

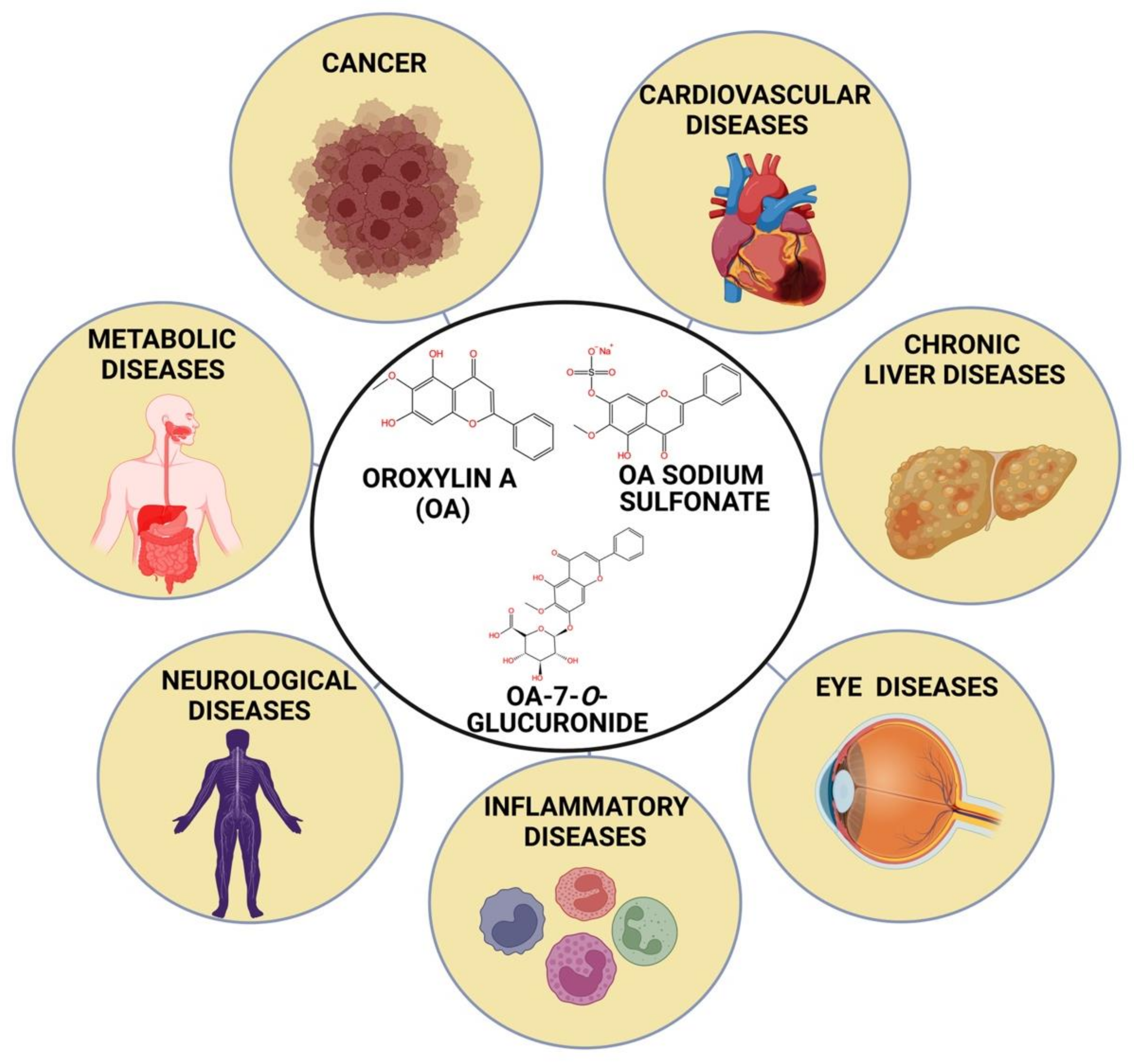
Oroxylin A: A Promising Flavonoid for Prevention and Treatment of Chronic Diseases

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Isolation and Purification of OA
3. Structural Analysis
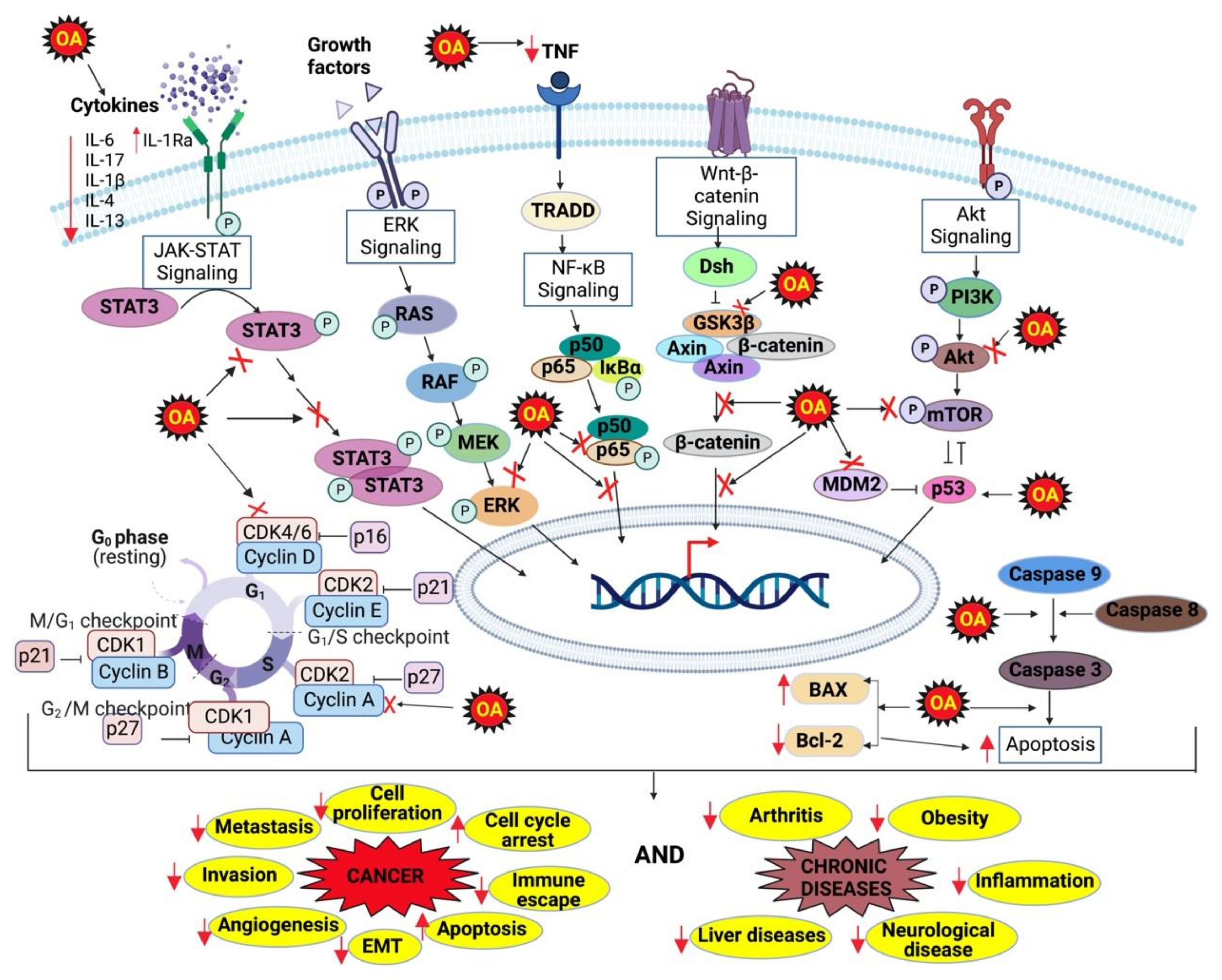
4. Molecular Targets of OA
5. OA for Cancer
6. OA for Cardiovascular Diseases (CVDs)
7. Endotoxemia
8. Hind Limb Ischemia (HLI)
9. OA for Chronic Liver Diseases
9.1. Liver Fibrosis
9.2. Alcohol Liver Disease (ALD)
10. OA for Eye Diseases
11. OA for Inflammatory Diseases
11.1. Allergic Asthma
11.2. Inflammatory Bowel Disease (IBD)
11.3. Osteoarthritis
11.4. Rheumatoid Arthritis (RA)
12. OA for Neurological Disorders
12.1. Attention-Deficit/Hyperactivity Disorder (ADHD)
12.2. Alzheimer’s Disease
12.3. Memory Impairment
13. OA for Obesity
14. OA for Other Diseases
15. Pharmacokinetic Studies of OA
16. Safety and Toxicities of OA
17. Discussion and Conclusions

{kind=link}
{kind=link}
| Disease | In Vitro/ In Vivo | Dose/Conc. | Model | Mechanism of Action or Outcome | References |
|---|---|---|---|---|---|
| Cancer | |||||
| Breast cancer | In vitro | 50, 100, 200 μM | MDA-MB-231, MCF-7 | ↑SIRT3, SOD2, PHD activity, ↓glycolysis, HIF-1α, mitochondrial ROS | [76] |
| In vivo | 100 mg/kg | MDA-MB-231 xenograft | ↑SIRT3, SOD2, ↓tumor volume and mass, glycolysis, HIF-1α, hexokinase II, | [76] | |
| In vitro | 10, 20, 40 μM | MDA-MB-231 | ↑E-cadherin, p27, ↓cell proliferation, CDK2, cyclin E, vimentin, N-cadherin, EMT, migration, invasion, COX-2, NF-κB, IL-6, IL-8, TNF-α | [114] | |
| Cervical cancer | In vitro | 5, 20, 80 μM | HeLa | ↑Procaspase-3, procaspase-8, procaspase-9, cleaved PARP, apoptosis ↓Bcl-2, cell growth | [75] |
| In vivo | 40, 80 mg/kg | HeLa xenograft | ↑Cleaved PARP, ↓tumor growth, Bcl-2, procaspase-3, procaspase-8, procaspase-9 | [75] | |
| Colon cancer | In vitro | 200 μM/L | HT-29 cells | ↑Bax, p53, PARP, procaspase-3, ROS, ↓COX-2, Bcl-2, PGE2 | [118] |
| In vivo | 100 mg/kg | HT-29 | ↓Tumor, COX-2 | [118] | |
| In vitro | 100 μM/L | HCT-116 | ↑Caspase-3, caspase-9, Bax, ROS, Nrf2, HO-1, NQO1, ↓Bcl-2, | [56] | |
| In vivo | 50, 100, 200 mg/kg | HCT-116 xenograft | ↑Nrf-2, apoptosis, ↓tumor growth | [56] | |
| ESCC | In vitro | 10, 50 μM | TE13, ECA109 | ↑Apoptosis, G2/M arrest, radiosensitization, ↓cyclin B1, cdc2 | [136] |
| Glioma | In vitro | 25, 50, 75, 100, 125, 150, 175, 200 μM | U251, U118, U87 | ↑Autophagy, Beclin, ↓Akt, ERK Notch-1, Mcl-1 | [126] |
| In vitro | 50 μM | C6, U251 | ↑Apoptosis, ↓p-Akt, β-catenin, IP3R1, p-GSK-3β | [94] | |
| In vivo | 150 mg/kg | C6 xenograft | ↑Apoptosis, ↓Akt/β-catenin, IP3R1, p-GSK-3β, | [94] | |
| Hematological malignancies | In vitro | 60 μM | K562, KU812 with M2-10B4 | ↑Apoptosis, ↓CXCL12/CXCR7, p-ERK, p-BAD, survivin | [130] |
| In vivo | 200 mg/kg | K562 xenograft | ↑Apoptosis, ↓CXCR7, p-ERK, CD13+ cells | [130] | |
| In vitro | 20 μM | HL-60, NB4 | ↑TNF-α sensitivity, ↓tRXRα, PI3K/Akt, NF-κB | [83] | |
| In vivo | 80 mg/kg | AML cell xenograft | ↑Survival, ↓NF-κB, AML cell population | [83] | |
| In vitro | 10–160 μM | t (8i21)-positive kasumi-l, primary AML cells | ↑C/EBPα, p21, CD11b/CD14, ↓AML 1/ETO, HDAC-1 | [129] | |
| In vivo | 200 mg/kg | NOD/SCID mice | ↑Survival, ↓HDAC-1, AML1/ETO, CD45+ cells, | [129] | |
| Hepatocellular carcinoma | In vitro | 50 μM | HepG2 cells | ↑Apoptosis, p53, cleaved PARP, ↓Cell viability, TS and DPD mRNA, COX-2, Bcl-2, procaspase-3 | [119] |
| In vivo | 1000 mg/kg/day | H22 xenograft | ↓Tumor growth, tumor weight | [119] | |
| In vitro | 12.5, 25, 50 μM | SMMC-7721 | ↑NAG1, acetylation of C/EBPβ, ↓migration, invasion, EMT, p-SMAD2/3, TGF-β1/SMAD axis | [120] | |
| In vivo | 200 mg/kg | SMMC-7721 | ↑E-cadherin, ↓pulmonary metastasis, vimentin, twist1 | [120] | |
| Kaposi’s sarcoma | In vitro | 20–1000 μM | KSHVvIL-6 | ↑Apoptosis, PPARγ, invasion, neovascularization, ↓Prox1, VEGFR3, LYVE-1, podoplanin | [80] |
| Lung cancer | In vitro | 40 μM | H460 | ↓Tregs, TGF-β, NF-κB | [134] |
| In vivo | 60 mg/kg | H460 xenograft | ↓Tumor, Tregs, FOXP3, | [134] | |
| In vitro | 50 μM/L | H460, A549, 95D, PC9, HCC827, H1975 | ↑Apoptosis, ↓tumor, XPC transcription | [135] | |
| In vivo | 50 mg/kg | H460 xenografts | ↑Cisplatin sensitivity, ↓tumor growth, Ki67, PCNA, XPC expression | [135] | |
| Skin cancer | In vitro | 20 μM | JB6P+ | ↓Transformation, inflammation, SHCBP1, NF-κB p65, IL-1β, IL-6, IL-18, TNF-α, COX-2, iNOS, | [137] |
| In vivo | 40 mg/kg | DMBA/TPA mice | ↓SHCBP1, IL-1β, IL-4, IL-6, IL-18, TNF-α, NLRP3, PCNA tumorigenesis, incidence rate, tumor multiplicity, epidermal thickness | [137] | |
| Cardiovascular diseases | |||||
| Cardioprotective effects | In vivo | 40 mg/kg | C57BL/6 mice | ↑Body weight, SIRT1, cAMP/protein kinase A, improved contractile function, Nrf2, HO-1, NQO1, Bcl-2, ↓plasma and cardiac CK-MB, LDH, LVEDP, 4-HNE, nitrotyrosine, gp91phox, NADPH oxidase 4, p47phox, p67phox, IL-6, IL-1β, MMP-2, MMP-9, p-IκBα, caspase 3/7 activity, PARP activity, apoptosis | [143] |
| Endotoxemia | In vivo | 10, 20 μM | Sprague-Dawley rats | ↑Coronary flow, LVDP ↓CPP | [55] |
| Hind limb ischemia | In vivo | 10 mg/kg/day | C57BL/6 mice | ↑VEGFA, Ang-2, FGF-2, PDGF-BB, angiogenesis, perfusion recovery, regeneration of myocytes ↓IL-1β, tissue injury, ischemia, apoptosis of myocytes | [153] |
| Chronic liver diseases | |||||
| Acute liver injury | In vivo | 60 mg/kg | C57 BL/6 mice | ↑IL-1Ra, HGF, EGF, PCNA positive cells, survival, ↓IL-1β, IL-6, TNF-α, necrotic areas | [69] |
| In vivo | 15, 30, 60 mg/kg | BALB/c mice | ↑Nrf 2, HO-1, ↓AST, ALT, TNF-α, MDA, MPO activity, NF-κB, TLR4, necrosis | [159] | |
| ALD | In vitro | 10–100 μM | LO2 cells | ↑YAP, ↓AST, ALT, LDH, p21, p16 and HMGA1 | [169] |
| In vivo | 30 mg/kg | ICR mice | ↑YAP, ↓AST, ALT, ALP, lipid vacuolation | [169] | |
| In vitro | 10, 20, 40 μM | LO2 cells | ↑Mfn2, PGC-1α, ↓LDH, IL-1β, IL-18, caspase-1, NF-κB, ROS, NLRP3 inflammasome | [77] | |
| In vivo | 40 mg/kg | ICR mice | ↓Inflammation, lipid accumulation, ALT, ALP, AST | [77] | |
| Hepatic steatosis | In vitro | 10, 20, 40 μM | LO2 cells | ↑CPT1, PPARα, PPARγ, ↓lipid droplet accumulation, HIF-1α, apoptosis, SREBP1, FAS, SCD1 | [59] |
| In vivo | 30 mg/kg | ICR mice | ↓ Apoptosis, ALT, AST, ALP, IL-6, IL-8, TNF-α | [59] | |
| Hepatic fibrosis | In vitro | 20, 30, 40 μM | HSCs | ↑LC3-B, Atg3, Atg4, Atg5, Atg7, Atg9, beclin, Atg12, Atg14, ↓α-SMA, desmin, α1collagen, fibronectin, TGF-β, TNF-α, p62 | [162] |
| In vivo | 20, 30, 40 mg/kg | ICR mice | ↑LC3-B, Atg5, beclin1, ↓AST, ALT, ALP, α-SMA, α1collagen, fibronectin, PDGF-βR, TGF-βR1, p62, fibrosis lesions, necrosis, inflammation | [162] | |
| In vitro | 20, 30, 40 μM | HSCs | ↓Hexokinase II, LDH-A, PFK1, PKM2, actin stress fibers, p-MLC2, contraction | [163] | |
| In vivo | 40 mg/kg | ICR mice | ↓Liver injury, glycolysis, α-SMA, α1collagen, fibronectin, ALT, AST, TBIL, IBIL, p-MLC2 | [163] | |
| In vitro | 20, 30, 40 μM | HSC | ↑SLC7A11, GSH, lipid droplet content, retinol, cholesterol, triglyceride ↓ATGL, α-SMA, α1collagen, fibronectin, desmin, ROS | [79] | |
| In vivo | 20 mg/kg | C57BL/6 | ↑Antioxidant activity, ↓liver fibrosis, collagen deposition lipid droplet content, retinol, cholesterol, triglyceride, α-SMA, collagen I | [79] | |
| In vitro | 20, 30, 40 μM | HSC | ↑Autophagy, Atg5, Atg12, beclin, LC3B, ↓ NF-κB, NLRP3, TNF-α, IL-1β, p-PI3K, p-Akt p-mTOR, ROS, p62, IL-1β, IL-4, IL-6, IL18, TNF-α, IFNγ | [92] | |
| In vivo | 20, 30, 40 mg/kg | C57BL/6 | ↓ NF-κB, α-SMA, IL-1β, IL-4, IL-6, IL18, TNF-α, IFNγ | [92] | |
| In vitro | 20, 30, 40 μM | LSECs | ↓VEGF-A, angiogenesis | [256] | |
| In vivo | 40 mg/kg | ICR mice | ↓VEGF-A, Ang-2, CD31, HIF-1α | [256] | |
| In vitro | 20, 30, 40 μM | HSC | ↑Cleaved caspase-9, cleaved caspase-3, cleaved PARP, p51, p21, p27, S-phase arrest, Bax, collagen degradation, MMP-9, ATF4, p-PERK, cleaved ATF6, ↓Fibrogenesis, PDGF-β, TGF-β, EGFR, cyclin A, cyclin E, CDK-2, Bcl-2, collagen synthesis, TIMP-2, α-SMA, collagen I | [165] | |
| In vivo | 20, 30, 40 mg/kg | ICR mice | ↑ERS pathway, IL-6, IL18, TNF-α, AST, ALT | [165] | |
| Inflammatory diseases | |||||
| Allergic asthma | In vitro | 0.1, 0.3, 1, 3, 10, 30 μM | RBL-2H3 mast cells | ↓β-Hexosaminidase release, antigen-induced degranulation | [70] |
| In vivo | 5 mg/kg | Female BALB/c mice | ↓IFNγ, IL-2, IL-4, IL-5, IL-13, eosinophils, inflammation score, mucin | [70] | |
| In vivo | 15, 30, 60 mg/kg | BALB/c mice | ↓ IgE, p-IκB, p-NF-κB, IL-4, IL-5, IL-13, airway hyporesponsiveness, inflammatory cells infiltration, thickening of alveolar wall | [183] | |
| Inflammatory bowel disease | In vivo | 50 mg/kg | BALB/c mice | ↓Inflammation, IL-1β, IL-6, IL-17, TNF- α, Muc2, IFNγ | [189] |
| In vivo | 100, 200 Mg/kg | BALB/c mice | ↓Distribution of CD11b+ inflammatory cells and F4/80+ macrophages, MPO, iNOS, NLRP3, IL-1β, IL-6, TNF- α, p65 | [190] | |
