 1937–1943. [Google Scholar] [CrossRef]
- Sood, S.; Chelu, M.G.; Van Oort, R.J.; Skapura, D.G.; Santonastasi, M.; Dobrev, D.; Wehrens, X.H.T. Intracellular calcium leak due to FKBP12.6 deficiency in mice facilitates the inducibility of atrial fibrillation. Heart Rhythm 2009, 1, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Landstrom, A.P.; Dobrev, D.; Wehrens, X.H.T. Calcium Signaling and Cardiac Arrhythmias. Circ. Res. 2017, 120, 1969–1993. [Google Scholar] [CrossRef]
- Maupoil, V.; Bronquard, C.; Freslon, J.-L.; Cosnay, P.; Findlay, I. Ectopic activity in the rat pulmonary vein can arise from simultaneous activation of alpha1- and beta1-adrenoceptors. Br. J. Pharmacol. 2007, 150, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Pasqualin, C.; Yu, A.; Malécot, C.O.; Gannier, F.; Cognard, C.; Godin-Ribuot, D.; Morand, J.; Bredeloux, P.; Maupoil, V. Structural heterogeneity of the rat pulmonary vein myocardium: Consequences on intracellular calcium dynamics and arrhythmogenic potential. Sci. Rep. 2018, 8, 3244. [Google Scholar] [CrossRef]
- Ehrlich, J.R.; Cha, T.-J.; Zhang, L.; Chartier, D.; Villeneuve, L.; Hébert, T.E.; Nattel, S. Characterization of a hyperpolarization-activated time-dependent potassium current in canine cardiomyocytes from pulmonary vein myocardial sleeves and left atrium. J. Physiol. 2004, 557, 583–597. [Google Scholar] [CrossRef]
- Okamoto, Y.; Kawamura, K.; Nakamura, Y.; Ono, K. Pathological impact of hyperpolarization-activated chloride current peculiar to rat pulmonary vein cardiomyocytes. J. Mol. Cell. Cardiol. 2014, 66, 53–62. [Google Scholar] [CrossRef]
- Chen, P.S.; Chen, L.S.; Fishbein, M.C.; Lin, S.F.; Nattel, S. Role of the autonomic nervous system in atrial fibrillation: Pathophysiology and therapy. Circ. Res. 2014, 114, 1500–1515. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.Y.; Chen, P.-S.; Chen, L.S.; Fishbein, M.C. Autonomic nerves in pulmonary veins. Heart Rythm. 2007, 4, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, T.; Maeda, A.; Yamazawa, T.; Hirose, K.; Kurosaki, T.; Iino, M. Encoding of Ca2+ signals by differential expression of IP3 receptor subtypes. EMBO J. 1999, 18, 1303–1308. [Google Scholar] [CrossRef]
- Okubo, Y.; Kanemaru, K.; Suzuki, J.; Kobayashi, K.; Hirose, K.; Iino, M. IP 3 R2-independent Ca 2+ release from the endoplasmic reticulum in astrocytes. Glia 2018, 14, 321042. [Google Scholar] [CrossRef] [Green Version]
- Sadana, R.; Dessauer, C.W. Physiological roles for G protein-regulated adenylyl cyclase isoforms: Insights from knockout and overexpression studies. NeuroSignals 2009, 17, 5–22. [Google Scholar] [CrossRef]
- Efendiev, R.; Dessauer, C.W. AKAPs and Adenylyl Cyclase in Cardiovascular Physiology and Pathology. Acc. Chem. Res. 2008, 45, 788–802. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, T.M.; Lyashkov, A.E.; Zhu, W.; Ruknudin, A.M.; Sirenko, S.; Yang, D.; Deo, S.; Barlow, M.; Johnson, S.; Caffrey, J.L.; et al. High basal protein kinase A-dependent phosphorylation drives rhythmic internal Ca2+ store oscillations and spontaneous beating of cardiac pacemaker cells. Circ. Res. 2006, 98, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Mattick, P.; Parrington, J.; Odia, E.; Simpson, A.; Collins, T.; Terrar, D. Ca2+-stimulated adenylyl cyclase isoform AC1 is preferentially expressed in guinea-pig sino-atrial node cells and modulates the I(f) pacemaker current. J. Physiol. 2007, 582, 1195–1203. [Google Scholar] [CrossRef]
