Host Plant-Derived miRNAs Potentially Modulate the Development of a Cosmopolitan Insect Pest, Plutella xylostella

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Sampling
2.2. Plants
2.3. Small RNA Sequencing
2.4. RNA Extraction and qRT-PCR
2.5. Target Prediction and Transcriptome Sequencing
2.6. Cell Line and Luciferase Assays
2.7. miRNA Agomir Treatments
2.8. Western Blots
2.9. Statistical Analysis
2.10. Data Availability
3. Results
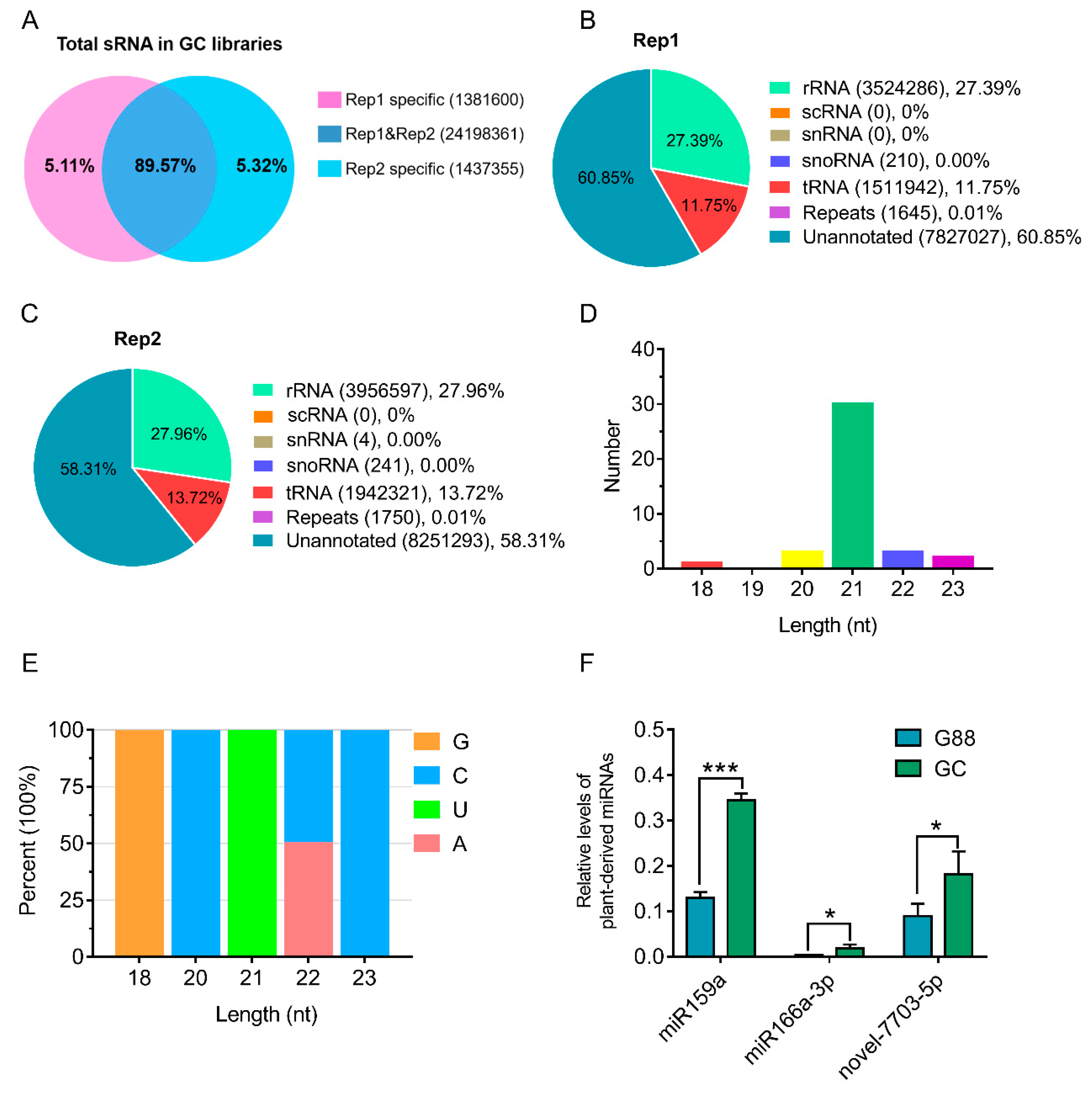
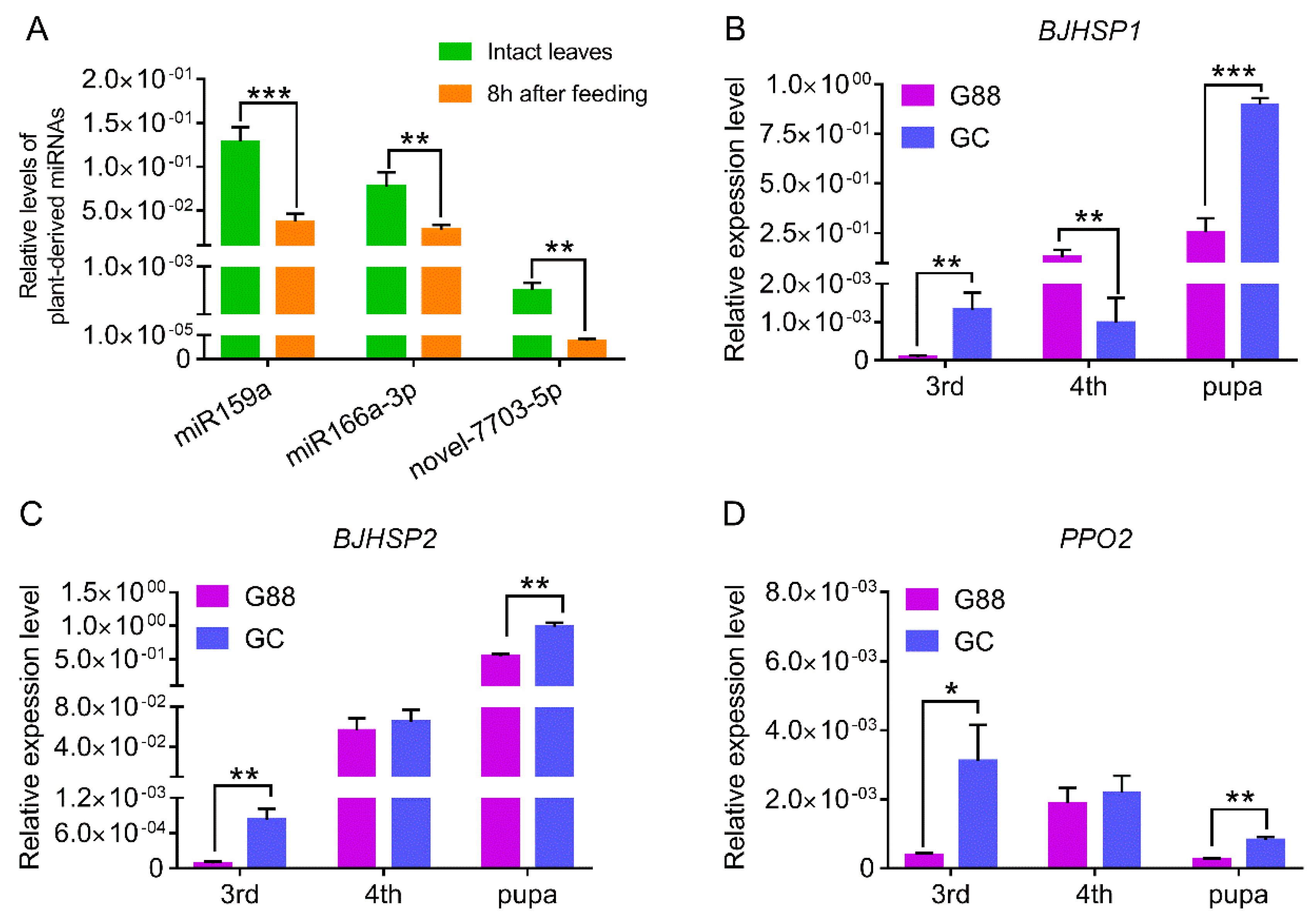
3.1. Characterization of Plant-Derived miRNAs in Larval Hemolymph of P. xylostella
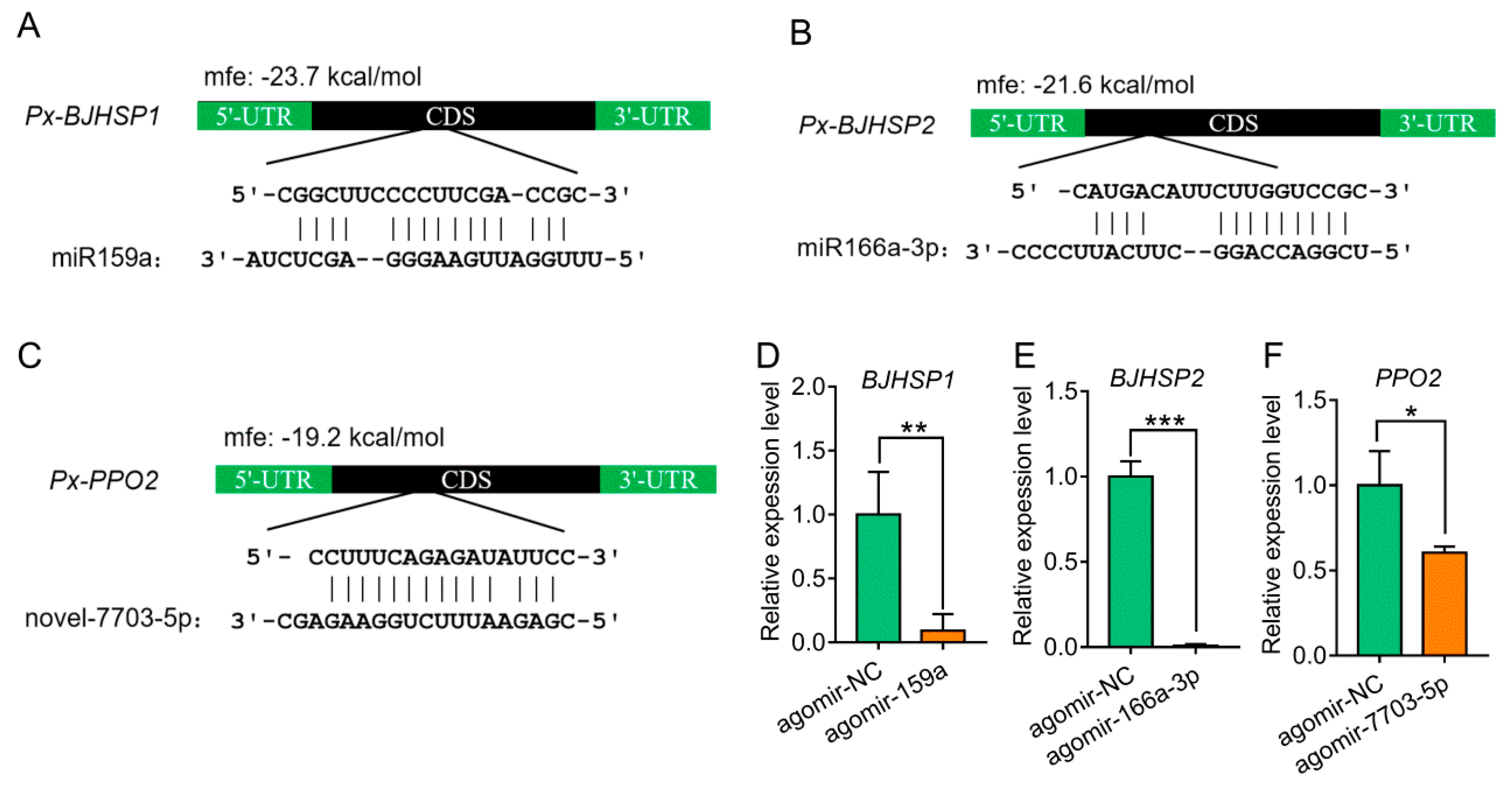
3.2. Target Prediction of Plant-Derived miRNAs in P. xylostella
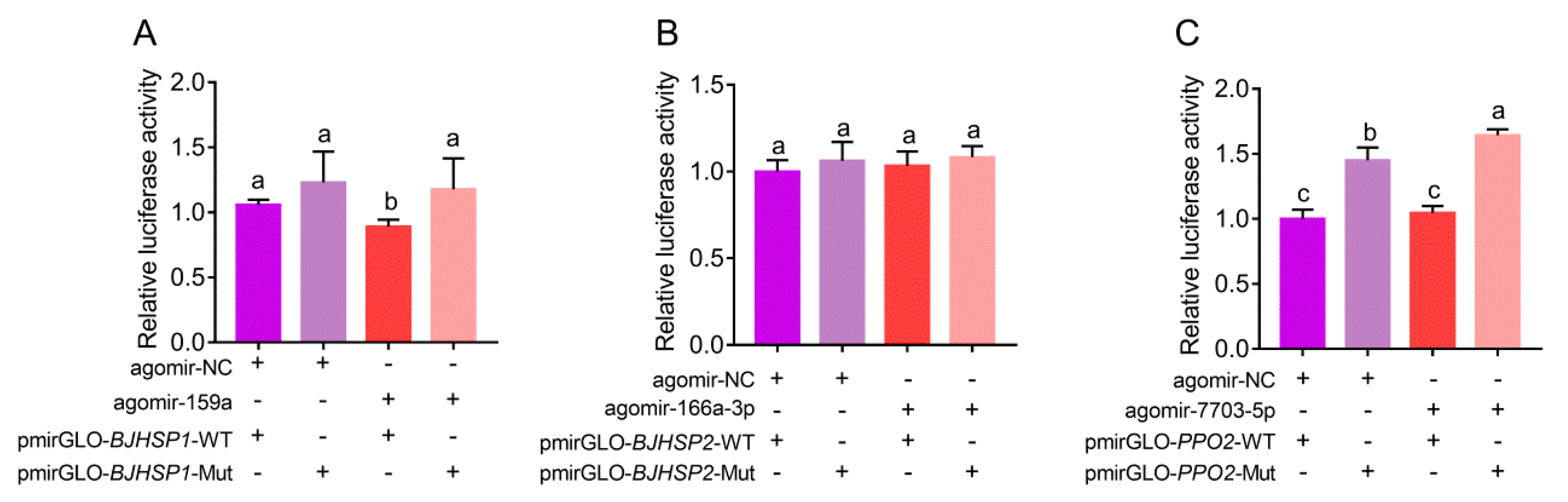
3.3. Suppression and Binding of Plant-Derived miRNAs to Target Genes in P. xylostella
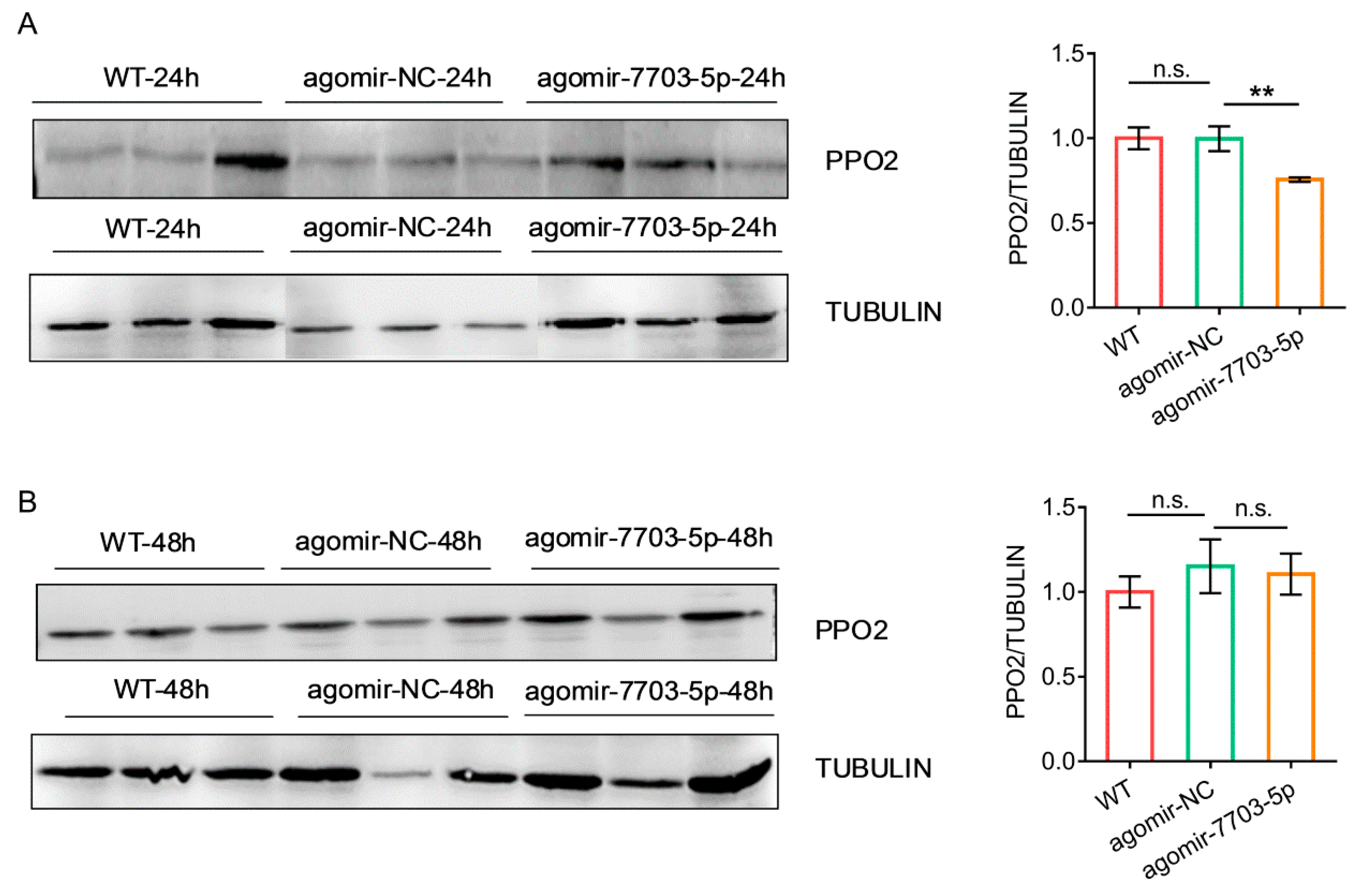
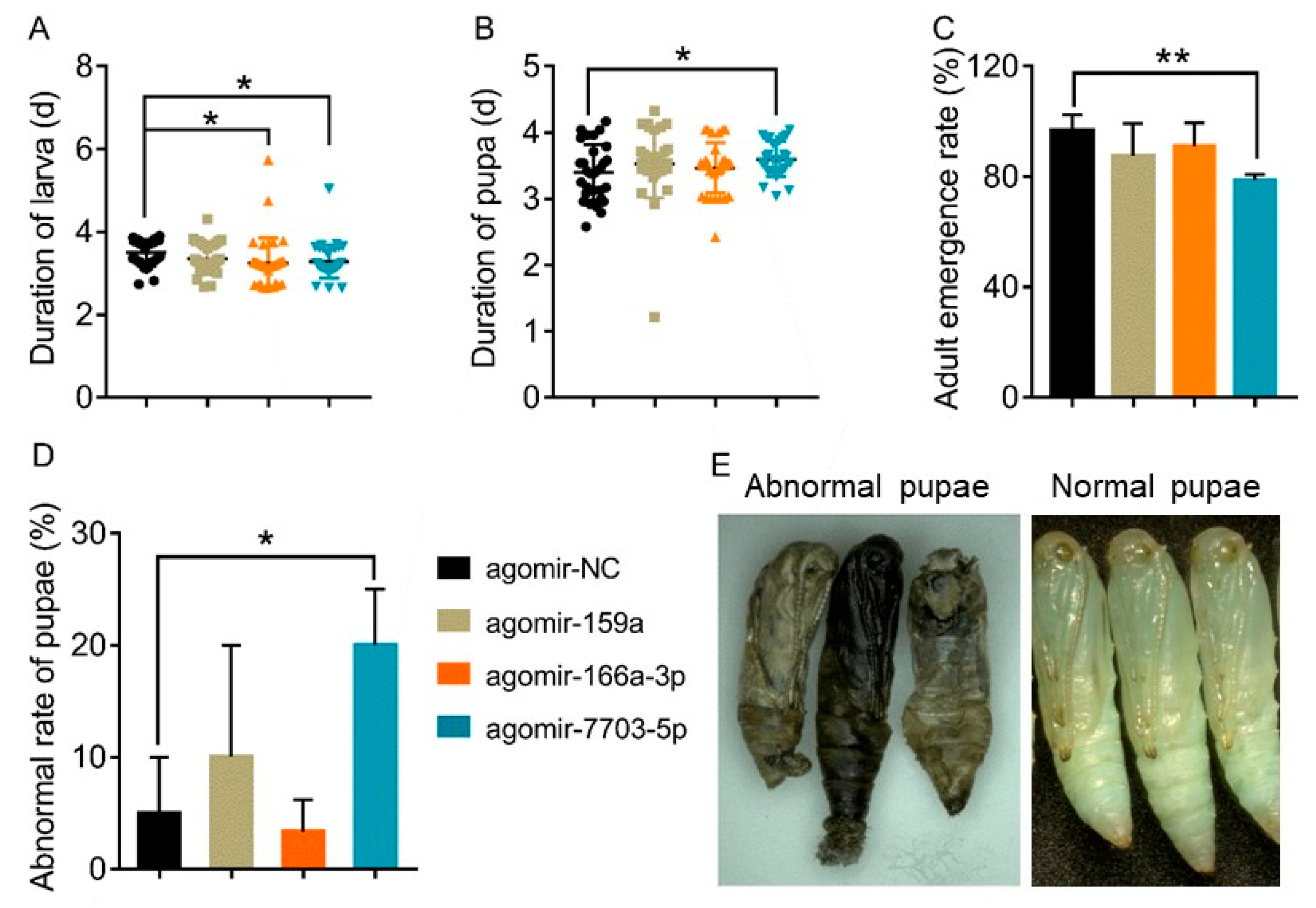
3.4. Effects of Plant-Derived miRNAs on the Development of P. xylostella
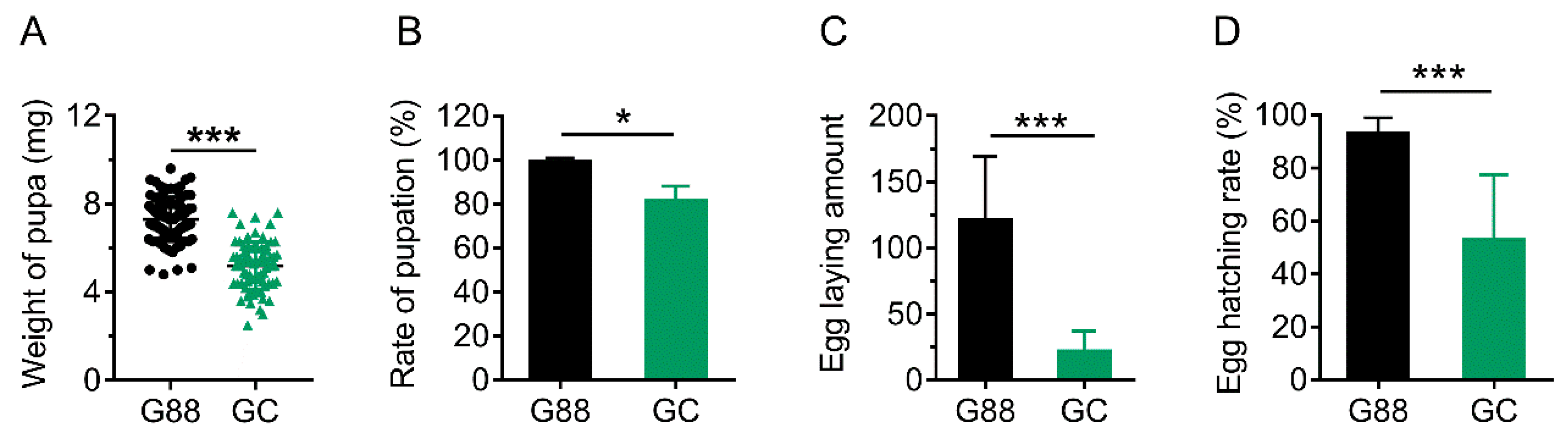
3.5. Link of Decreased Fitness of the Larvae on Host Plant with Plant-Derived miRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Carrington, J.C.; Ambros, V. Role of microRNAs in plant and animal development. Science 2003, 301, 336–338. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J.; Westholm, J.O.; Lai, E.C. Vive la différence: Biogenesis and evolution of microRNAs in plants and animals. Genome Biol. 2011, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Agron, M.; Praher, D.; Technau, U. The evolutionary origin of plant and animal microRNAs. Nat. Ecol. Evol. 2017, 1, 27. [Google Scholar] [CrossRef]
- Yu, B.; Yang, Z.Y.; Li, J.J.; Minakhina, S.; Yang, M.C.; Padgett, R.W.; Steward, R.; Chen, X.M. Methylation as a crucial step in plant microRNA biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef]
- Stark, A.; Brennecke, J.; Bushati, N.; Russell, R.B.; Cohen, S.M. Animal microRNAs confer robustness to gene expression and have a significant impact on 3 ‘ UTR evolution. Cell 2005, 123, 1133–1146. [Google Scholar] [CrossRef]