| Lung inflammation | In vitro | 50, 100, 150 μM | BEAS-2B and RAW 2647 cells | ↑Nrf2, GSH, HO-1, ↓TNF-α, IL-1β | [143] |
| In vivo | 15, 30, 60 mg/kg | C57BL/6 | ↑GR activity, GSH, ↓interstitial edema, infiltrated immune cells, alveolar wall thickness, TNF α, IL-1β, MCP-1, 3-nitrotyrosine, 8-OHdG, 8-isoprostane | [143] | |
| Rheumatoid arthritis | In vitro | 1, 4, 16 μM | FLS cells | ↑IL-10, ↓IL-1β, IL-6, p-ERK, p-MAPK, p65 | [202] |
| In vivo | 10 mg/kg | DBA/1 mice with CIA | ↑Tregs ↓total IgG, IgG1, IgG2a, IgG2b, IL-1β, IL-6, IL-17, TNF-α, arthritis score, swelling, joint inflammation, Th17 cells | [202] | |
| Osteoarthritis | In vitro | 2.5, 5, 10, 20, 50 μM | Chondrocytes | ↓IL-1β, MMP-13, ADAMTS-5, NO, PGE2, ALP IL-6, TNF-α, NF-κB, RUNX-2, collagen X, β-catenin | [196] |
| In vivo | 10 mg/kg | OA-induced mice model | ↓OARSI score | [196] | |
| In vitro | 2–128 μM | Chondrocytes | ↓NOS, cox-2, MMP-3, MMP-13, ERK1/2, PI3K/Akt | [72] | |
| Obesity | In vitro | 25, 50, 75 μM | 3T3-L1 pre-adipocytes | ↓Lipid accumulation, adipogenesis, PPARγ, C/EBPα | [225] |
| Neurological diseases | |||||
| ADHD | In vivo | 5, 10 mg/kg | SHR, WKY | ↓Drinking attempts, drinking frequency, dopamine reuptake | [209] |
| Alzheimer’s disease | In vitro | 10, 50, 100 μM | PC12 cells | ↓Ca2+, Bax, iNOS, cleaved caspase-8, cleaved PARP-1, TNF-α, NO, PGE2, p-IκBα, cox-2, p-NF-κB, p-p38, p-JNK, ROS, apoptosis, cell cycle arrest, | [71] |
| Memory impairment | In vivo | 5 mg/kg | ICR mice | ↑ChAT, ↓Nissl bodies, OX-42 positive cells, GFAP positive cells, iNOS, spontaneous alteration behavior, micro glial cell activation, lipid peroxidation | [218] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kumar, A.; Harsha, C.; Parama, D.; Girisa, S.; Daimary, U.D.; Mao, X.; Kunnumakkara, A.B. Current clinical developments in curcumin-based therapeutics for cancer and chronic diseases. Phytother. Res. 2021, 35, 6768–6801. [Google Scholar] [CrossRef] [PubMed]
- Parama, D.; Girisa, S.; Khatoon, E.; Kumar, A.; Alqahtani, M.S.; Abbas, M.; Sethi, G.; Kunnumakkara, A.B. An overview of the pharmacological activities of scopoletin against different chronic diseases. Pharmacol. Res. 2022, 179, 106202. [Google Scholar] [CrossRef] [PubMed]
- Heymach, J.V.; Shackleford, T.J.; Tran, H.T.; Yoo, S.Y.; Do, K.A.; Wergin, M.; Saintigny, P.; Vollmer, R.T.; Polascik, T.J.; Snyder, D.C.; et al. Effect of low-fat diets on plasma levels of NF-kappaB-regulated inflammatory cytokines and angiogenic factors in men with prostate cancer. Cancer Prev. Res. 2011, 4, 1590–1598. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Sailo, B.L.; Roy, N.K.; Thakur, K.K.; Banik, K.; Shakibaei, M.; Gupta, S.C.; Aggarwal, B.B. Cancer drug development: The missing links. Exp. Biol. Med. 2019, 244, 663–689. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Yang, S.F.; Sethi, G.; Hu, D.N. Natural bioactives in cancer treatment and prevention. Biomed. Res. Int. 2015, 2015, 182835. [Google Scholar] [CrossRef]
- Kirtonia, A.; Gala, K.; Fernandes, S.G.; Pandya, G.; Pandey, A.K.; Sethi, G.; Khattar, E.; Garg, M. Repurposing of drugs: An attractive pharmacological strategy for cancer therapeutics. Semin. Cancer Biol. 2021, 68, 258–278. [Google Scholar] [CrossRef]
- Deldar Abad Paskeh, M.; Asadi, S.; Zabolian, A.; Saleki, H.; Khoshbakht, M.A.; Sabet, S.; Naghdi, M.J.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; et al. Targeting cancer stem cells by dietary agents: An important therapeutic strategy against human malignancies. Int. J. Mol. Sci. 2021, 22, 11669. [Google Scholar] [CrossRef]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef]
- Manu, K.A.; Shanmugam, M.K.; Li, F.; Chen, L.; Siveen, K.S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J. Mol. Med. 2014, 92, 267–276. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Kim, S.H.; Sethi, G.; Ahn, K.S. Farnesol inhibits tumor growth and enhances the anticancer effects of bortezomib in multiple myeloma xenograft mouse model through the modulation of STAT3 signaling pathway. Cancer Lett. 2015, 360, 280–293. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Banik, K.; Girisa, S.; Shabnam, B.; Shakibaei, M.; Fan, L.; Arfuso, F.; Monisha, J.; Wang, H.; Mao, X.; et al. The vital role of ATP citrate lyase in chronic diseases. J. Mol. Med. 2020, 98, 71–95. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Parama, D.; Daimari, E.; Girisa, S.; Banik, K.; Harsha, C.; Dutta, U.; Kunnumakkara, A.B. Rationalizing the therapeutic potential of apigenin against cancer. Life Sci. 2021, 267, 118814. [Google Scholar] [CrossRef]
- Bordoloi, D.; Banik, K.; Shabnam, B.; Padmavathi, G.; Monisha, J.; Arfuso, F.; Dharmarajan, A.; Mao, X.; Lim, L.H.K.; Wang, L.; et al. TIPE family of proteins and its implications in different chronic diseases. Int. J. Mol. Sci. 2018, 19, 2974. [Google Scholar] [CrossRef]
- Padmavathi, G.; Roy, N.K.; Bordoloi, D.; Arfuso, F.; Mishra, S.; Sethi, G.; Bishayee, A.; Kunnumakkara, A.B. Butein in health and disease: A comprehensive review. Phytomedicine 2017, 25, 118–127. [Google Scholar] [CrossRef]
- Shabnam, B.; Padmavathi, G.; Banik, K.; Girisa, S.; Monisha, J.; Sethi, G.; Fan, L.; Wang, L.; Mao, X.; Kunnumakkara, A.B. Sorcin a potential molecular target for cancer therapy. Transl. Oncol. 2018, 11, 1379–1389. [Google Scholar] [CrossRef]
- Buhrmann, C.; Kunnumakkara, A.B.; Kumar, A.; Samec, M.; Kubatka, P.; Aggarwal, B.B.; Shakibaei, M. Multitargeting effects of Calebin A on malignancy of CRC cells in multicellular tumor microenvironment. Front. Oncol. 2021, 11, 650603. [Google Scholar] [CrossRef]
- Babu, B.H.; Jayram, H.N.; Nair, M.G.; Ajaikumar, K.B.; Padikkala, J. Free radical scavenging, antitumor and anticarcinogenic activity of gossypin. J. Exp. Clin. Cancer Res. 2003, 22, 581–589. [Google Scholar]
- Mastron, J.K.; Siveen, K.S.; Sethi, G.; Bishayee, A. Silymarin and hepatocellular carcinoma: A systematic, comprehensive, and critical review. Anticancer Drugs 2015, 26, 475–486. [Google Scholar] [CrossRef]
- Patel, S.M.; Nagulapalli Venkata, K.C.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Zhang, J.; Ahn, K.S.; Kim, C.; Shanmugam, M.K.; Siveen, K.S.; Arfuso, F.; Samym, R.P.; Deivasigamanim, A.; Lim, L.H.; Wang, L.; et al. Nimbolide-induced oxidative stress abrogates STAT3 signaling cascade and inhibits tumor growth in transgenic adenocarcinoma of mouse prostate model. Antioxid. Redox Signal 2016, 24, 575–589. [Google Scholar] [CrossRef]
- Lee, J.H.; Chiang, S.Y.; Nam, D.; Chung, W.S.; Lee, J.; Na, Y.S.; Sethi, G.; Ahn, K.S. Capillarisin inhibits constitutive and inducible STAT3 activation through induction of SHP-1 and SHP-2 tyrosine phosphatases. Cancer Lett. 2014, 345, 140–148. [Google Scholar] [CrossRef]
- Roy, N.K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A.K.; Thakur, K.K.; Padmavathi, G.; Shakibaei, M.; Fan, L.; Sethi, G.; et al. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. Int. J. Mol. Sci. 2019, 20, 74101. [Google Scholar] [CrossRef]
- Shishodia, S.; Sethi, G.; Ahn, K.S.; Aggarwal, B.B. Guggulsterone inhibits tumor cell proliferation, induces S-phase arrest, and promotes apoptosis through activation of c-Jun N-terminal kinase, suppression of Akt pathway, and downregulation of antiapoptotic gene products. Biochem. Pharmacol. 2007, 74, 118–130. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Banik, K.; Bordoloi, D.; Harsha, C.; Sailo, B.L.; Padmavathi, G.; Roy, N.K.; Gupta, S.C.; Aggarwal, B.B. Googling the guggul (commiphora and boswellia) for prevention of chronic diseases. Front. Pharm. 2018, 9, 686. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Bordoloi, D.; Thakur, K.K.; Monisha, J.; Arfuso, F.; Sethi, G.; Mishra, S.; Kumar, A.P.; Kunnumakkara, A.B. Possible use of Punica granatum (Pomegranate) in cancer therapy. Pharmacol. Res. 2018, 133, 53–64. [Google Scholar] [CrossRef]
- Moballegh Nasery, M.; Abadi, B.; Poormoghadam, D.; Zarrabi, A.; Keyhanvar, P.; Khanbabaei, H.; Ashrafizadeh, M.; Mohammadinejad, R.; Tavakol, S.; Sethi, G. Curcumin Delivery Mediated by Bio-Based Nanoparticles: A Review. Molecules 2020, 25, 689. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Sung, B.; Ravindran, J.; Diagaradjane, P.; Deorukhkar, A.; Dey, S.; Koca, C.; Tong, Z.; Gelovani, J.G.; Guha, S.; et al. Zyflamend suppresses growth and sensitizes human pancreatic tumors to gemcitabine in an orthotopic mouse model through modulation of multiple targets. Int. J. Cancer 2012, 131, E292–E303. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Nair, A.S.; Ahn, K.S.; Pandey, M.K.; Yi, Z.; Liu, M.; Aggarwal, B.B. Gossypin, a pentahydroxy glucosyl flavone, inhibits the transforming growth factor beta-activated kinase-1-mediated NF-kappaB activation pathway, leading to potentiation of apoptosis, suppression of invasion, and abrogation of osteoclastogenesis. Blood 2007, 109, 5112–5121. [Google Scholar] [CrossRef]