- Lakatta, E.G.; Maltsev, V.A.; Vinogradova, T.M. A coupled SYSTEM of intracellular Ca2+ clocks and surface membrane voltage clocks controls the timekeeping mechanism of the heart’s pacemaker. Circ. Res. 2010, 106, 659–673. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Baldwin, T.A.; Wang, Y.; Subramaniam, J.; Carbajal, A.G.; Brand, C.S.; Cunha, S.R.; Dessauer, C.W. Loss of type 9 adenylyl cyclase triggers reduced phosphorylation of Hsp20 and diastolic dysfunction. Sci. Rep. 2017, 7, 5522. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, T.A. Function of Adenylyl Cyclase in Heart: The AKAP Connection. J. Cardiovasc. Dev. Dis. 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Henry, A.D.; MacQuaide, N.; Burton, F.L.; Rankin, A.C.; Rowan, E.G.; Drummond, R.M. Spontaneous Ca2+transients in rat pulmonary vein cardiomyocytes are increased in frequency and become more synchronous following electrical stimulation. Cell Calcium 2018, 76, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Rietdorf, K.; Bootman, M.D.; Sanderson, M.J. Spontaneous, pro-arrhythmic calcium signals disrupt electrical pacing in mouse pulmonary vein sleeve cells. PLoS ONE 2014, 9, e88649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsushika, S.; Chen, L.; Kawabe, J.; Nilakantan, R.; Halnon, N.J.; Homcy, C.J.; Ishikawa, Y. Cloning and characterization of a sixth adenylyl cyclase isoform: Types V and VI constitute a subgroup within the mammalian adenylyl cyclase family. Proc. Natl. Acad. Sci. USA 1992, 89, 8774–8778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Puri, T.S.; Gerhardstein, B.L.; Chien, A.J.; Green, R.D.; Hosey, M.M. Identification and subcellular localization of the subunits of L-type calcium channels and adenylyl cyclase in cardiac myocytes. J. Biol. Chem. 1997, 272, 19401–19407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, Z.T.; Jiang, M.; Geng, J.; Kaur, S.; Workman, S.W.; Hao, J.; Bernas, T.; Tseng, G.N. Delayed KCNQ1/KCNE1 assembly on the cell surface helps IKs fulfil its function as a repolarization reserve in the heart. J. Physiol. 2021, 599, 3337–3361. [Google Scholar] [CrossRef]
- Crossman, D.J.; Young, A.A.; Ruygrok, P.N.; Nason, G.P.; Baddelely, D.; Soeller, C.; Cannell, M.B. T-tubule disease: Relationship between t-tubule organization and regional contractile performance in human dilated cardiomyopathy. J. Mol. Cell. Cardiol. 2015, 84, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Kashino, Y.; Obara, Y.; Okamoto, Y.; Saneyoshi, T.; Hayashi, Y.; Ishii, K. ERK5 phosphorylates Kv4.2 and inhibits inactivation of the A-type current in PC12 cells. Int. J. Mol. Sci. 2018, 19, 2008. [Google Scholar] [CrossRef] [Green Version]
- Findlay, I. Voltage- and cation-dependent inactivation of L-type Ca2+ channel currents in guinea-pig ventricular myocytes. J. Physiol. 2002, 541, 731–740. [Google Scholar] [CrossRef]
- Brand, C.S.; Hocker, H.J.; Gorfe, A.A.; Cavasotto, C.N.; Dessauer, C.W. Isoform selectivity of adenylyl cyclase inhibitors: Characterization of known and novel compounds. J. Pharmacol. Exp. Ther. 2013, 347, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Gottle, M.; Geduhn, J.; Ko, B.; Gille, A.; Ho, K.; Seifert, R. Characterization of Mouse Heart Adenylyl Cyclase. Jpet 2009, 329, 1156–1165. [Google Scholar] [CrossRef] [Green Version]