- Jiang, M.X.; Sang, X.L.; Hong, Z. Beyond nutrients: Food-derived microRNAs provide cross-kingdom regulation. Bioessays 2012, 34, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Luan, Y.S. Horizontal transfer of small RNAs to and from plants. Front Plant Sci. 2015, 6, 1113. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.P.; Liu, Y.N.; Liu, H.M.; Wang, H.Y.; Jin, W.; Zhang, Y.M.; Zhang, C.; Xu, D. Role of plant microRNA in cross-species regulatory networks of humans. BMC Syst. Biol. 2016, 10, 60. [Google Scholar] [CrossRef]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-kingdom small RNAs among animals, plants and microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Kumar, S.; Ayachit, G.; Bhairappanavar, S.B.; Ansari, A.; Sharma, P.; Soni, S.; Das, J. Cross-kingdom regulation of putative miRNAs derived from happy tree in cancer pathway: A systems biology approach. Int. J. Mol. Sci. 2017, 18, 1191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, Y.; Sun, M.; Ji, H.; Dou, H.; Hu, J.; Yan, Y.; Wang, X.; Chen, L. Honeysuckle-encoded microRNA2911 inhibits Enterovirus 71 replication via targeting VP1 gene. Antiviral Res. 2018, 152, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Sala-Cirtog, M.; Marian, C.; Anghel, A. New insights of medicinal plant therapeutic activity-the miRNA transfer. Biomed. Pharmacother. 2015, 74, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wiggins, B.E.; Lawrence, C.; Petrick, J.; Ivashuta, S.; Heck, G. Analysis of plant-derived miRNAs in animal small RNA datasets. BMC Genomics 2012, 13, 381. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zhang, D.; Xiang, Z.; He, N. Nonfunctional ingestion of plant miRNAs in silkworm revealed by digital droplet PCR and transcriptome analysis. Sci. Rep. 2015, 5, 12290. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Liu, M.; Fu, Z.; Zhou, Z.; Kong, Y.; Liang, H.; Lin, Z.; Luo, J.; Zheng, H.; Wan, P.; et al. Plant microRNAs in larval food regulate honeybee caste development. PLoS Genet. 