- Girisa, S.; Kumar, A.; Rana, V.; Parama, D.; Daimary, U.D.; Warnakulasuriya, S.; Kumar, A.P.; Kunnumakkara, A.B. From simple mouth cavities to complex oral mucosal disorders—curcuminoids as a promising therapeutic approach. ACS Pharmacol. Transl. Sci. 2021, 4, 647–665. [Google Scholar] [CrossRef]
- Daimary, U.D.; Parama, D.; Rana, V.; Banik, K.; Kumar, A.; Harsha, C.; Kunnumakkara, A.B. Emerging roles of cardamonin, a multitargeted nutraceutical in the prevention and treatment of chronic diseases. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100008. [Google Scholar] [CrossRef]
- Khwairakpam, A.D.; Damayenti, Y.D.; Deka, A.; Monisha, J.; Roy, N.K.; Padmavathi, G.; Kunnumakkara, A.B. Acorus calamus: A bio-reserve of medicinal values. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Devi Daimary, U.; Girisa, S.; Parama, D.; Verma, E.; Kumar, A.; Kunnumakkara, A.B. Embelin: A novel XIAP inhibitor for the prevention and treatment of chronic diseases. J. Biochem. Mol. Toxicol. 2022, 36, e22950. [Google Scholar] [CrossRef] [PubMed]
- Jude, S.; Amalraj, A.; Kunnumakkara, A.B.; Divya, C.; Löffler, B.M.; Gopi, S. Development of validated methods and quantification of curcuminoids and curcumin metabolites and their pharmacokinetic study of oral administration of complete natural turmeric formulation (Cureit™) in human plasma via UPLC/ESI-Q-TOF-MS spectrometry. Molecules 2018, 23, 2415. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, J.H.; Sethi, G.; Kim, C.; Baek, S.H.; Nam, D.; Chung, W.S.; Kim, S.H.; Shim, B.S.; Ahn, K.S. Bergamottin, a natural furanocoumarin obtained from grapefruit juice induces chemosensitization and apoptosis through the inhibition of STAT3 signaling pathway in tumor cells. Cancer Lett. 2014, 354, 153–163. [Google Scholar] [CrossRef]
- Gopi, S.; Jacob, J.; Varma, K.; Jude, S.; Amalraj, A.; Arundhathy, C.; George, R.; Sreeraj, T.; Divya, C.; Kunnumakkara, A.B. Comparative oral absorption of curcumin in a natural turmeric matrix with two other curcumin formulations: An open-label parallel-arm study. Phytother. Res. 2017, 31, 1883–1891. [Google Scholar] [CrossRef]
- Olennikov, D.; Chirikova, N.; Tankhaeva, L. Phenolic compounds of Scutellaria baicalensis Georgi. Russ. J. Bioorganic Chem. 2010, 36, 816–824. [Google Scholar] [CrossRef]
- Li, H.-B.; Chen, F. Isolation and purification of baicalein, wogonin and oroxylin A from the medicinal plant Scutellaria baicalensis by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1074, 107–110. [Google Scholar] [CrossRef]
- Li, J.; Ding, Y.; Li, X.C.; Ferreira, D.; Khan, S.; Smillie, T.; Khan, I.A. Scuteflorins A and B, dihydropyranocoumarins from Scutellaria lateriflora. J. Nat. Prod. 2009, 72, 983–987. [Google Scholar] [CrossRef]
- Dinda, B.; SilSarma, I.; Dinda, M.; Rudrapaul, P. Oroxylum indicum (L.) Kurz, an important Asian traditional medicine: From traditional uses to scientific data for its commercial exploitation. J. Ethnopharmacol. 2015, 161, 255–278. [Google Scholar] [CrossRef]
- Sajeev, A.; Hegde, M.; Daimary, U.D.; Kumar, A.; Girisa, S.; Sethi, G.; Kunnumakkara, A.B. Modulation of diverse oncogenic signaling pathways by oroxylin A: An important strategy for both cancer prevention and treatment. Phytomedicine 2022, 105, 154369. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Herbal Remedies: Rational Western Therapy, Ayurvedic, and Other Traditional Usage, Botany; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Bhattacharjee, S.K. Use Of Flavours And Fragrances. In Handbook Of Aromatic Plants, 2nd ed.; Pointer Publishers: Jaipur, India, 2005. [Google Scholar]
- Zaveri, M.N.; Khandhar, A.P.; Jain, S.M. Quantification of baicalein, chrysin, biochanin-A and ellagic acid in root bark of Oroxylum indicum by RP-HPLC with UV detection. Eurasian J. Anal. Chem. 2008, 3, 245–257. [Google Scholar]
- Jabbar, S.; Khan, M.T.; Choudhuri, M.S.; Sil, B.K. Bioactivity studies of the individual ingredients of the Dashamularishta. Pak. J. Pharm. Sci. 2004, 17, 9–17. [Google Scholar]
- Singh, V.; Chaudhary, A.K. A review on the taxonomy, ethnobotany, chemistry and pharmacology of Oroxylum indicum vent. Indian J. Pharm. Sci. 2011, 73, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharm. 2019, 71, 1353–1369. [Google Scholar] [CrossRef]
- Awad, R.; Arnason, J.T.; Trudeau, V.; Bergeron, C.; Budzinski, J.W.; Foster, B.C.; Merali, Z. Phytochemical and biological analysis of skullcap (Scutellaria lateriflora L.): A medicinal plant with anxiolytic properties. Phytomedicine 2003, 10, 640–649. [Google Scholar] [CrossRef]
- Zhang, Z.; Lian, X.Y.; Li, S.; Stringer, J.L. Characterization of chemical ingredients and anticonvulsant activity of American skullcap (Scutellaria lateriflora). Phytomedicine 2009, 16, 485–493. [Google Scholar] [CrossRef]
- Lohani, M.; Ahuja, M.; Buabeid, M.A.; Dean, S.; Dennnis, S.; Suppiramaniam, V.; Kemppainen, B.; Dhanasekaran, M. Anti-oxidative and DNA protecting effects of flavonoids-rich Scutellaria lateriflora. Nat. Prod. Commun. 2013, 8, 1415–1418. [Google Scholar] [CrossRef]
- Rojsanga, P.; Bunsupa, S.; Sithisarn, P. Flavones contents in extracts from Oroxylum indicum seeds and plant tissue cultures. Molecules 2020, 25, 1545. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharm. 2021, 133, 110917. [Google Scholar] [CrossRef]
- Shah, R.; Mehta, C.; Wheeler, T. 131. The constitution of oroxylin-A, a yellow colouring matter from the root-bark of Oroxylum indicum, vent. J. Chem. Soc. 1936, 591–593. [Google Scholar] [CrossRef]
- Pal-Bhadra, M.; Ramaiah, M.J.; Reddy, T.L.; Krishnan, A.; Pushpavalli, S.; Babu, K.S.; Tiwari, A.K.; Rao, J.M.; Yadav, J.S.; Bhadra, U. Plant HDAC inhibitor chrysin arrest cell growth and induce p21 WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer 2012, 12, 180. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, Y.; Li, Z.; Zhang, H.; Yang, Y.; Qin, H.; Xu, Q.; Zhao, L. Oroxylin A reversed Fibronectin-induced glioma insensitivity to Temozolomide by suppressing IP3R1/AKT/β-catenin pathway. Life Sci. 2020, 260, 118411. [Google Scholar] [CrossRef]
- Liu, C.H.; Chen, M.F.; Tseng, T.L.; Chen, L.G.; Kuo, J.S.; Lee, T.J. Oroxylin a, but not vasopressin, ameliorates cardiac dysfunction of endotoxemic rats. Evid. Based Complementary Altern. Med. 2012, 2012, 408187. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Chen, N.; Yao, J.; Zhao, Q.; Zhang, F.; Li, Z.Y.; You, Q.D.; Guo, Q.L. The role of Nrf2 and apoptotic signaling pathways in oroxylin A-mediated responses in HCT-116 colorectal adenocarcinoma cells and xenograft tumors. Anti-Cancer Drugs 2012, 23, 651–658. [Google Scholar] [CrossRef]
- Jeon, S.J.; Bak, H.; Seo, J.; Han, S.M.; Lee, S.H.; Han, S.H.; Kwon, K.J.; Ryu, J.H.; Cheong, J.H.; Ko, K.H.; et al. Oroxylin A induces BDNF expression on cortical neurons through adenosine A2A receptor stimulation: A possible role in neuroprotection. Biomol. Ther. 2012, 20, 27–35. [Google Scholar] [CrossRef]
- Jin, J.; Chen, S.; Wang, D.; Chen, Y.; Wang, Y.; Guo, M.; Zhou, C.; Dou, J. Oroxylin A suppresses influenza A virus replication correlating with neuraminidase inhibition and induction of IFNs. Biomed. Pharm. 2018, 97, 385–394. [Google Scholar] [CrossRef]
- Jin, H.; Lian, N.; Bian, M.; Zhang, C.; Chen, X.; Shao, J.; Wu, L.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A prevents alcohol-induced hepatic steatosis through inhibition of hypoxia inducible factor 1alpha. Chem. Biol. Interact. 2018, 285, 14–20. [Google Scholar] [CrossRef]
- Lu, Z.; Lu, N.; Li, C.; Li, F.; Zhao, K.; Lin, B.; Guo, Q. Oroxylin A inhibits matrix metalloproteinase-2/9 expression and activation by up-regulating tissue inhibitor of metalloproteinase-2 and suppressing the ERK1/2 signaling pathway. Toxicol. Lett. 2012, 209, 211–220. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, D.; Zhang, Y.; Dong, J.; Wang, J.; Niu, X. Molecular modeling reveals the novel inhibition mechanism and binding mode of three natural compounds to staphylococcal α-hemolysin. PLoS ONE 2013, 8, e80197. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, N.; Ling, Y.; Gao, Y.; Chen, Y.; Wang, L.; Hu, R.; Qi, Q.; Liu, W.; Yang, Y.; et al. Oroxylin A suppresses invasion through down-regulating the expression of matrix metalloproteinase-2/9 in MDA-MB-435 human breast cancer cells. Eur. J. Pharm. 2009, 603, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Banik, K.; Kunnumakkara, A.B.; Kubatka, P.; Koklesova, L.; Shakibaei, M. Targeting NF-κB signaling by calebin a, a compound of turmeric, in multicellular tumor microenvironment: Potential role of apoptosis induction in CRC cells. Biomedicines 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Harsha, C.; Banik, K.; Ang, H.L.; Girisa, S.; Vikkurthi, R.; Parama, D.; Rana, V.; Shabnam, B.; Khatoon, E.; Kumar, A.P.; et al. Targeting AKT/mTOR in oral cancer: Mechanisms and advances in clinical trials. Int. J. Mol. Sci. 2020, 21, 3285. [Google Scholar] [CrossRef]