- Timofeyev, V.; Myers, R.E.; Kim, H.J.; Woltz, R.L.; Sirish, P.; Heiserman, J.P.; Li, N.; Singapuri, A.; Tang, T.; Yarov-Yarovoy, V.; et al. Adenylyl cyclase subtype-specific compartmentalization: Differential regulation of L-Type Ca2+current in ventricular myocytes. Circ. Res. 2013, 112, 1567–1576. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Brown, M.J. Differential expression of adenylyl cyclase subtypes in human cardiovascular system. Mol. Cell. Endocrinol. 2004, 223, 55–62. [Google Scholar] [CrossRef]
- Stern, M.D. Theory of excitation-contraction coupling in cardiac muscle. Biophys. J. 1992, 63, 497–517. [Google Scholar] [CrossRef] [Green Version]
- Laver, D.R.; Kong, C.H.T.; Imtiaz, M.S.; Cannell, M.B. Termination of calcium-induced calcium release by induction decay: An emergent property of stochastic channel gating and molecular scale architecture. J. Mol. Cell. Cardiol. 2013, 54, 98–100. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Homcy, C.J. The adenylyl cyclases as integrators of transmembrane signal transduction. Circ. Res. 1997, 80, 297–304. [Google Scholar] [CrossRef]
- Cooper, D.M. Regulation and organization of adenylyl cyclases and {cAMP.}. Biochem. J. 2003, 375, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Bakalyar, H.; Randall, R. Identification of a Specialized Adenylyl Cyclase That May Mediate Odorant Detection. Science 1990, 250, 1403–1406. [Google Scholar] [CrossRef]
- Choi, E.J.; Xia, Z.; Storm, D.R. Stimulation of the Type III Olfactory Adenylyl Cyclase by Calciumand Calmodulin. Biochemistry 1992, 31, 6492–6498. [Google Scholar] [CrossRef]
- Trinh, K.; Storm, D.R. Vomeronasal organ detects odorants in absence of signaling through main olfactory epithelium. Nat. Neurosci. 2003, 6, 519–525. [Google Scholar] [CrossRef]
- Wang, Z.; Sindreu, C.B.; Li, V.; Nudelman, A.; Chan, G.C.K.; Storm, D.R. Pheromone detection in male mice depends on signaling through the type 3 adenylyl cyclase in the main olfactory epithelium. J. Neurosci. 2006, 26, 7375–7379. [Google Scholar] [CrossRef]
- Wei, J.; Wayman, G.; Storm, D.R. Phosphorylation and inhibition of type III adenylyl cyclase by calmodulin-dependent protein kinase II in vivo. J. Biol. Chem. 1996, 271, 24231–24235. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.M.F.; Mons, N.; Karpen, J.W. Adenylyl cyclases and the interaction between calcium and cAMP signaling. Nature 1995, 374, 421–424. [Google Scholar] [CrossRef]
- Guillou, J.L.; Nakata, H.; Cooper, D.M.F. Inhibition by calcium of mammalian adenylyl cyclases. J. Biol. Chem. 1999, 274, 35539–35545. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Nakata, H.; Gu, C.; De Beer, T.; Cooper, D.M.F. A critical interplay between Ca2+ inhibition and activation by Mg2+ of AC5 revealed by mutants and chimeric constructs. J. Biol. Chem. 2002, 277, 33139–33147. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, T.M.; Lakatta, E.G. Dual activation of phosphodiesterase 3 and 4 regulates basal cardiac pacemaker function and beyond. Int. J. Mol. Sci. 2021, 22, 8414. [Google Scholar] [CrossRef]