2017, 13, e1006946. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, C.; Dou, Y.; Yu, B.; Liu, Y.; Heng-Moss, T.M.; Lu, G.; Wachholtz, M.; Bradshaw, J.D.; Twigg, P.; et al. Insect and plant-derived miRNAs in greenbug (Schizaphis graminum) and yellow sugarcane aphid (Sipha flava) revealed by deep sequencing. Gene 2017, 599, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Masood, M.; Everett, C.P.; Chan, S.Y.; Snow, J.W. Negligible uptake and transfer of diet-derived pollen microRNAs in adult honey bees. RNA Biol. 2016, 13, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; McAlexander, M.A.; Queen, S.E.; Adams, R.J. Real-time quantitative PCR and droplet digital PCR for plant miRNAs in mammalian blood provide little evidence for general uptake of dietary miRNAs: Limited evidence for general uptake of dietary plant xenomiRs. RNA Biol. 2013, 10, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Mico, V.; Martin, R.; Lasuncion, M.A.; Ordovas, J.M.; Daimiel, L. Unsuccessful detection of plant microRNAs in beer, extra virgin olive oil and human plasma after an acute ingestion of extra virgin olive oil. Plant Foods Hum. Nutr. 2016, 71, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.; Zhang, Y.; Petrick, J.S.; Heck, G.; Ivashuta, S.; Marshall, W.S. Lack of detectable oral bioavailability of plant microRNAs after feeding in mice. Nat. Biotechnol. 2013, 31, 965. [Google Scholar] [CrossRef] [PubMed]
- Talekar, N.S.; Shelton, A.M. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, X.; Liu, S.S.; You, M.; Furlong, M.J. Biology, ecology, and management of the diamondback moth in China. Annu. Rev. Entomol. 2016, 61, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Guo, L.; Zhou, X.G.; Gao, X.W.; Liang, P. miRNAs regulated overexpression of ryanodine receptor is involved in chlorantraniliprole resistance in Plutella xylostella (L.). Sci. Rep. 2015, 5, 14095. [Google Scholar] [CrossRef]