- Muralimanoharan, S.B.; Kunnumakkara, A.B.; Shylesh, B.; Kulkarni, K.H.; Haiyan, X.; Ming, H.; Aggarwal, B.B.; Rita, G.; Kumar, A.P. Butanol fraction containing berberine or related compound from Nexrutine® inhibits NFκB signaling and induces apoptosis in prostate cancer cells. Prostate 2009, 69, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Bordoloi, D.; Banik, K.; Padmavathi, G.; Vikkurthi, R.; Harsha, C.; Roy, N.K.; Singh, A.K.; Monisha, J.; Wang, H.; Kumar, A.P.; et al. TIPE2 induced the proliferation, survival, and migration of lung cancer cells through modulation of Akt/mTOR/NF-κB signaling cascade. Biomolecules 2019, 9, 836. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kunnumakkara, A.B. Molecular Targets and Therapeutic Uses of Spices: Modern Uses for Ancient Medicine; World Scientific: Singapore, 2009. [Google Scholar]
- Lu, L.; Guo, Q.; Zhao, L. Overview of oroxylin A: A promising flavonoid compound. Phytother. Res. 2016, 30, 1765–1774. [Google Scholar] [CrossRef]
- Zhu, R.; Zeng, G.; Chen, Y.; Zhang, Q.; Liu, B.; Liu, J.; Chen, H.; Li, M. Oroxylin A accelerates liver regeneration in CCl4-induced acute liver injury mice. PLoS ONE 2013, 8, e71612. [Google Scholar] [CrossRef]
- Lee, A.Y.; Kang, S.; Park, S.J.; Huang, J.; Im, D.S. Anti-allergic effect of Oroxylin A from Oroxylum indicum using in vivo and in vitro experiments. Biomol. Ther. 2016, 24, 283–290. [Google Scholar] [CrossRef]
- Ji, Y.; Han, J.; Lee, N.; Yoon, J.H.; Youn, K.; Ha, H.J.; Yoon, E.; Kim, D.H.; Jun, M. Neuroprotective effects of Baicalein, Wogonin, and Oroxylin A on Amyloid Beta-Induced Toxicity via NF-κB/MAPK pathway modulation. Molecules 2020, 25, 5087. [Google Scholar] [CrossRef]
- Zhang, Y.; Weng, Q.; Chen, J.; Li, M.; Han, J. Oroxylin A attenuates IL-1β-induced inflammatory reaction via inhibiting the activation of the ERK and PI3K/AKT signaling pathways in osteoarthritis chondrocytes. Exp. Med. 2021, 21, 388. [Google Scholar] [CrossRef]
- Wang, M.H.; Li, L.Z.; Sun, J.B.; Wu, F.H.; Liang, J.Y. A new antioxidant flavone glycoside from Scutellaria baicalensis Georgi. Nat. Prod. Res. 2014, 28, 1772–1776. [Google Scholar] [CrossRef]
- Han, Q.; Wang, H.; Xiao, C.; Fu, B.D.; Du, C.T. Oroxylin A inhibits H(2)O(2)-induced oxidative stress in PC12 cells. Nat. Prod. Res. 2017, 31, 1339–1342. [Google Scholar] [CrossRef] [PubMed]
- Li, H.N.; Nie, F.F.; Liu, W.; Dai, Q.S.; Lu, N.; Qi, Q.; Li, Z.Y.; You, Q.D.; Guo, Q.L. Apoptosis induction of oroxylin A in human cervical cancer HeLa cell line in vitro and in vivo. Toxicology 2009, 257, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhou, Y.; Qiao, C.; Ni, T.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Oroxylin A inhibits glycolysis-dependent proliferation of human breast cancer via promoting SIRT3-mediated SOD2 transcription and HIF1alpha destabilization. Cell Death Dis. 2015, 6, e1714. [Google Scholar] [CrossRef] [Green Version]
- Kai, J.; Yang, X.; Wang, Z.; Wang, F.; Jia, Y.; Wang, S.; Tan, S.; Chen, A.; Shao, J.; Zhang, F.; et al. Oroxylin a promotes PGC-1α/Mfn2 signaling to attenuate hepatocyte pyroptosis via blocking mitochondrial ROS in alcoholic liver disease. Free Radic. Biol. Med. 2020, 153, 89–102. [Google Scholar] [CrossRef]
- Huang, X.E.; Wei, D.; Yang, Y.N.; Chen, S.Q.; Zhu, M.; Zhang, X.M.; Yu, J. MPTP related mitochondrial pathway in oroxylin A induced-apoptosis in HepG2 cancer cells. Int. J. Clin. Exp. Pathol. 2016, 9, 11139–11148. [Google Scholar]
- Zhang, Z.; Guo, M.; Shen, M.; Li, Y.; Tan, S.; Shao, J.; Zhang, F.; Chen, A.; Wang, S.; Zheng, S. Oroxylin A regulates the turnover of lipid droplet via downregulating adipose triglyceride lipase (ATGL) in hepatic stellate cells. Life Sci. 2019, 238, 116934. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y.; Zhu, W.; Ji, M.; Xu, J.; Guo, Y.; Gao, F.; Gu, W.; Yang, X.; Zhang, C. Oroxylin A inhibits Kaposi’s sarcoma-associated herpes virus (KSHV) vIL-6-mediated lymphatic reprogramming of vascular endothelial cells through modulating PPARgamma/Prox1 axis. J. Med. Virol. 2019, 91, 463–472. [Google Scholar] [CrossRef]
- Qiao, C.; Wei, L.; Dai, Q.; Zhou, Y.; Yin, Q.; Li, Z.; Xiao, Y.; Guo, Q.; Lu, N. UCP2-related mitochondrial pathway participates in oroxylin A-induced apoptosis in human colon cancer cells. J. Cell. Physiol. 2015, 230, 1054–1063. [Google Scholar] [CrossRef]
- Ni, T.; He, Z.; Dai, Y.; Yao, J.; Guo, Q.; Wei, L. Oroxylin A suppresses the development and growth of colorectal cancer through reprogram of HIF1α-modulated fatty acid metabolism. Cell Death Dis. 2017, 8, e2865. [Google Scholar] [CrossRef]
- Li, H.; Lu, N.; Yu, X.; Liu, X.; Hu, P.; Zhu, Y.; Shen, L.; Xu, J.; Li, Z.; Guo, Q.; et al. Oroxylin A, a natural compound, mitigates the negative effects of TNFalpha-treated acute myelogenous leukemia cells. Carcinogenesis 2018, 39, 1292–1303. [Google Scholar] [CrossRef]
- Liu, W.; Mu, R.; Nie, F.F.; Yang, Y.; Wang, J.; Dai, Q.S.; Lu, N.; Qi, Q.; Rong, J.J.; Hu, R.; et al. MAC-related mitochondrial pathway in oroxylin-A-induced apoptosis in human hepatocellular carcinoma HepG2 cells. Cancer Lett. 2009, 284, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, F.; Wang, Y.; Cai, M.; Wang, Q.; Guo, Q.; Li, Z.; Hu, R. Oroxylin A inhibits colitis-associated carcinogenesis through modulating the IL-6/STAT3 signaling pathway. Inflamm. Bowel. Dis. 2013, 19, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and chronic diseases: How are they linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Saghari, S.; Bassiri, F.; Raesi, R.; Zarrabi, A.; Hushmandi, K.; Sethi, G.; Tergaonkar, V. NF-κB as a regulator of cancer metastasis and therapy response: A focus on epithelial-mesenchymal transition. J. Cell. Physiol. 2022, 237, 2770–2795. [Google Scholar] [CrossRef]
- Morgan, D.; Garg, M.; Tergaonkar, V.; Tan, S.Y.; Sethi, G. Pharmacological significance of the non-canonical NF-κB pathway in tumorigenesis. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188449. [Google Scholar] [CrossRef]
- Kim, C.; Lee, S.G.; Yang, W.M.; Arfuso, F.; Um, J.Y.; Kumar, A.P.; Bian, J.; Sethi, G.; Ahn, K.S. Formononetin-induced oxidative stress abrogates the activation of STAT3/5 signaling axis and suppresses the tumor growth in multiple myeloma preclinical model. Cancer Lett. 2018, 431, 123–141. [Google Scholar] [CrossRef]
- Hiremath, I.S.; Goel, A.; Warrier, S.; Kumar, A.P.; Sethi, G.; Garg, M. The multidimensional role of the Wnt/β-catenin signaling pathway in human malignancies. J. Cell. Physiol. 2022, 237, 199–238. [Google Scholar] [CrossRef]
- Dai, X.; Ahn, K.S.; Kim, C.; Siveen, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Shi, J.; Kumar, A.P.; Wang, L.Z.; et al. Ascochlorin, an isoprenoid antibiotic inhibits growth and invasion of hepatocellular carcinoma by targeting STAT3 signaling cascade through the induction of PIAS3. Mol. Oncol. 2015, 9, 818–833. [Google Scholar] [CrossRef]
- Shen, M.; Guo, M.; Wang, Z.; Li, Y.; Kong, D.; Shao, J.; Tan, S.; Chen, A.; Zhang, F.; Zhang, Z.; et al. ROS-dependent inhibition of the PI3K/Akt/mTOR signaling is required for Oroxylin A to exert anti-inflammatory activity in liver fibrosis. Int. Immunopharmacol. 2020, 85, 106637. [Google Scholar] [CrossRef]
- Xuan, L.; Zhang, J.; Ji, J.; Hu, J.; Li, F. Oroxylin A exerts its antitumor effects in human gallbladder cancer via inhibition of the PTEN/PI3K/AKT signaling pathway. Biol. Pharm. Bull. 2020, 43, 1511–1518. [Google Scholar] [CrossRef]
- Wei, L.; Dai, Y.; Zhou, Y.; He, Z.; Yao, J.; Zhao, L.; Guo, Q.; Yang, L. Oroxylin A activates PKM1/HNF4 alpha to induce hepatoma differentiation and block cancer progression. Cell Death Dis. 2017, 8, e2944. [Google Scholar] [CrossRef]
- Yang, M.H.; Lee, J.H.; Ko, J.H.; Jung, S.H.; Sethi, G.; Ahn, K.S. Brassinin represses invasive potential of lung carcinoma cells through deactivation of PI3K/Akt/mTOR signaling cascade. Molecules 2019, 24, 1584. [Google Scholar] [CrossRef]
- Ong, P.S.; Wang, L.Z.; Dai, X.; Tseng, S.H.; Loo, S.J.; Sethi, G. Judicious toggling of mTOR activity to combat insulin resistance and cancer: Current evidence and perspectives. Front. Pharm. 2016, 7, 395. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef]
- Siegel, R.; DeSantis, C.; Virgo, K.; Stein, K.; Mariotto, A.; Smith, T.; Cooper, D.; Gansler, T.; Lerro, C.; Fedewa, S. Cancer treatment and survivorship statistics, 2012. CA A Cancer J. Clin. 2012, 62, 220–241. [Google Scholar] [CrossRef]
- Sleire, L.; Førde, H.E.; Netland, I.A.; Leiss, L.; Skeie, B.S.; Enger, P.Ø. Drug repurposing in cancer. Pharmacol. Res. 2017, 124, 74–91. [Google Scholar] [CrossRef]
- Mondal, M.; Guo, J.; He, P.; Zhou, D. Recent advances of oncolytic virus in cancer therapy. Hum. Vaccin. Immunother. 2020, 16, 2389–2402. [Google Scholar] [CrossRef]