- Tobise, K.; Ishikawa, Y.; Holmer, S.R.; Im, M.J.; Newell, J.B.; Yoshie, H.; Fujita, M.; Susannie, E.E.; Homcy, C.J. Changes in type VI adenylyl cyclase isoform expression correlate with a decreased capacity for cAMP generation in the aging ventricle. Circ. Res. 1994, 74, 596–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, S.; Kawabe, J.I.; Yatani, A.; Takagi, G.; Lee, M.C.; Hong, C.; Liu, J.; Takagi, I.; Sadoshima, J.; Vatner, D.E.; et al. Type 5 adenylyl cyclase disruption alters not only sympathetic but also parasympathetic and calcium-mediated cardiac regulation. Circ. Res. 2003, 93, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Gao, M.H.; Lai, N.C.; Firth, A.L.; Takahashi, T.; Guo, T.; Yuan, J.X.J.; Roth, D.M.; Hammond, H.K. Adenylyl cyclase type 6 deletion decreases left ventricular function via impaired calcium handling. Circulation 2008, 117, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumbay, M.G.; Watts, V.J. Novel regulatory properties of human type 9 adenylate cyclase. J. Pharmacol. Exp. Ther. 2004, 310, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Pálvölgyi, A.; Simpson, J.; Bodnár, I.; Bíró, J.; Palkovits, M.; Radovits, T.; Skehel, P.; Antoni, F.A. Auto-inhibition of adenylyl cyclase 9 (AC9) by an isoform-specific motif in the carboxyl-terminal region. Cell. Signal. 2018, 51, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Hacker, B.M.; Tomlinson, J.E.; Wayman, G.A.; Sultana, R.; Chan, G.; Villacres, E.; Disteche, C.; Storm, D.R. Cloning, chromosomal mapping, and regulatory properties of the human type 9 adenylyl cyclase (ADCY9). Genomics 1998, 50, 97–104. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Kass, R.S.; Dessauer, C.W. The A-kinase anchoring protein yotiao facilitates complex formation between adenylyl cyclase type 9 and the IKs potassium channel in heart. J. Biol. Chem. 2012, 287, 29815–29824. [Google Scholar] [CrossRef] [Green Version]
- Franco, D.; Domínguez, J.; Del Pilar de Castro, M.; Aránega, A. Regulation of myocardial gene expression during heart development. Rev. Esp. Cardiol. 2002, 55, 167–184. [Google Scholar] [CrossRef]
- Bruneau, B.G.; Bao, Z.Z.; Tanaka, M.; Schott, J.J.; Izumo, S.; Cepko, C.L.; Seidman, J.G.; Seidman, C.E. Cardiac expression of the ventricle-specific homeobox gene Irx4 is modulated by Nkx2-5 and dHand. Dev. Biol. 2000, 217, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, O.; Nakagawa, M.; Richardson, J.A.; Olson, E.N.; Srivastava, D. HRT1, HRT2, and HRT3: A new subclass of bHLH transcription factors marking specific cardiac, somitic, and pharyngeal arch segments. Dev. Biol. 1999, 216, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Mommersteeg, M.T.M.; Brown, N.A.; Prall, O.W.J.; De Gier-de Vries, C.; Harvey, R.P.; Moorman, A.F.M.; Christoffels, V.M. Pitx2c and Nkx2-5 are required for the formation and identity of the pulmonary myocardium. Circ. Res. 2007, 101, 902–909. [Google Scholar] [CrossRef] [Green Version]
- Espinoza-lewis, R.A.; Yu, L.; He, F.; Liu, H.; Tang, R.; Shi, J.; Sun, X.; Martin, J.F.; Wang, D.; Yang, J.; et al. Shox2 is essential for the differentiation of cardiac pacemaker cells by repressing Nkx2-5. Dev. Biol. 2009, 327, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, S.; Berger, I.M.; Glaser, A.; Bacon, C.; Li, L.; Gretz, N.; Steinbeisser, H.; Rottbauer, W.; Just, S.; Rappold, G. Islet1 is a direct transcriptional target of the homeodomain transcription factor Shox2 and rescues the Shox2-mediated bradycardia. Basic Res. Cardiol. 2013, 108, 339. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Sequences | Reverse Sequences | |