- Zhu, B.; Li, X.; Liu, Y.; Gao, X.; Liang, P. Global identification of microRNAs associated with chlorantraniliprole resistance in diamondback moth Plutella xylostella (L.). Sci. Rep. 2017, 7, 40713. [Google Scholar] [CrossRef]
- Etebari, K.; Afrad, M.H.; Tang, B.; Silva, R.; Furlong, M.J.; Asgari, S. Involvement of microRNA miR-2b-3p in regulation of metabolic resistance to insecticides in Plutella xylostella. Insect Mol. Biol. 2018, 27, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Xu, X.X.; Xu, J.; Li, S.Z.; Yu, J.L.; Zhou, X.Q.; Xu, X.J.; Hu, Q.B.; Yu, X.Q.; Jin, F.L. Genome-wide identification of Destruxin A-responsive immunity-related microRNAs in diamondback moth, Plutella xylostella. Front. Immunol. 2018, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, X.X.; Li, S.Z.; Wang, S.; Xu, X.J.; Zhou, X.Q.; Yu, J.L.; Yu, X.Q.; Shakeel, M.; Jin, F.L. Genome-wide profiling of Plutella xylostella immunity-related miRNAs after Isaria fumosorosea infection. Front. Physiol. 2017, 8, 1054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jiang, L.; Wang, J.; Gu, P.; Chen, M. MTide: An integrated tool for the identification of miRNA-target interaction in plants. Bioinformatics 2015, 31, 290–291. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Asgari, S. Revised annotation of Plutella xylostella microRNAs and their genome-wide target identification. Insect Mol. Biol. 2016, 25, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.L.; Wu, Q.J.; Liu, Y.; Zhou, X.M.; Zhou, X.G.; Zhang, Y.J. Exploring valid reference genes for quantitative real-time PCR analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792–802. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, 149–153. [Google Scholar] [CrossRef]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Yu, L.; He, W.; Yang, G.; Ke, F.; Baxter, S.W.; You, S.; Douglas, C.J.; You, M. DBM-DB: The diamondback moth genome database. Database 2014, 2014, 87. [Google Scholar] [CrossRef] [PubMed]