- Roy, N.K.; Bordoloi, D.; Monisha, J.; Padmavathi, G.; Kotoky, J.; Golla, R.; Kunnumakkara, A.B. Specific targeting of Akt kinase isoforms: Taking the precise path for prevention and treatment of cancer. Curr. Drug Targets 2017, 18, 421–435. [Google Scholar] [CrossRef]
- Monisha, J.; Padmavathi, G.; Roy, N.K.; Deka, A.; Bordoloi, D.; Anip, A.; Kunnumakkara, A.B. NF-κB blockers gifted by mother nature: Prospectives in cancer cell chemosensitization. Curr. Pharm. Des. 2016, 22, 4173–4200. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Rana, V.; Parama, D.; Banik, K.; Girisa, S.; Henamayee, S.; Thakur, K.K.; Dutta, U.; Garodia, P.; Gupta, S.C.; et al. COVID-19, cytokines, inflammation, and spices: How are they related? Life Sci. 2021, 284, 119201. [Google Scholar] [CrossRef]
- Sethi, G.; Ahn, K.S.; Sung, B.; Kunnumakkara, A.B.; Chaturvedi, M.M.; Aggarwal, B.B. SH-5, an AKT inhibitor potentiates apoptosis and inhibits invasion through the suppression of anti-apoptotic, proliferative and metastatic gene products regulated by IκBα kinase activation. Biochem. Pharmacol. 2008, 76, 1404–1416. [Google Scholar] [CrossRef]
- Goyal, B.; Yadav, S.R.M.; Awasthee, N.; Gupta, S.; Kunnumakkara, A.B.; Gupta, S.C. Diagnostic, prognostic, and therapeutic significance of long non-coding RNA MALAT1 in cancer. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2021, 1875, 188502. [Google Scholar] [CrossRef]
- Khatoon, E.; Banik, K.; Harsha, C.; Sailo, B.L.; Thakur, K.K.; Khwairakpam, A.D.; Vikkurthi, R.; Devi, T.B.; Gupta, S.C.; Kunnumakkara, A.B. Phytochemicals in cancer cell chemosensitization: Current knowledge and future perspectives. Semin. Cancer Biol. 2022, 80, 306–339. [Google Scholar] [CrossRef]
- Maruthanila, V.L.; Elancheran, R.; Kunnumakkara, A.B.; Kabilan, S.; Kotoky, J. Recent development of targeted approaches for the treatment of breast cancer. Breast Cancer 2017, 24, 191–219. [Google Scholar] [CrossRef]
- Thakur, K.K.; Bordoloi, D.; Kunnumakkara, A.B. Alarming burden of triple-negative breast cancer in India. Clin. Breast Cancer 2018, 18, e393–e399. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Golani, A.; Kumar, L.D. EMT in breast cancer metastasis: An interplay of microRNAs, signaling pathways and circulating tumor cells. Front. Biosci. 2020, 25, 979–1010. [Google Scholar] [CrossRef] [PubMed]
- Thakur, K.K.; Kumar, A.; Banik, K.; Verma, E.; Khatoon, E.; Harsha, C.; Sethi, G.; Gupta, S.C.; Kunnumakkara, A.B. Long noncoding RNAs in triple-negative breast cancer: A new frontier in the regulation of tumorigenesis. J. Cell. Physiol. 2021, 236, 7938–7965. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.Y.; Shanmugam, M.K.; Sethi, G.; Bishayee, A. Potential role of targeted therapies in the treatment of triple-negative breast cancer. Anticancer Drugs 2016, 27, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Ahn, K.S.; Hsu, A.; Woo, C.C.; Yuan, Y.; Tan, K.H.B.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Koh, A.P.F.; et al. Thymoquinone inhibits bone metastasis of breast cancer cells through abrogation of the CXCR4 signaling axis. Front. Pharm. 2018, 9, 1294. [Google Scholar] [CrossRef]
- Sun, X.; Chang, X.; Wang, Y.; Xu, B.; Cao, X. Oroxylin A suppresses the cell proliferation, migration, and EMT via NF-kappaB signaling pathway in human breast cancer cells. Biomed Res. Int. 2019, 2019, 9241769. [Google Scholar] [CrossRef]
- Katoch, A.; Nayak, D.; Faheem, M.M.; Kumar, A.; Sahu, P.K.; Gupta, A.P.; Kumar, L.D.; Goswami, A. Natural podophyllotoxin analog 4DPG attenuates EMT and colorectal cancer progression via activation of checkpoint kinase 2. Cell Death Discov. 2021, 7, 25. [Google Scholar] [CrossRef]
- Chakraborty, S.; Kumar, A.; Faheem, M.M.; Katoch, A.; Kumar, A.; Jamwal, V.L.; Nayak, D.; Golani, A.; Rasool, R.U.; Ahmad, S.M.; et al. Vimentin activation in early apoptotic cancer cells errands survival pathways during DNA damage inducer CPT treatment in colon carcinoma model. Cell Death Dis. 2019, 10, 467. [Google Scholar] [CrossRef]
- Qi, L.; Ding, Y. Screening of differentiation-specific molecular biomarkers for colon cancer. Cell Physiol. Biochem. 2018, 46, 2543–2550. [Google Scholar] [CrossRef]
- Ha, J.; Zhao, L.; Zhao, Q.; Yao, J.; Zhu, B.B.; Lu, N.; Ke, X.; Yang, H.Y.; Li, Z.; You, Q.D.; et al. Oroxylin A improves the sensitivity of HT-29 human colon cancer cells to 5-FU through modulation of the COX-2 signaling pathway. Biochem. Cell Biol. 2012, 90, 521–531. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, Z.; Wang, J.; Yang, L.; Zhao, Q.; Wang, J.; Qi, Q.; Mu, R.; You, Q.D.; Guo, Q.L. Synergistic effect of 5-fluorouracil and the flavanoid oroxylin A on HepG2 human hepatocellular carcinoma and on H22 transplanted mice. Cancer Chemother. Pharm. 2010, 65, 481–489. [Google Scholar] [CrossRef]
- Huo, T.X.; Wang, X.P.; Yu, Z.; Kong, B.; He, Y.; Guo, Q.L.; Zhang, X.B.; Qiang, L. Oroxylin A inhibits the migration of hepatocellular carcinoma cells by inducing NAG-1 expression. Acta Pharm. Sin. 2022, 43, 724–734. [Google Scholar] [CrossRef]
- Weller, M.; Wick, W.; Aldape, K.; Brada, M.; Berger, M.; Pfister, S.M.; Nishikawa, R.; Rosenthal, M.; Wen, P.Y.; Stupp, R. Glioma. Nat. Rev. Dis. Primers 2015, 1, 15017. [Google Scholar] [CrossRef]
- Chen, R.; Smith-Cohn, M.; Cohen, A.L.; Colman, H. Glioma subclassifications and their clinical significance. Neurotherapeutics 2017, 14, 284–297. [Google Scholar] [CrossRef]
- Bradshaw, A.; Wickremsekera, A.; Tan, S.T.; Peng, L.; Davis, P.F.; Itinteang, T. Cancer stem cell hierarchy in glioblastoma multiforme. Front. Surg. 2016, 3, 21. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Gamit, N.; Patil, M.; Arfuso, F.; Sethi, G.; Dharmarajan, A.; Kumar, A.P.; Warrier, S. Stemness, pluripotentiality, and Wnt antagonism: sFRP4, a Wnt antagonist mediates pluripotency and stemness in glioblastoma. Cancers 2018, 11, 25. [Google Scholar] [CrossRef]
- Sun, T.; Xu, Y.J.; Jiang, S.Y.; Xu, Z.; Cao, B.Y.; Sethi, G.; Zeng, Y.Y.; Kong, Y.; Mao, X.L. Suppression of the USP10/CCND1 axis induces glioblastoma cell apoptosis. Acta Pharm. Sin. 2021, 42, 1338–1346. [Google Scholar] [CrossRef]
- Zou, M.; Hu, C.; You, Q.; Zhang, A.; Wang, X.; Guo, Q. Oroxylin A induces autophagy in human malignant glioma cells via the mTOR-STAT3-Notch signaling pathway. Mol. Carcinog. 2015, 54, 1363–1375. [Google Scholar] [CrossRef]
- Arora, L.; Kumar, A.P.; Arfuso, F.; Chng, W.J.; Sethi, G. The role of signal transducer and activator of transcription 3 (STAT3) and its targeted inhibition in hematological malignancies. Cancers 2018, 10, 327. [Google Scholar] [CrossRef] [Green Version]
- Kirtonia, A.; Pandya, G.; Sethi, G.; Pandey, A.K.; Das, B.C.; Garg, M. A comprehensive review of genetic alterations and molecular targeted therapies for the implementation of personalized medicine in acute myeloid leukemia. J. Mol. Med. 2020, 98, 1069–1091. [Google Scholar] [CrossRef] [PubMed]
- Hui, H.; Zhang, X.; Li, H.; Liu, X.; Shen, L.; Zhu, Y.; Xu, J.; Guo, Q.; Lu, N. Oroxylin A, a natural anticancer flavonoid compound, induces differentiation of t(8;21)-positive Kasumi-1 and primary acute myeloid leukemia cells. J. Cancer Res. Clin. Oncol. 2016, 142, 1449–1459. [Google Scholar] [CrossRef]
- Li, W.; Ding, Q.; Ding, Y.; Lu, L.; Wang, X.; Zhang, Y.; Zhang, X.; Guo, Q.; Zhao, L. Oroxylin A reverses the drug resistance of chronic myelogenous leukemia cells to imatinib through CXCL12/CXCR7 axis in bone marrow microenvironment. Mol. Carcinog. 2017, 56, 863–876. [Google Scholar] [CrossRef]
- Wang, L.; Syn, N.L.; Subhash, V.V.; Any, Y.; Thuya, W.L.; Cheow, E.S.H.; Kong, L.; Yu, F.; Peethala, P.C.; Wong, A.L.; et al. Pan-HDAC inhibition by panobinostat mediates chemosensitization to carboplatin in non-small cell lung cancer via attenuation of EGFR signaling. Cancer Lett. 2018, 417, 152–160. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Shanmugam, M.K.; Narula, A.S.; Kim, C.; Lee, J.H.; Namjoshi, O.A.; Blough, B.E.; Sethi, G.; Ahn, K.S. Oxymatrine attenuates tumor growth and deactivates STAT5 signaling in a lung cancer xenograft model. Cancers 2019, 11, 49. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Lee, S.G.; Sethi, G.; Ahn, K.S. Ophiopogonin D, a steroidal glycoside abrogates STAT3 signaling cascade and exhibits anti-cancer activity by causing GSH/GSSG imbalance in lung carcinoma. Cancers 2018, 10, 427. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, L.L.; Li, H.; Liu, X.; Yu, X.X.; Hu, P.; Hui, H.; Guo, Q.L.; Zhang, S. Oroxylin A inhibits the generation of Tregs in non-small cell lung cancer. Oncotarget 2017, 8, 49395–49408. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Li, W.; Xu, Y.; Zhuo, Y.; Li, M.; He, Y.; Wang, X.; Guo, Q.; Zhao, L.; et al. Oroxylin A reverses hypoxia-induced cisplatin resistance through inhibiting HIF-1alpha mediated XPC transcription. Oncogene 2020, 39, 6893–6905. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Qian, X.; Ge, Y.; Yang, B.; Wang, F.; Guan, Z.; Cai, J. Oroxylin a could be a promising radiosensitizer for esophageal squamous cell carcinoma by inducing G2/M arrest and activating apoptosis. Pathol. Oncol. Res. 2017, 23, 323–328. [Google Scholar] [CrossRef]
