A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of Phyllomedusa tarsius Skin Secretion
2.2. “Shotgun” Cloning of DPT9 Precursor-Encoding cDNA from a Skin Secretion-Derived cDNA Library
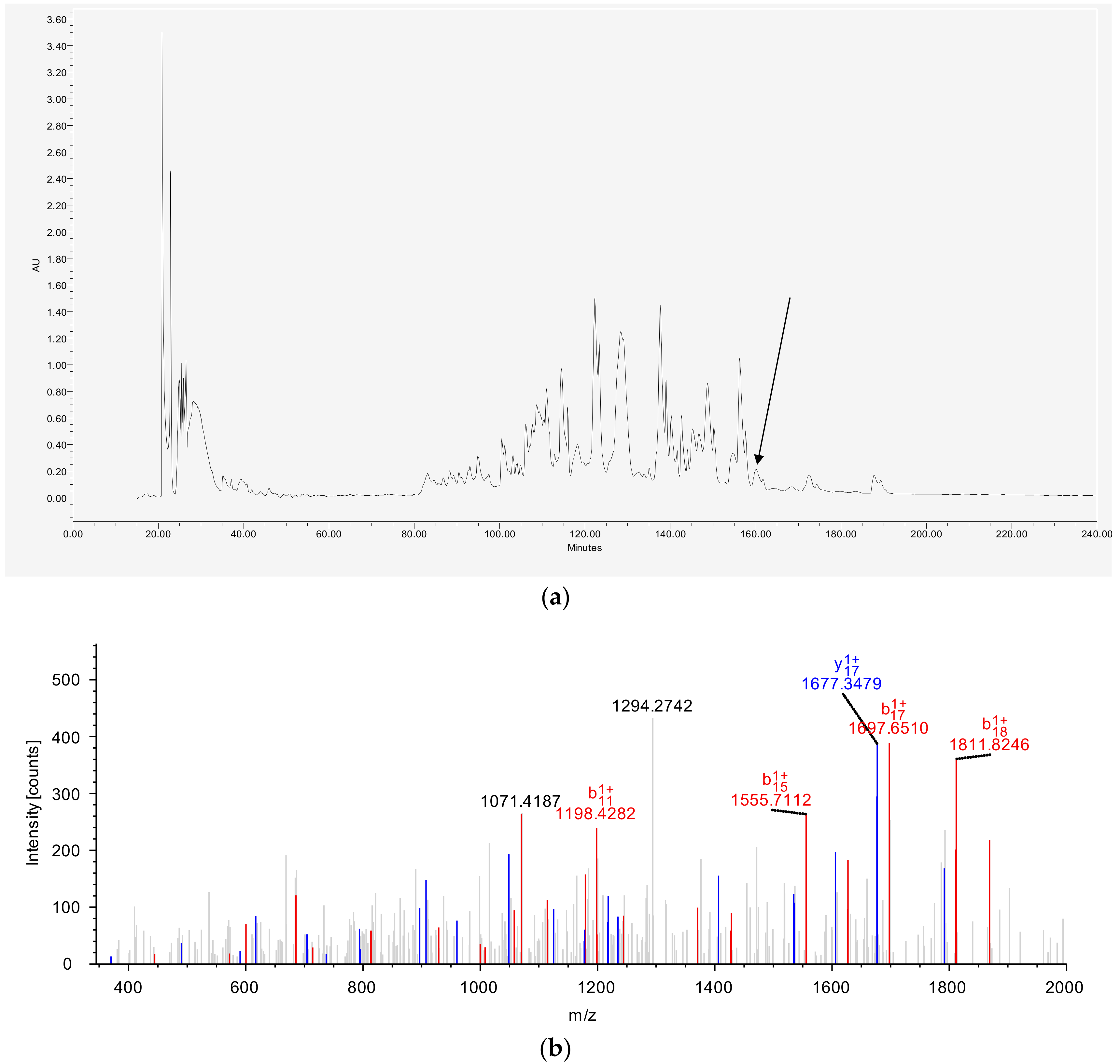
2.3. Isolation and Identification of DPT9 from Crude Skin Secretion
2.4. Chemical Synthesis of DPT9 and K8, 23-DTP9
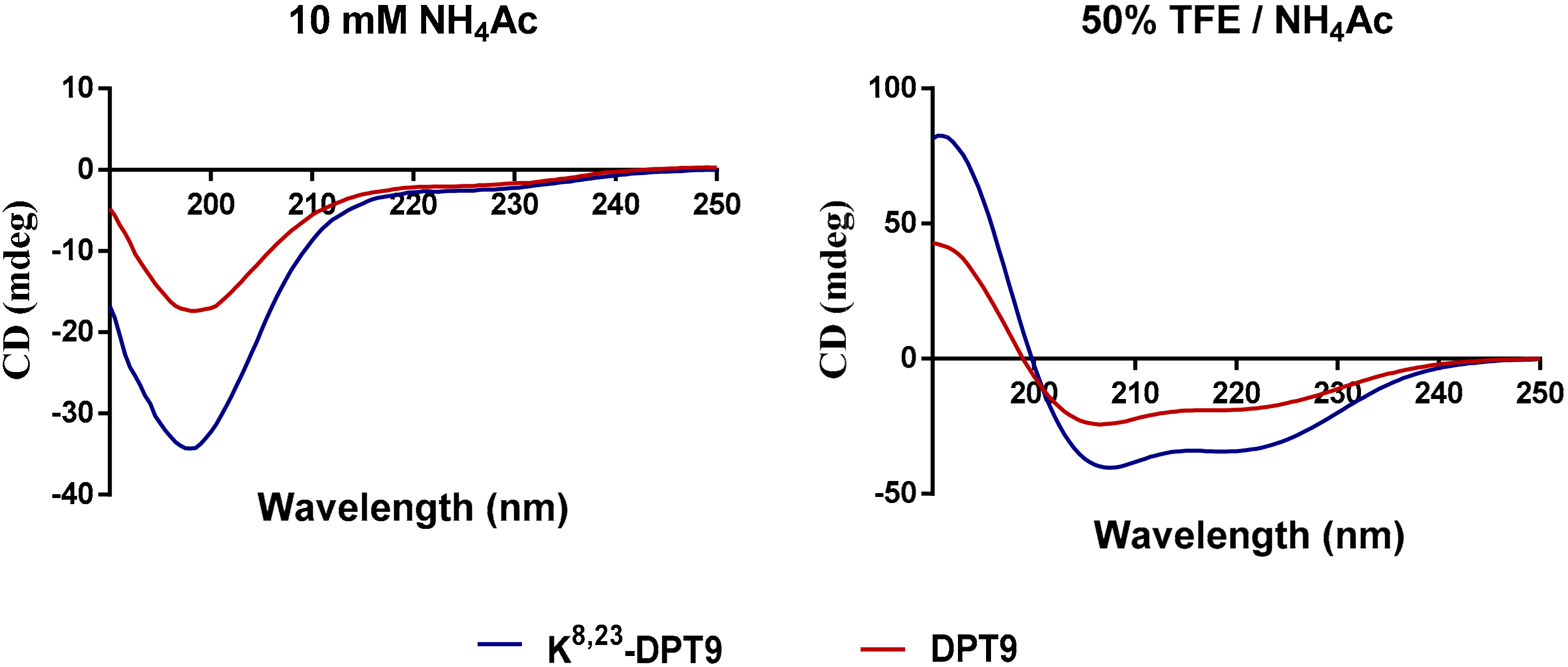
2.5. Secondary Structure Analysis of DPT9 and K8, 23-DTP9
2.6. Antimicrobial and Antibiofilm Susceptibility Assay
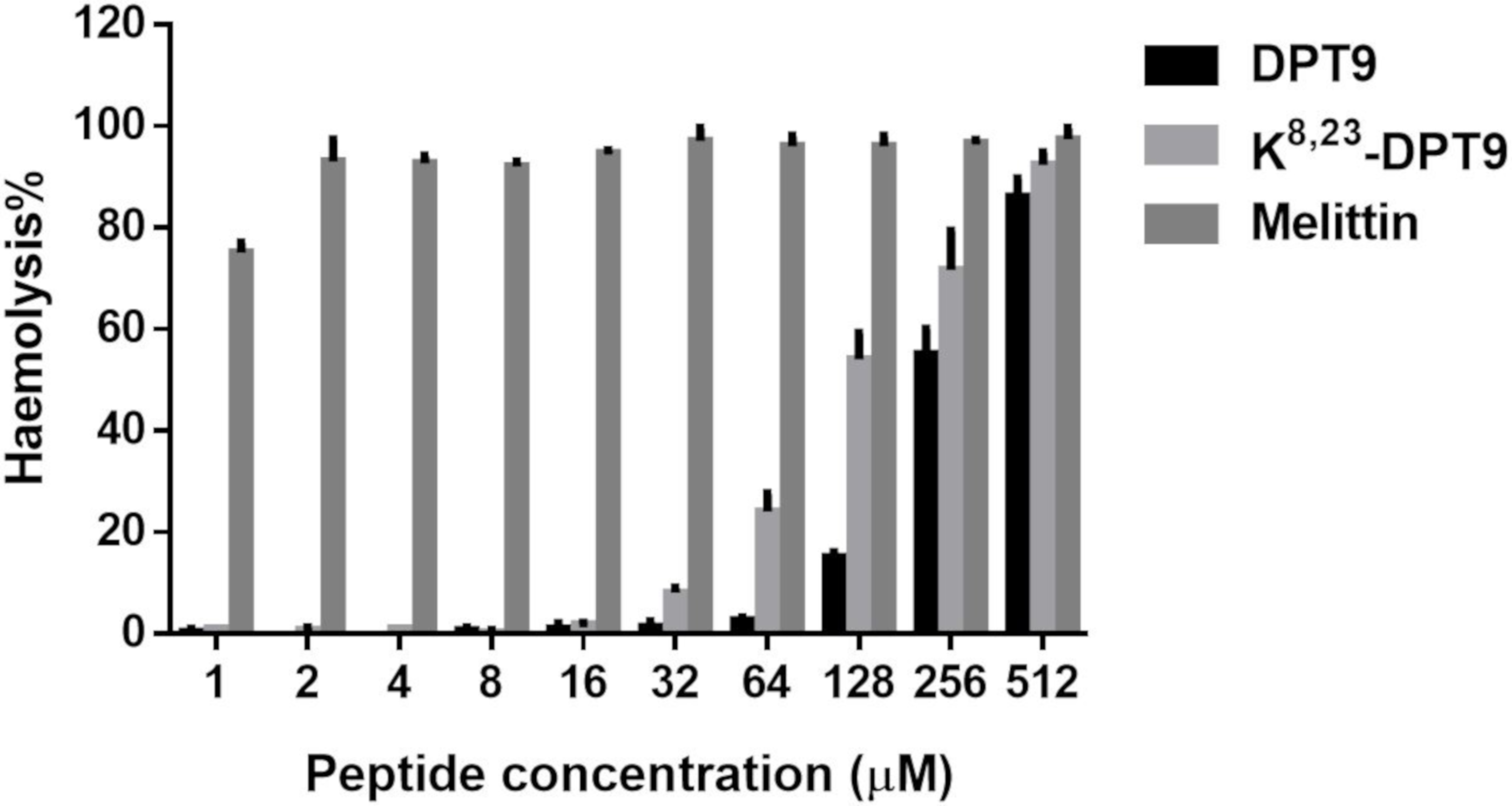
2.7. Haemolysis Assay
2.8. Membrane Permeability Assay