- He, W.; You, M.; Vasseur, L.; Yang, G.; Xie, M.; Cui, K.; Bai, J.; Liu, C.; Li, X.; Xu, X.; et al. Developmental and insecticide-resistant insights from the de novo assembled transcriptome of the diamondback moth, Plutella xylostella. Genomics 2012, 99, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Scali, M.; Vignani, R.; Spadafora, A.; Sensi, E.; Mazzuca, S.; Cresti, M. Protein extraction for two-dimensional electrophoresis from olive leaf, a plant tissue containing high levels of interfering compounds. Electrophoresis 2003, 24, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, F.; Zhu, J.; Zhang, S.; Yang, Y.; Chen, T.; Tang, B.; Dong, L.; Ding, N.; Zhang, Q.; et al. GSA: Genome sequence archive. Genom. Proteom. Bioinf. 2017, 15, 14–18. [Google Scholar] [CrossRef] [PubMed]
- BIG Data Center Members. Database resources of the BIG data center in 2019. Nucleic Acids Res. 2018, 47, 8–14. [Google Scholar]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, 155–162. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Burmester, T. Origin and evolution of arthropod hemocyanins and related proteins. J. Comp. Physiol. B 2002, 172, 95–107. [Google Scholar]
- Ehlting, J.; Chowrira, S.G.; Mattheus, N.; Aeschliman, D.S.; Arimura, G.I.; Bohlmann, J. Comparative transcriptome analysis of Arabidopsis thaliana infested by diamondback moth (Plutella xylostella) larvae reveals signatures of stress response, secondary metabolism, and signalling. BMC Genomics 2008, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.W.; Hale, A.E.; Isaacs, S.K.; Baggish, A.L.; Chan, S.Y. Ineffective delivery of diet-derived microRNAs to recipient animal organisms. RNA Biol. 2013, 10, 1107–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Wong, A.Y.P.; Wang, S.; Jia, Q.; Chuang, W.P.; Bendena, W.G.; Tobe, S.S.; Yang, S.H.; Chung, G.; Chan, T.F.; et al. miRNA-mediated interactions in and between plants and insects. Int. J. Mol. Sci. 2018, 19, 3239. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Snow, J.W. Uptake and impact of natural diet-derived small RNA in invertebrates: Implications for ecology and agriculture. RNA Biol. 2017, 14, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Karasov, W.H.; Martinez del Rio, C.; Caviedes-Vidal, E. Ecological physiology of diet and digestive systems. Annu. Rev. Physiol. 2011, 73, 69–93. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef]