- Huang, H.; Cai, H.; Zhang, L.; Hua, Z.; Shi, J.; Wei, Y. Oroxylin A inhibits carcinogen-induced skin tumorigenesis through inhibition of inflammation by regulating SHCBP1 in mice. Int. Immunopharmacol. 2020, 80, 106123. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Zhao, D. Epidemiological features of cardiovascular disease in Asia. JACC Asia 2021, 1, 1–13. [Google Scholar] [CrossRef]
- Jagannathan, R.; Patel, S.A.; Ali, M.K.; Narayan, K. Global updates on cardiovascular disease mortality trends and attribution of traditional risk factors. Curr. Diabetes Rep. 2019, 19, 44. [Google Scholar] [CrossRef]
- Octavia, Y.; Tocchetti, C.G.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-induced cardiomyopathy: From molecular mechanisms to therapeutic strategies. J. Mol. Cell. Cardiol. 2012, 52, 1213–1225. [Google Scholar] [CrossRef]
- Takemura, G.; Fujiwara, H. Doxorubicin-induced cardiomyopathy from the cardiotoxic mechanisms to management. Prog. Cardiovasc. Dis. 2007, 49, 330–352. [Google Scholar] [CrossRef]
- Zhang, W.-B.; Zheng, Y.-F.; Wu, Y.-G. Protective effects of oroxylin a against doxorubicin-induced cardiotoxicity via the activation of Sirt1 in mice. Oxidative Med. Cell. Longev. 2021, 2021, 6610543. [Google Scholar] [CrossRef]
- Mohammad, S.; Thiemermann, C. Role of Metabolic Endotoxemia in Systemic Inflammation and Potential Interventions. Front. Immunol. 2020, 11, 594150. [Google Scholar] [CrossRef]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Kitabatake, H.; Tanaka, N.; Fujimori, N.; Komatsu, M.; Okubo, A.; Kakegawa, K.; Kimura, T.; Sugiura, A.; Yamazaki, T.; Shibata, S.; et al. Association between endotoxemia and histological features of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 712–722. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Chaturvedi, M.M.; Aggarwal, B.B. Simvastatin, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor, suppresses osteoclastogenesis induced by receptor activator of nuclear factor-kappaB ligand through modulation of NF-kappaB pathway. Int. J. Cancer 2008, 123, 1733–1740. [Google Scholar] [CrossRef]
- Glaros, T.G.; Chang, S.; Gilliam, E.A.; Maitra, U.; Deng, H.; Li, L. Causes and consequences of low grade endotoxemia and inflammatory diseases. Front. Biosci. -Sch. 2013, 5, 754–765. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M.; Kibbe, M.R. Improving Lower Extremity Functioning in Peripheral Artery Disease: Exercise, Endovascular Revascularization, or Both? JAMA 2017, 317, 689–690. [Google Scholar] [CrossRef]
- Cooke, J.P.; Losordo, D.W. Modulating the vascular response to limb ischemia: Angiogenic and cell therapies. Circ. Res. 2015, 116, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Lu, H.; Liang, W.; Garcia-Barrio, M.T.; Guo, Y.; Zhang, J.; Zhu, T.; Hao, Y.; Zhang, J.; Chen, Y.E. Endothelial TFEB (Transcription Factor EB) Positively Regulates Postischemic Angiogenesis. Circ. Res. 2018, 122, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, L.; Li, C.; Shi, H.; Wang, Q.; Yang, W.; Fang, L.; Leng, Y.; Sun, W.; Li, M.; et al. Oroxylin a attenuates limb ischemia by promoting angiogenesis via modulation of endothelial cell migration. Front. Pharm. 2021, 12, 705617. [Google Scholar] [CrossRef] [PubMed]
- Pinzani, M.; Rombouts, K. Liver fibrosis: From the bench to clinical targets. Dig. Liver Dis. 2004, 36, 231–242. [Google Scholar] [CrossRef]
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary epidemiology of chronic liver disease and cirrhosis. Clin. Gastroenterol. Hepatol. 2020, 18, 2650–2666. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef]
- Singh, Y.P.; Girisa, S.; Banik, K.; Ghosh, S.; Swathi, P.; Deka, M.; Padmavathi, G.; Kotoky, J.; Sethi, G.; Fan, L. Potential application of zerumbone in the prevention and therapy of chronic human diseases. J. Funct. Foods 2019, 53, 248–258. [Google Scholar] [CrossRef]
- Shu, Y.; Liu, X.; Huang, H.; Wen, Q.; Shu, J. Research progress of natural compounds in anti-liver fibrosis by affecting autophagy of hepatic stellate cells. Mol. Biol. Rep. 2021, 48, 1915–1924. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, X.; Li, J. Protective effect of oroxylin A against lipopolysaccharide and/or D-galactosamine-induced acute liver injury in mice. J. Surg. Res. 2015, 195, 522–528. [Google Scholar] [CrossRef]
- Bonder, A.; Tapper, E.B.; Afdhal, N.H. Contemporary assessment of hepatic fibrosis. Clin. Liver Dis. 2015, 19, 123–134. [Google Scholar] [CrossRef]
- Friedman, S.L. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Z.; Yao, Z.; Wang, L.; Zhang, F.; Shao, J.; Chen, A.; Zheng, S. Activation of autophagy is required for Oroxylin A to alleviate carbon tetrachloride-induced liver fibrosis and hepatic stellate cell activation. Int. Immunopharmacol. 2018, 56, 148–155. [Google Scholar] [CrossRef]
- Wang, F.; Jia, Y.; Li, M.; Wang, L.; Shao, J.; Guo, Q.; Tan, S.; Ding, H.; Chen, A.; Zhang, F.; et al. Blockade of glycolysis-dependent contraction by oroxylin a via inhibition of lactate dehydrogenase-a in hepatic stellate cells. Cell Commun. Signal. 2019, 17, 11. [Google Scholar] [CrossRef]
- Gandhi, C.R. Hepatic stellate cell activation and pro-fibrogenic signals. J. Hepatol. 2017, 67, 1104–1105. [Google Scholar] [CrossRef] [PubMed]
- Bian, M.; He, J.; Jin, H.; Lian, N.; Shao, J.; Guo, Q.; Wang, S.; Zhang, F.; Zheng, S. Oroxylin A induces apoptosis of activated hepatic stellate cells through endoplasmic reticulum stress. Apoptosis 2019, 24, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Pessione, F.; Ramond, M.J.; Peters, L.; Pham, B.N.; Batel, P.; Rueff, B.; Valla, D.C. Five-year survival predictive factors in patients with excessive alcohol intake and cirrhosis. Effect of alcoholic hepatitis, smoking and abstinence. Liver Int. 2003, 23, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lian, N.; Bian, M.; Zhang, C.; Chen, X.; Shao, J.; Wu, L.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A inhibits ethanol-induced hepatocyte senescence via YAP pathway. Cell Prolif. 2018, 51, e12431. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Lee, Y.; Gullen, E.A.; Cheng, Y.C. Impacts of baicalein analogs with modification of the 6th position of A ring on the activity toward NF-kappaB-, AP-1-, or CREB-mediated transcription. Bioorg. Med. Chem. Lett. 2008, 18, 5046–5049. [Google Scholar] [CrossRef]
- Parisi, V.; Oddone, F.; Ziccardi, L.; Roberti, G.; Coppola, G.; Manni, G. Citicoline and retinal ganglion cells: Effects on morphology and function. Curr. Neuropharmacol. 2018, 16, 919–932. [Google Scholar] [CrossRef]
- Shindler, K.S.; Ventura, E.; Dutt, M.; Rostami, A. Inflammatory demyelination induces axonal injury and retinal ganglion cell apoptosis in experimental optic neuritis. Exp. Eye Res. 2008, 87, 208–213. [Google Scholar] [CrossRef]
- Potilinski, M.C.; Lorenc, V.; Perisset, S.; Gallo, J.E. Mechanisms behind retinal ganglion cell loss in diabetes and therapeutic approach. Int. J. Mol. Sci. 2020, 21, 2351. [Google Scholar] [CrossRef]
- Levin, L.A.; Louhab, A. Apoptosis of retinal ganglion cells in anterior ischemic optic neuropathy. Arch. Ophthalmol. 1996, 114, 488–491. [Google Scholar] [CrossRef]
- Quigley, H.A. Ganglion cell death in glaucoma: Pathology recapitulates ontogeny. Aust. N. Z. J. Ophthalmol. 1995, 23, 85–91. [Google Scholar] [CrossRef]
- Lin, S.F.; Chien, J.Y.; Kapupara, K.; Huang, C.F.; Huang, S.P. Oroxylin A promotes retinal ganglion cell survival in a rat optic nerve crush model. PLoS ONE 2017, 12, e0178584. [Google Scholar] [CrossRef]
- Biousse, V.; Newman, N.J. Ischemic optic neuropathies. N. Engl. J. Med. 2015, 372, 2428–2436. [Google Scholar] [CrossRef]
- Luneau, K.; Newman, N.J.; Biousse, V. Ischemic optic neuropathies. Neurologist 2008, 14, 341–354. [Google Scholar] [CrossRef]
- Chien, J.Y.; Lin, S.F.; Chou, Y.Y.; Huang, C.F.; Huang, S.P. Protective effects of Oroxylin A on retinal ganglion cells in experimental model of anterior ischemic optic neuropathy. Antioxidants 2021, 10, 902. [Google Scholar] [CrossRef]
- Hanania, N.A. Targeting airway inflammation in asthma: Current and future therapies. Chest 2008, 133, 989–998. [Google Scholar] [CrossRef]
- Goh, F.Y.; Upton, N.; Guan, S.; Cheng, C.; Shanmugam, M.K.; Sethi, G.; Leung, B.P.; Wong, W.S. Fisetin, a bioactive flavonol, attenuates allergic airway inflammation through negative regulation of NF-κB. Eur. J. Pharm. 2012, 679, 109–116. [Google Scholar] [CrossRef]
- Doeing, D.C.; Solway, J. Airway smooth muscle in the pathophysiology and treatment of asthma. J. Appl. Physiol. 2013, 114, 834–843. [Google Scholar] [CrossRef]