2.9. MTT Assay
2.10. Lactate Dehydrogenase (LDH) Assay
3. Results
3.1. “Shotgun” Cloning of DPT9 Biosynthetic Precursor cDNA from a Skin Secretion-Derived cDNA Library of Phyllomedusa tarsius
3.2. Identification and Structural Characterization of DPT9
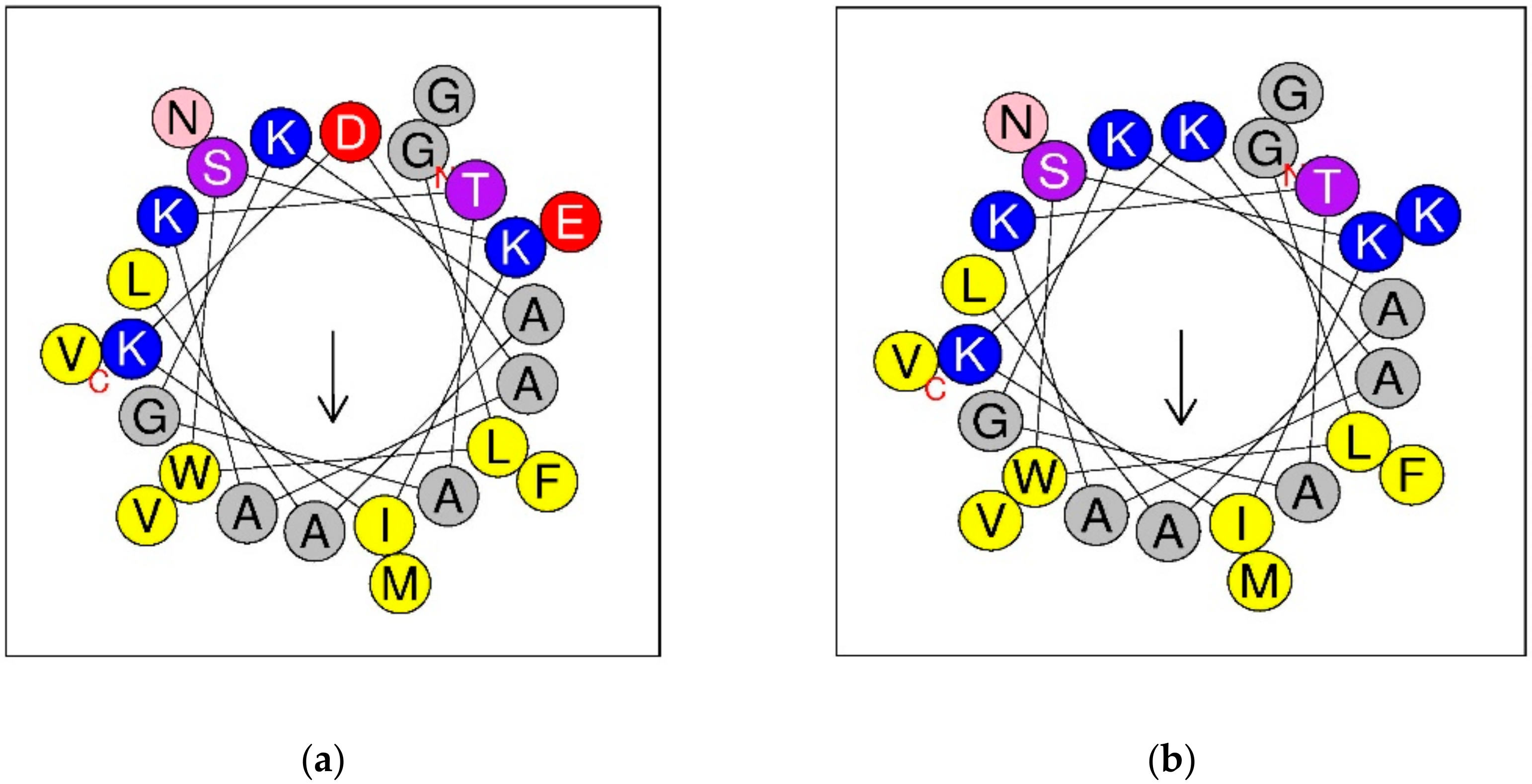
3.3. Synthesis, Predicted Physicochemical Properties and Secondary Structure of DPT9 and its Analogue K8, 23-DTP9
3.4. Antimicrobial and Haemolytic Activities of DPT9 and its Analogue K8, 23-DTP9
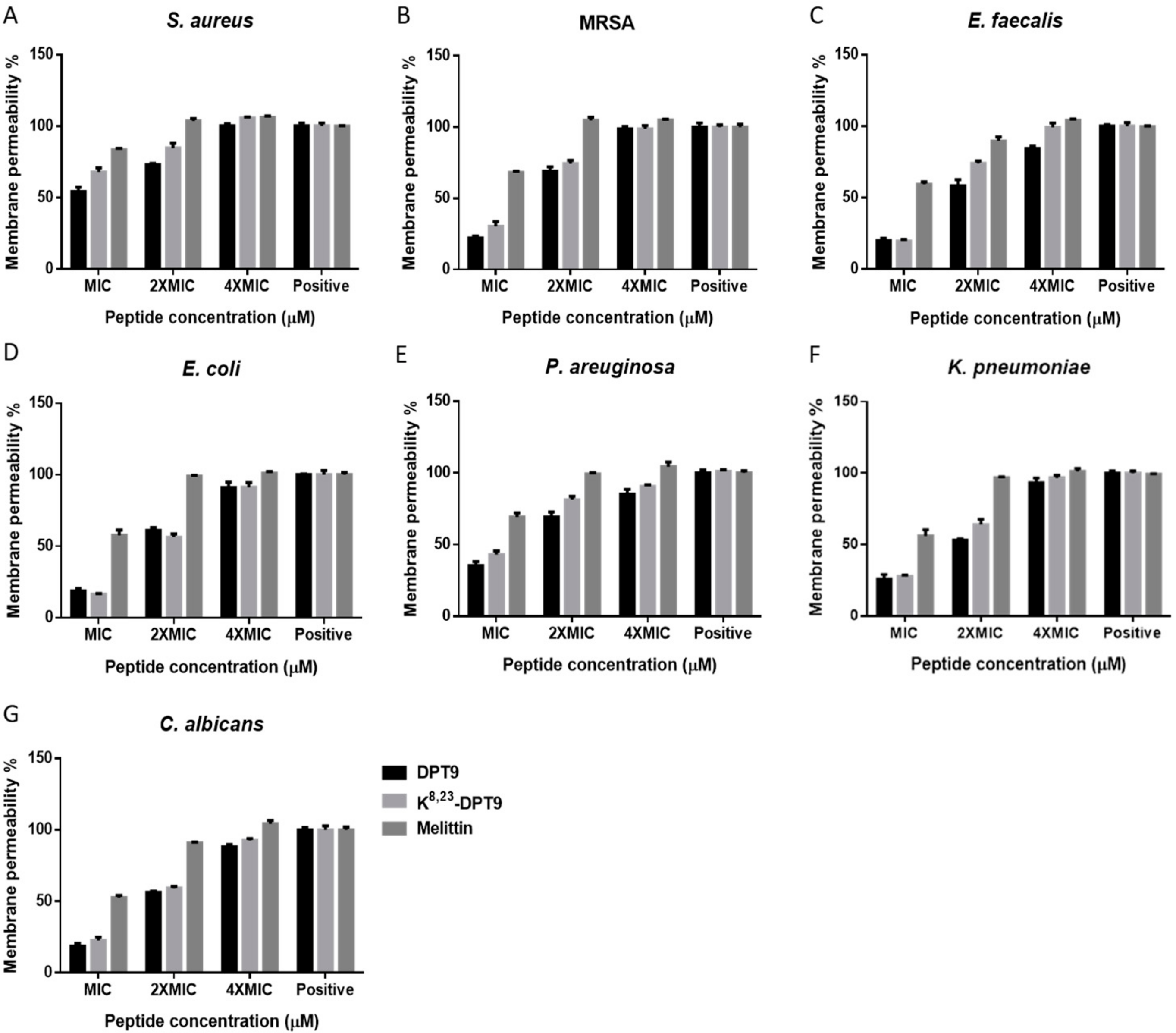
3.5. Membrane Permeabilisation of DPT9 and Its Analogue K8, 23-DTP9