- Rera, M.; Clark, R.I.; Walker, D.W. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 21528–21533. [Google Scholar] [CrossRef]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.L.; Barthel, A.; et al. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef]
- Samuel, M.; Bleackley, M.; Anderson, M.; Mathivanan, S. Extracellular vesicles including exosomes in cross kingdom regulation: A viewpoint from plant-fungal interactions. Front. Plant Sci. 2015, 6, 766. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, X.; Zheng, Z.; Zheng, J.; Shakeel, M.; Jin, F. MicroRNA expression profiling of Plutella xylostella after challenge with B. thuringiensis. Dev. Comp. Immunol. 2019, 93, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Melzig, M.F. The stability of medicinal plant microRNAs in the herb preparation process. Molecules 2018, 23, 919. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Mature Sequence (5′–3′) | Read Count | |
|---|---|---|---|
| Rep1 | Rep2 | ||
| miR156a-5p | UGACAGAAGAGAGUGAGCAC | 6 | 3 |
| miR156b-3p | UGCUCACCUCUCUUUCUGUCAGU | 6 | 4 |
| miR157a-5p | UUGACAGAAGAUAGAGAGCAC | 0 | 2 |
| miR159a | UUUGGAUUGAAGGGAGCUCUA | 204 | 313 |
| miR159b-3p | UUUGGAUUGAAGGGAGCUCUU | 42 | 80 |
| miR159c | UUUGGAUUGAAGGGAGCUCCU | 15 | 18 |
| miR160a-5p | UGCCUGGCUCCCUGUAUGCCA | 1 | 1 |
| miR161.1 | UGAAAGUGACUACAUCGGGGU | 1 | 3 |
| miR162a-3p | UCGAUAAACCUCUGCAUCCAG | 50 | 58 |
| miR164a | UGGAGAAGCAGGGCACGUGCA | 6 | 9 |
| miR164c-5p | UGGAGAAGCAGGGCACGUGCG | 3 | 0 |
| miR165a-3p | UCGGACCAGGCUUCAUCCCCC | 1 | 3 |
| miR166a-3p | UCGGACCAGGCUUCAUUCCCC | 120 | 151 |
| miR167a-5p | UGAAGCUGCCAGCAUGAUCUA | 24 | 16 |
| miR167d | UGAAGCUGCCAGCAUGAUCUGG | 22 | 13 |
| miR168a-3p | UCGCUUGGUGCAGGUCGGGAA | 4 | 9 |
| miR168a-5p | CCCGCCUUGCAUCAACUGAAU | 3 | 8 |
| miR171a-3p | UGAUUGAGCCGCGCCAAUAUC | 0 | 2 |
| miR172a | AGAAUCUUGAUGAUGCUGCAU | 7 | 3 |