|---|---|---|
| rAdcy1 | 5’-TATCCTGCTGTTCTCATGCACG-3’ | 5’-TTGGACATGAGGAAGTGCTGTG-3’ |
| rAdcy2 | 5’-ATCATCAGGCATCATCGCCAAC-3’ | 5’-ATTCACTCTGTCTGCCCAGAAC-3’ |
| rAdcy3 | 5’-ATGGTGAAGCTGACACTCATGC-3’ | 5’-AAATGGCGTGCAACATGCTC-3’ |
| rAdcy4 | 5’-TGCCAAGTTCTTCCAGGTCATC-3’ | 5’-TGGGAGTGCAGAAATAGGGAAC-3’ |
| rAdy5 | 5’-ATTGACAGACTGCGATCCGAAC-3’ | 5’-ATGACATGCGTTCACCTGGTTC-3’ |
| rAdcy6 | 5’-ATGGCAGTTTGATGTCTGGTCC-3’ | 5’-ATCGGACGGTGTTAAGTTCAGC-3’ |
| rAdcy7 | 5’-ATGAACAGCCACACCCTTGCTC-3’ | 5’-GCTCCTTCTTAAACTTCTTCT-3’ |
| rAdcy8 | 5’-AGAATACTCTGGCTGCCCTAAC-3’ | 5’-AGCCTCGAAGAAGGAAGCAAAC-3’ |
| rGapdh | 5’-ACCACAGTCCATGCCATCAC-3’ | 5’-TCCACCACCCTGTTGCTGTA-3’ |
| ×109 | LA | LV | PV | ||||
|---|---|---|---|---|---|---|---|
| corresponding abundance | AC1 * | 53,200.08 ± 78,097.71 | (7) | 340,281.06 ± 402,755.99 | (5) | 876,466.31 ±1,272,990.19 | (6) |
| AC2 # | 1.95 ± 2.79 | (6) | 3.38 ± 3.79 | (3) | 3.52 ± 4.59 | (5) | |
| AC3 $ | 80,931.17 ±126,819.78 | (6) | 46,348.88 ± 57,065.14 | (5) | 150,449.16 ± 301,138.14 | (6) | |
| AC4 † | 20.37 ±28.00 | (6) | 9.43 ± 8.09 | (4) | 12.32 ± 12.63 | (5) | |
| AC5 & | 1,451,129.43 ±3,541,141.54 | (8) | 408,442.32 ± 785,280.03 | (5) | 536,885.37 ± 661,839.27 | (5) | |
| AC6 | 549.27 ±730.34 | (8) | 417.79 ± 681.51 | (5) | 810.37 ± 1038.31 | (5) | |
| AC7 | 4051.16 ± 6371.10 | (7) | 393.87 ± 725.97 | (5) | 29,443.41 ± 39,503.56 | (5) | |
| AC8 | 130.21 ± 168.44 | (5) | 35.74 ± 67.52 | (5) | 167.89 ± 205.04 | (5) | |
| Cell Type | [Ca2+]i | Baseline (pA/pF) | Isopreterenol (pA/pF) | Effect (-Fold) | Cm (pF) | n = |
|---|---|---|---|---|---|---|
| PV | [EGTA]i = 5 mM | 9.45 ± 4.18 | 14.27 ± 6.89 | * 1.50 ± 0.18 | 194.80 ± 76.97 | 5 |
| PV | [Ca2+]i = 0.1 mM | 7.07 ± 1.92 | 17.67 ± 3.24 | * 2.59 ± 0.58 | 230.28 ± 40.64 | 5 |
| LV | [EGTA]i = 5 mM | 11.0 ± 3.14 | 21.66 ± 3.97 | 2.05 ± 0.43 | 223.07 ± 51.45 | 5 |
| LV | [Ca2+]i = 0.1 mM | 6.86 ± 2.76 | 12.62 ± 4.96 | 1.91 ± 0.55 | 246.05 ± 40.62 | 5 |
| LA | [EGTA]i = 5 mM | 14.1 ± 2.54 | 21.55 ± 3.12 | # 1.54 ± 0.12 | 117.13 ± 41.96 | 5 |
| LA | [Ca2+]i = 0.1 mM | 9.47 ± 4.49 | 17.74 ± 5.47 | # 1.97 ± 0.30 | 141.76 ± 38.50 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okamoto, Y.; Aung, N.Y.; Tanaka, M.; Takeda, Y.; Takagi, D.; Igarashi, W.; Ishii, K.; Yamakawa, M.; Ono, K. Preferential Expression of Ca2+-Stimulable Adenylyl Cyclase III in the Supraventricular Area, including Arrhythmogenic Pulmonary Vein of the Rat Heart. Biomolecules 2022, 12, 724. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050724
Okamoto Y, Aung NY, Tanaka M, Takeda Y, Takagi D, Igarashi W, Ishii K, Yamakawa M, Ono K. Preferential Expression of Ca2+-Stimulable Adenylyl Cyclase III in the Supraventricular Area, including Arrhythmogenic Pulmonary Vein of the Rat Heart. Biomolecules. 2022; 12(5):724. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050724
Chicago/Turabian StyleOkamoto, Yosuke, Naing Ye Aung, Masahiro Tanaka, Yuji Takeda, Daichi Takagi, Wataru Igarashi, Kuniaki Ishii, Mitsunori Yamakawa, and Kyoichi Ono. 2022. "Preferential Expression of Ca2+-Stimulable Adenylyl Cyclase III in the Supraventricular Area, including Arrhythmogenic Pulmonary Vein of the Rat Heart" Biomolecules 12, no. 5: 724. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050724