- Zhou, D.G.; Diao, B.Z.; Zhou, W.; Feng, J.L. Oroxylin A inhibits allergic airway inflammation in ovalbumin (OVA)-induced asthma murine model. Inflammation 2016, 39, 867–872. [Google Scholar] [CrossRef]
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef]
- Ponder, A.; Long, M.D. A clinical review of recent findings in the epidemiology of inflammatory bowel disease. Clin. Epidemiol. 2013, 5, 237–247. [Google Scholar] [CrossRef]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef]
- Biasi, F.; Leonarduzzi, G.; Oteiza, P.I.; Poli, G. Inflammatory bowel disease: Mechanisms, redox considerations, and therapeutic targets. Antioxid. Redox Signal. 2013, 19, 1711–1747. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.H.; Lamkanfi, M.; Kanneganti, T.D. The Nlrp3 inflammasome: Contributions to intestinal homeostasis. Trends Immunol. 2011, 32, 171–179. [Google Scholar] [CrossRef]
- Bai, D.; Sun, T.; Zhao, J.; Du, J.; Bu, X.; Cao, W.; Zhao, Y.; Lu, N. Oroxylin A maintains the colonic mucus barrier to reduce disease susceptibility by reconstituting a dietary fiber-deprived gut microbiota. Cancer Lett. 2021, 515, 73–85. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, X.; Zhang, X.; Tang, J.; Li, Z.; Wang, Q.; Hu, R. Oroxylin A inhibits colitis by inactivating NLRP3 inflammasome. Oncotarget 2017, 8, 58903–58917. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Y.; Zhao, Y.; Ding, Y.; Zhang, X.; Kong, L.; Li, Z.; Guo, Q.; Zhao, L. Oroxyloside prevents dextran sulfate sodium-induced experimental colitis in mice by inhibiting NF-κB pathway through PPARγ activation. Biochem. Pharmacol. 2016, 106, 70–81. [Google Scholar] [CrossRef]
- Dahaghin, S.; Bierma-Zeinstra, S.M.; Ginai, A.Z.; Pols, H.A.; Hazes, J.M.; Koes, B.W. Prevalence and pattern of radiographic hand osteoarthritis and association with pain and disability (the Rotterdam study). Ann. Rheum. Dis. 2005, 64, 682–687. [Google Scholar] [CrossRef]
- Oliveria, S.A.; Felson, D.T.; Reed, J.I.; Cirillo, P.A.; Walker, A.M. Incidence of symptomatic hand, hip, and knee osteoarthritis among patients in a health maintenance organization. Arthritis Rheum 1995, 38, 1134–1141. [Google Scholar] [CrossRef]
- Felson, D.T. Epidemiology of hip and knee osteoarthritis. Epidemiol. Rev. 1988, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.H.; Zheng, G.; Zhong, X.Y.; Lin, Z.H.; Yang, S.W.; Liu, H.X.; Shang, P. Oroxylin A attenuates osteoarthritis progression by dual inhibition of cell inflammation and hypertrophy. Food Funct. 2021, 12, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Ferro, F.; Elefante, E.; Luciano, N.; Talarico, R.; Todoerti, M. One year in review 2017: Novelties in the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 721–734. [Google Scholar]
- Bortoluzzi, A.; Furini, F.; Generali, E.; Silvagni, E.; Luciano, N.; Scirè, C.A. One year in review 2018: Novelties in the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2018, 36, 347–361. [Google Scholar]
- Xu, T.; Liu, S.; Zhao, J.; Feng, G.; Pi, Z.; Song, F.; Liu, Z. A study on the effective substance of the Wu-tou formula based on the metabonomic method using UPLC-Q-TOF-HDMS. Mol. Biosyst. 2015, 11, 3081–3091. [Google Scholar] [CrossRef]
- Jacob, J.; Amalraj, A.; Raj, K.K.J.; Divya, C.; Kunnumakkara, A.B.; Gopi, S. A novel bioavailable hydrogenated curcuminoids formulation (CuroWhite™) improves symptoms and diagnostic indicators in rheumatoid arthritis patients-A randomized, double blind and placebo controlled study. J. Tradit. Complementary Med. 2019, 9, 346–352. [Google Scholar] [CrossRef]
- Wang, Y.L.; Gao, J.M.; Xing, L.Z. Therapeutic potential of Oroxylin A in rheumatoid arthritis. Int. Immunopharmacol. 2016, 40, 294–299. [Google Scholar] [CrossRef]
- Polanczyk, G.; de Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The worldwide prevalence of ADHD: A systematic review and metaregression analysis. Am. J. Psychiatry 2007, 164, 942–948. [Google Scholar] [CrossRef]
- Faraone, S.V.; Asherson, P.; Banaschewski, T.; Biederman, J.; Buitelaar, J.K.; Ramos-Quiroga, J.A.; Rohde, L.A.; Sonuga-Barke, E.J.; Tannock, R.; Franke, B. Attention-deficit/hyperactivity disorder. Nat. Rev. Dis. Primers 2015, 1, 15020. [Google Scholar] [CrossRef]
- Thapar, A.; Van den Bree, M.; Fowler, T.; Langley, K.; Whittinger, N. Predictors of antisocial behaviour in children with attention deficit hyperactivity disorder. Eur. Child Adolesc. Psychiatry 2006, 15, 118–125. [Google Scholar] [CrossRef]
- Van den Berg, A.E.; Van den Berg, C. A comparison of children with ADHD in a natural and built setting. Child Care Health Dev. 2011, 37, 430–439. [Google Scholar] [CrossRef]
- Schachter, H.M.; Pham, B.; King, J.; Langford, S.; Moher, D. How efficacious and safe is short-acting methylphenidate for the treatment of attention-deficit disorder in children and adolescents? A meta-analysis. Can. Med. Assoc. J. 2001, 165, 1475–1488. [Google Scholar]
- Wilens, T.E.; Kwon, A.; Tanguay, S.; Chase, R.; Moore, H.; Faraone, S.V.; Biederman, J. Characteristics of adults with attention deficit hyperactivity disorder plus substance use disorder: The role of psychiatric comorbidity. Am. J. Addict. 2005, 14, 319–327. [Google Scholar] [CrossRef]
- Yoon, S.Y.; dela Peña, I.; Kim, S.M.; Woo, T.S.; Shin, C.Y.; Son, K.H.; Park, H.; Lee, Y.S.; Ryu, J.H.; Jin, M.; et al. Oroxylin A improves attention deficit hyperactivity disorder-like behaviors in the spontaneously hypertensive rat and inhibits reuptake of dopamine in vitro. Arch. Pharm. Res. 2013, 36, 134–140. [Google Scholar] [CrossRef]
- Selkoe, D.J. The origins of Alzheimer disease: A is for amyloid. Jama 2000, 283, 1615–1617. [Google Scholar] [CrossRef]
- Segaran, R.C.; Chan, L.Y.; Wang, H.; Sethi, G.; Tang, F.R. Neuronal development-related miRNAs as biomarkers for Alzheimer’s disease, depression, schizophrenia and ionizing radiation exposure. Curr. Med. Chem. 2021, 28, 19–52. [Google Scholar] [CrossRef]
- Chai, Y.L.; Chong, J.R.; Raquib, A.R.; Xu, X.; Hilal, S.; Venketasubramanian, N.; Tan, B.Y.; Kumar, A.P.; Sethi, G.; Chen, C.P.; et al. Plasma osteopontin as a biomarker of Alzheimer’s disease and vascular cognitive impairment. Sci. Rep. 2021, 11, 4010. [Google Scholar] [CrossRef]
- Kukull, W.A.; Bowen, J.D. Dementia epidemiology. Med. Clin. N. Am. 2002, 86, 573–590. [Google Scholar] [CrossRef]
- Teng, E.; Lu, P.H.; Cummings, J.L. Neuropsychiatric symptoms are associated with progression from mild cognitive impairment to Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2007, 24, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Doody, R.S.; Pavlik, V.; Massman, P.; Rountree, S.; Darby, E.; Chan, W. Predicting progression of Alzheimer’s disease. Alzheimer’s Res. Ther. 2010, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Ahadieh, S.; Corrigan, B.; French, J.; Fullerton, T.; Tensfeldt, T.; Group, A.s.D.W. Disease progression meta-analysis model in Alzheimer’s disease. Alzheimer’s Dement. 2010, 6, 39–53. [Google Scholar] [CrossRef] [PubMed]
- DeKosky, S.T.; Marek, K. Looking backward to move forward: Early detection of neurodegenerative disorders. Science 2003, 302, 830–834. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.; Jeon, S.J.; Son, K.H.; Lee, S.; Yoon, B.H.; Cheong, J.H.; Ko, K.H.; Ryu, J.H. The effects of acute and repeated oroxylin A treatments on Abeta(25-35)-induced memory impairment in mice. Neuropharmacology 2008, 55, 639–647. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeon, S.J.; Son, K.H.; Jung, J.W.; Lee, S.; Yoon, B.H.; Lee, J.J.; Cho, Y.W.; Cheong, J.H.; Ko, K.H.; et al. The ameliorating effect of oroxylin A on scopolamine-induced memory impairment in mice. Neurobiol. Learn. Mem. 2007, 87, 536–546. [Google Scholar] [CrossRef]
- Nuttall, F.Q. Body Mass Index: Obesity, BMI, and Health: A Critical Review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Skelton, J.A.; Irby, M.B.; Grzywacz, J.G.; Miller, G. Etiologies of obesity in children: Nature and nurture. Pediatr. Clin. N. Am. 2011, 58, 1333–1354. [Google Scholar] [CrossRef]
- McAllister, E.J.; Dhurandhar, N.V.; Keith, S.W.; Aronne, L.J.; Barger, J.; Baskin, M.; Benca, R.M.; Biggio, J.; Boggiano, M.M.; Eisenmann, J.C.; et al. Ten putative contributors to the obesity epidemic. Crit. Rev. Food Sci. Nutr. 2009, 49, 868–913. [Google Scholar] [CrossRef]
- Calle, E.E.; Kaaks, R. Overweight, obesity and cancer: Epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef]
- Skelton, J.A.; DeMattia, L.; Miller, L.; Olivier, M. Obesity and its therapy: From genes to community action. Pediatr. Clin. N. Am. 2006, 53, 777–794. [Google Scholar] [CrossRef]
- Mangal, P.; Khare, P.; Jagtap, S.; Bishnoi, M.; Kondepudi, K.K.; Bhutani, K.K. Screening of six Ayurvedic medicinal plants for anti-obesity potential: An investigation on bioactive constituents from Oroxylum indicum (L.) Kurz bark. J. Ethnopharmacol. 2017, 197, 138–146. [Google Scholar] [CrossRef]