| miR172c | AGAAUCUUGAUGAUGCUGCAG | 2 | 0 |
| miR319a | UUGGACUGAAGGGAGCUCCCU | 8 | 17 |
| miR390a-5p | AAGCUCAGGAGGGAUAGCGCC | 1 | 4 |
| miR394a | UUGGCAUUCUGUCCACCUCC | 6 | 7 |
| miR395a | CUGAAGUGUUUGGGGGAACUC | 6 | 10 |
| miR396a-3p | GUUCAAUAAAGCUGUGGGAAG | 58 | 85 |
| miR396a-5p | UUCCACAGCUUUCUUGAACUG | 24 | 28 |
| miR396b-3p | GCUCAAGAAAGCUGUGGGAAA | 1 | 0 |
| miR396b-5p | UUCCACAGCUUUCUUGAACUU | 23 | 29 |
| miR398a-3p | UGUGUUCUCAGGUCACCCCUU | 0 | 2 |
| miR398b-3p | UGUGUUCUCAGGUCACCCCUG | 0 | 2 |
| miR403-3p | UUAGAUUCACGCACAAACUCG | 7 | 8 |
| miR408-3p | AUGCACUGCCUCUUCCCUGGC | 6 | 1 |
| miR858b | UUCGUUGUCUGUUCGACCUUG | 11 | 17 |
| novel-2599-3p | AUCCGUGGUUUCGCGUAUCGGC | 0 | 4 |
| novel-2783-3p | CGCGGAGAAGGGGAAGGGGUGCU | 0 | 7 |
| novel-6634-5p | CUGAGAAUUUCUGGAAGAGCUC | 9 | 15 |
| novel-7211-3p | UCUCGGACCAGGCUUCAUUCC | 10 | 8 |
| novel-7477-5p | GACUGUAAGGCUGUGGAC | 5 | 1 |
| novel-7703-5p | CGAGAAUUUCUGGAAGAGCU | 100 | 101 |
| miRNA | Gene ID | Agomir-NC 1 | miRNA Agomir 1 | log2 Ratio 2 | q-Value 3 | p-Value 3 | Annotation (DBM-DB) |
|---|---|---|---|---|---|---|---|
| miR159a | Px000121 | 28.56 | 0 | −5.77 | 6.95E-08 | 3.88E-08 | Peroxisomal targeting signal 1 receptor |
| Px002198 | 1463 | 383 | −1.87 | 8.38E-137 | 3.78E-138 | 4-coumarate-CoA ligase-like 5 | |
| Px002761 | 88 | 26 | −1.69 | 1.87E-08 | 9.88E-09 | uncharacterized protein | |
| Px003612 | 1317.25 | 604.38 | −1.06 | 5.86E-53 | 6.26E-54 | Sterol | |
| O-acyltransferase 2 | |||||||
| Px006820 | 59809 | 362 | −7.3 | 0 | 0 | Basic juvenile hormone-suppressible protein 1 | |
| Px006897 | 1731.5 | 762.4 | −1.12 | 1.42E-75 | 1.12E-76 | Venom serine protease 34 | |
| Px007030 | 21806.66 | 205.71 | −6.66 | 0 | 0 | Basic juvenile hormone-suppressible protein 2 | |
| Px007031 | 47614.34 | 4669.29 | −3.28 | 0 | 0 | Basic juvenile hormone-suppressible protein 2 | |
| Px009267 | 54 | 11 | −2.23 | 1.22E-07 | 7.03E-08 | Nucleic-acid-binding protein from mobile element jockey | |
| Px010115 | 97.12 | 38.01 | −1.29 | 1.39E-06 | 9.05E-07 | Moricin-like peptide C5 | |