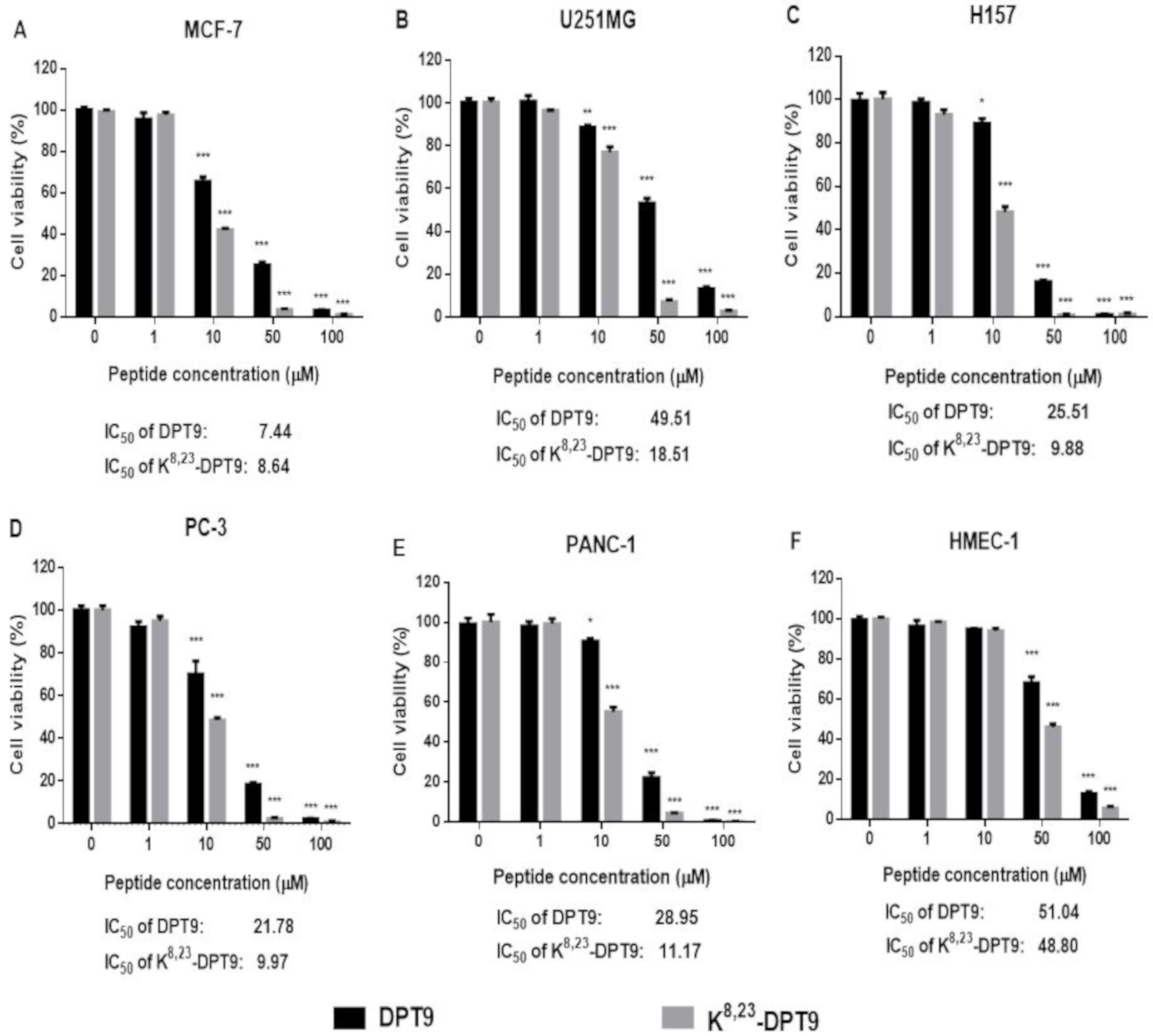
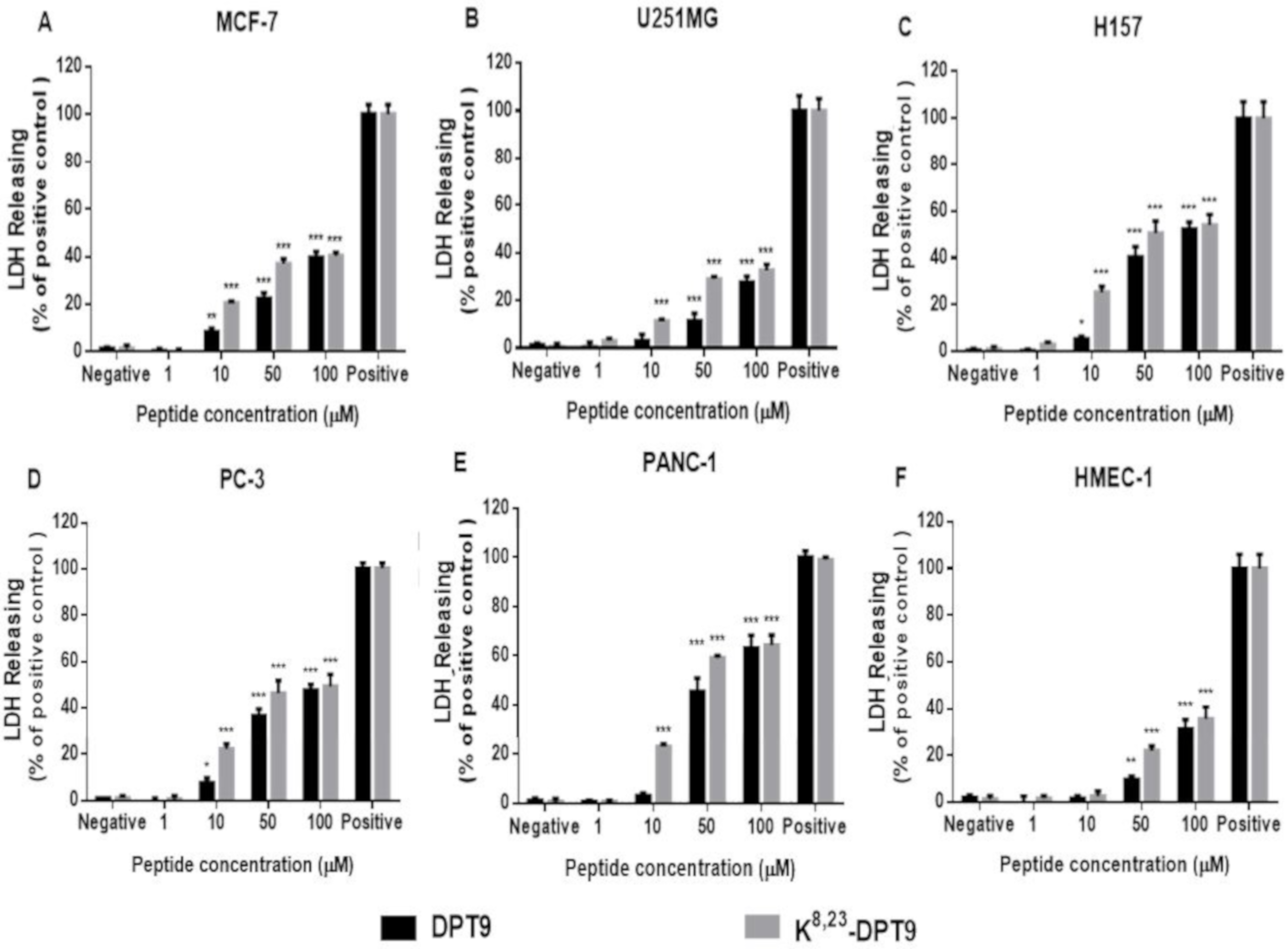
3.6. MTT and LDH Assays on the Human Cancer and Normal Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Lee, T.H.; Hall, K.N.; Aguilar, M.I. Antimicrobial Peptide Structure and Mechanism of Action: A Focus on the Role of Membrane Structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, P.; El Amri, C. The dermaseptin superfamily: A gene-based combinatorial library of antimicrobial peptides. Biochim. Biophys. Acta 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Zairi, A.; Tangy, F.; Bouassida, K.; Hani, K. Dermaseptins