- Tebruegge, M.; Curtis, N. Enterovirus infections in neonates. Semin. Fetal. Neonatal. Med. 2009, 14, 222–227. [Google Scholar] [CrossRef]
- Garmaroudi, F.S.; Marchant, D.; Hendry, R.; Luo, H.; Yang, D.; Ye, X.; Shi, J.; McManus, B.M. Coxsackievirus B3 replication and pathogenesis. Future Microbiol. 2015, 10, 629–653. [Google Scholar] [CrossRef]
- Kwon, B.E.; Song, J.H.; Song, H.H.; Kang, J.W.; Hwang, S.N.; Rhee, K.J.; Shim, A.; Hong, E.H.; Kim, Y.J.; Jeon, S.M.; et al. Antiviral activity of Oroxylin A against Coxsackievirus B3 alleviates virus-induced acute pancreatic damage in mice. PLoS ONE 2016, 11, e0155784. [Google Scholar] [CrossRef]
- Xian, Y.; Su, Y.; Liang, J.; Long, F.; Feng, X.; Xiao, Y.; Lian, H.; Xu, J.; Zhao, J.; Liu, Q.; et al. Oroxylin A reduces osteoclast formation and bone resorption via suppressing RANKL-induced ROS and NFATc1 activation. Biochem. Pharm. 2021, 193, 114761. [Google Scholar] [CrossRef]
- Ren, G.; Chen, H.; Zhang, M.; Yang, N.; Yang, H.; Xu, C.; Li, J.; Ning, C.; Song, Z.; Zhou, S.; et al. Pharmacokinetics, tissue distribution and excretion study of Oroxylin A, Oroxylin A 7-O-glucuronide and Oroxylin A sodium sulfonate in rats after administration of Oroxylin A. Fitoterapia 2020, 142, 104480. [Google Scholar] [CrossRef]
- Ren, G.; Chen, H.; Zhang, M.; Yang, N.; Yang, H.; Xu, C.; Li, J.; Ning, C.; Song, Z.; Zhou, S.; et al. Determination of oroxylin A, oroxylin A 7-O-glucuronide, and oroxylin A sodium sulfonate in beagle dogs by using UHPLC MS/MS Application in a pharmacokinetic study. J. Sep. Sci. 2020, 43, 2290–2300. [Google Scholar] [CrossRef]
- Fong, S.Y.; Wong, Y.C.; Zuo, Z. Development of a SPE-LC/MS/MS method for simultaneous quantification of baicalein, wogonin, oroxylin A and their glucuronides baicalin, wogonoside and oroxyloside in rats and its application to brain uptake and plasma pharmacokinetic studies. J. Pharm. Biomed. Anal. 2014, 97, 9–23. [Google Scholar] [CrossRef]
- Zhang, Q.; Cong, D.; An, D.; Fan, A.; Liu, Q.; Yi, Y.; Song, Z.; Chen, X.; Lu, Y.; Zhao, D.; et al. Determination of oroxylin A and oroxylin A 7-O-d-glucuronide in HepG2 cell lysate and subcellular fractions with SPE-UPLC-MS/MS: Cellular pharmacokinetic study to indicate anti-cancer mechanisms. J. Pharm. Biomed. Anal. 2018, 154, 364–372. [Google Scholar] [CrossRef]
- Mu, R.; Qi, Q.; Gu, H.; Wang, J.; Yang, Y.; Rong, J.; Liu, W.; Lu, N.; You, Q.; Guo, Q. Involvement of p53 in oroxylin A-induced apoptosis in cancer cells. Mol. Carcinog. 2009, 48, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Miao, H.; Zhang, Y.; Li, W.; Li, Z.; Zhou, Y.; Zhao, L.; Guo, Q. Bone marrow microenvironment confers imatinib resistance to chronic myelogenous leukemia and oroxylin A reverses the resistance by suppressing Stat3 pathway. Arch. Toxicol. 2015, 89, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Ma, R.; Huang, S.; Liao, Y.; Ding, Y.; Li, Z.; Guo, Q.; Tan, R.; Zhang, L.; Zhao, L. Oroxylin A increases the sensitivity of temozolomide on glioma cells by hypoxia-inducible factor 1α/hedgehog pathway under hypoxia. J. Cell. Physiol. 2019, 234, 17392–17404. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S.; et al. Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study. Arch. Intern. Med. 1995, 155, 381–386. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Ferri, C. Flavonoids: Antioxidants against atherosclerosis. Nutrients 2010, 2, 889–902. [Google Scholar] [CrossRef]
- Arai, Y.; Watanabe, S.; Kimira, M.; Shimoi, K.; Mochizuki, R.; Kinae, N. Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 2000, 130, 2243–2250. [Google Scholar] [CrossRef] [Green Version]
- Rana, V.; Parama, D.; Girisa, S.; Harsha, C.; Kunnumakkara, A.B. Oxidative Stress And Inflammation. In Antioxidants and Functional Foods for Neurodegenerative Disorders; CRC Press: Boca Raton, FL, USA, 2021; pp. 21–36. [Google Scholar]
- Yang, Y.; Hu, Y.; Gu, H.Y.; Lu, N.; Liu, W.; Qi, Q.; Zhao, L.; Wang, X.T.; You, Q.D.; Guo, Q.L. Oroxylin A induces G2/M phase cell-cycle arrest via inhibiting Cdk7-mediated expression of Cdc2/p34 in human gastric carcinoma BGC-823 cells. J. Pharm. Pharmacol. 2008, 60, 1459–1463. [Google Scholar] [CrossRef]
- Yao, J.; Hu, R.; Sun, J.; Lin, B.; Zhao, L.; Sha, Y.; Zhu, B.; You, Q.D.; Yan, T.; Guo, Q.L. Oroxylin A prevents inflammation-related tumor through down-regulation of inflammatory gene expression by inhibiting NF-kappaB signaling. Mol. Carcinog. 2014, 53, 145–158. [Google Scholar] [CrossRef]
- Ku, W.T.; Tung, J.J.; Lee, T.J.; Lai, K.C. Long-Term Exposure to Oroxylin A Inhibits Metastasis by Suppressing CCL2 in Oral Squamous Cell Carcinoma Cells. Cancers 2019, 11, 353. [Google Scholar] [CrossRef]
- Zhao, K.; Li, X.; Lin, B.; Yang, D.; Zhou, Y.; Li, Z.; Guo, Q.; Lu, N. Oroxyloside inhibits angiogenesis through suppressing internalization of VEGFR2/Flk-1 in endothelial cells. J. Cell Physiol. 2018, 233, 3454–3464. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, Y.; You, Q.D.; Liu, W.; Gu, H.Y.; Zhao, L.; Zhang, K.; Wang, W.; Wang, X.T.; Guo, Q.L. Oroxylin A induced apoptosis of human hepatocellular carcinoma cell line HepG2 was involved in its antitumor activity. Biochem. Biophys. Res. Commun. 2006, 351, 521–527. [Google Scholar] [CrossRef]
- Zhu, B.; Zhao, L.; Zhu, L.; Wang, H.; Sha, Y.; Yao, J.; Li, Z.; You, Q.; Guo, Q. Oroxylin A reverses CAM-DR of HepG2 cells by suppressing Integrinbeta1 and its related pathway. Toxicol. Appl. Pharmacol. 2012, 259, 387–394. [Google Scholar] [CrossRef]
- Xu, Z.F.; Sun, X.K.; Chen, G.; Han, C.; Wang, F.; Zhang, Y.D. Oroxyloside inhibits human glioma progression by suppressing proliferation, metastasis and inducing apoptosis related pathways. Biomed. Pharmacother. 2018, 97, 1564–1574. [Google Scholar] [CrossRef]
- Zhang, C.; Bian, M.; Chen, X.; Jin, H.; Zhao, S.; Yang, X.; Shao, J.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A prevents angiogenesis of LSECs in liver fibrosis via inhibition of YAP/HIF-1α signaling. J. Cell. Biochem. 2018, 119, 2258–2268. [Google Scholar] [CrossRef]
- Li, J.; Tong, D.; Liu, J.; Chen, F.; Shen, Y. Oroxylin A attenuates cigarette smoke-induced lung inflammation by activating Nrf2. Int. Immunopharmacol. 2016, 40, 524–529. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, H.; Li, W.; Yao, J.; Sun, Y.; Li, Z.; Zhao, L.; Guo, Q. CXCL12/CXCR4 axis confers adriamycin resistance to human chronic myelogenous leukemia and oroxylin A improves the sensitivity of K562/ADM cells. Biochem. Pharm. 2014, 90, 212–225. [Google Scholar] [CrossRef]
- Yang, H.Y.; Zhao, L.; Yang, Z.; Zhao, Q.; Qiang, L.; Ha, J.; Li, Z.Y.; You, Q.D.; Guo, Q.L. Oroxylin A reverses multi-drug resistance of human hepatoma BEL7402/5-FU cells via downregulation of P-glycoprotein expression by inhibiting NF-kappaB signaling pathway. Mol. Carcinog. 2012, 51, 185–195. [Google Scholar] [CrossRef]
- Li, C.; Zhang, L.; Lin, G.; Zuo, Z. Identification and quantification of baicalein, wogonin, oroxylin A and their major glucuronide conjugated metabolites in rat plasma after oral administration of Radix scutellariae product. J. Pharm. Biomed. Anal. 2011, 54, 750–758. [Google Scholar] [CrossRef]
- Cai, Y.; Li, S.; Li, T.; Zhou, R.; Wai, A.T.; Yan, R. Oral pharmacokinetics of baicalin, wogonoside, oroxylin A 7-O-β-d-glucuronide and their aglycones from an aqueous extract of Scutellariae Radix in the rat. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1026, 124–133. [Google Scholar] [CrossRef]
- Tukey, R.H.; Strassburg, C.P. Human UDP-glucuronosyltransferases: Metabolism, expression, and disease. Annu. Rev. Pharm. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef]
- Fu, W.; Wang, J.; Yu, L.; Zhao, L.; Lu, N.; You, Q.; Guo, Q.; Li, Z. Synthesis and biological evaluation of 7-O-modified oroxylin A derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 1118–1121. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sajeev, A.; Hegde, M.; Girisa, S.; Devanarayanan, T.N.; Alqahtani, M.S.; Abbas, M.; Sil, S.K.; Sethi, G.; Chen, J.-T.; Kunnumakkara, A.B. Oroxylin A: A Promising Flavonoid for Prevention and Treatment of Chronic Diseases. Biomolecules 2022, 12, 1185. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12091185
Sajeev A, Hegde M, Girisa S, Devanarayanan TN, Alqahtani MS, Abbas M, Sil SK, Sethi G, Chen J-T, Kunnumakkara AB. Oroxylin A: A Promising Flavonoid for Prevention and Treatment of Chronic Diseases. Biomolecules. 2022; 12(9):1185. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12091185
Chicago/Turabian StyleSajeev, Anjana, Mangala Hegde, Sosmitha Girisa, Thulasidharan Nair Devanarayanan, Mohammed S. Alqahtani, Mohamed Abbas, Samir Kumar Sil, Gautam Sethi, Jen-Tsung Chen, and Ajaikumar B. Kunnumakkara. 2022. "Oroxylin A: A Promising Flavonoid for Prevention and Treatment of Chronic Diseases" Biomolecules 12, no. 9: 1185. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12091185