| Px013616 | 76.95 | 13.11 | −2.49 | 1.64E-11 | 6.62E-12 | Unknown | |
| Px015810 | 193 | 91 | −1.02 | 1.83E-08 | 9.64E-09 | uncharacterized protein | |
| miR166a-3p | Px007031 | 47614.34 | 852.67 | −5.83 | 0 | 0 | Basic juvenile hormone-suppressible protein 2 |
| novel-7703-5p | Px002274 | 4409.62 | 1854.67 | −1.08 | 1.34E-174 | 7.24E-176 | Phenoloxidase subunit 2 |
| Px003522 | 348.65 | 15.57 | −4.31 | 8.01E-73 | 1.03E-73 | Phospholipase A-2-activating protein | |
| Px003840 | 72050.28 | 29765.48 | −1.1 | 0 | 0 | Actin, muscle | |
| Px004854 | 1931.79 | 808.05 | −1.09 | 3.96E-78 | 4.69E-79 | Ecdysteroid UDP-glucosyltransferase | |
| Px006730 | 10017.44 | 4288.62 | −1.05 | 0 | 0 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial | |
| Px007989 | 72.83 | 29.51 | −1.13 | 1.37E-04 | 1.57E-04 | Elongation factor Tu, mitochondrial | |
| Px008572 | 48906.53 | 18617.36 | −1.22 | 0 | 0 | Transketolase | |
| Px009077 | 7787.63 | 2753.94 | −1.33 | 0 | 0 | ATP-citrate synthase |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.-l.; Jing, X.-d.; Chen, W.; Wang, Y.; Lin, J.-h.; Zheng, L.; Dong, Y.-h.; Zhou, L.; Li, F.-f.; Yang, F.-y.; et al. Host Plant-Derived miRNAs Potentially Modulate the Development of a Cosmopolitan Insect Pest, Plutella xylostella. Biomolecules 2019, 9, 602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100602
Zhang L-l, Jing X-d, Chen W, Wang Y, Lin J-h, Zheng L, Dong Y-h, Zhou L, Li F-f, Yang F-y, et al. Host Plant-Derived miRNAs Potentially Modulate the Development of a Cosmopolitan Insect Pest, Plutella xylostella. Biomolecules. 2019; 9(10):602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100602
Chicago/Turabian StyleZhang, Ling-ling, Xiao-dong Jing, Wei Chen, Yue Wang, Jun-han Lin, Ling Zheng, Yu-hong Dong, Li Zhou, Fei-fei Li, Fei-ying Yang, and et al. 2019. "Host Plant-Derived miRNAs Potentially Modulate the Development of a Cosmopolitan Insect Pest, Plutella xylostella" Biomolecules 9, no. 10: 602. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100602