and magainins: Antimicrobial peptides from frogs’ skin-new sources for a promising spermicides microbicides-a mini review. J. Biomed. Biotechnol. 2009, 2009, 452567. [Google Scholar] [CrossRef]
- Mor, A.; Nguyen, V.H.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, amino acid sequence, and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef]
- Mor, A.; Nicolas, P. Isolation and structure of novel defensive peptides from frog skin. Eur. J. Biochem. 1994, 219, 145–154. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef]
- Galanth, C.; Abbassi, F.; Lequin, O.; Ayala-Sanmartin, J.; Ladram, A.; Nicolas, P.; Amiche, M. Mechanism of antibacterial action of dermaseptin B2: Interplay between helix-hinge-helix structure and membrane curvature strain. Biochemistry 2009, 48, 313–327. [Google Scholar] [CrossRef]
- Shi, D.; Hou, X.; Wang, L.; Gao, Y.; Wu, D.; Xi, X.; Zhou, M.; Kwok, H.F.; Duan, J.; Chen, T.; et al. Two Novel Dermaseptin-Like Antimicrobial Peptides with Anticancer Activities from the Skin Secretion of Pachymedusa dacnicolor. Toxins 2016, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ding, X.; Li, W.; Lu, T.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burden, R.; Chen, T. Discovery of two skin-derived dermaseptins and design of a TAT-fusion analogue with broad-spectrum antimicrobial activity and low cytotoxicity on healthy cells. Peer. J. 2018, 6, e5635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zoggel, H.; Hamma-Kourbali, Y.; Galanth, C.; Ladram, A.; Nicolas, P.; Courty, J.; Amiche, M.; Delbe, J. Antitumor and angiostatic peptides from frog skin secretions. Amino Acids 2012, 42, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chen, X.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Kwok, H.F.; Chen, T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of. Toxins 2018, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhou, X.; Chen, X.; Huang, L.; Xi, X.; Ma, C.; Zhou, M.; Wang, L.; Chen, T. Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii. Molecules 2019, 24, 2974. [Google Scholar] [CrossRef]
- Mechkarska, M.; Coquet, L.; Leprince, J.; Auguste, R.J.; Jouenne, T.; Mangoni, M.L.; Conlon, J.M. Peptidomic analysis of the host-defense peptides in skin secretions of the Trinidadian leaf frog Phyllomedusa trinitatis (Phyllomedusidae). Comp Biochem. Physiol. Part D Genom. Proteom. 2018, 28, 72–79. [Google Scholar] [CrossRef]
- Roggemann, M.C.; Tyler, D.W.; Bilmont, M.F. Linear reconstruction of compensated images: Theory and experimental results. Appl. Opt. 1992, 31, 7429–7441. [Google Scholar] [CrossRef]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific alpha-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, Ranatuerin-2PLx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38, 6. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucl. Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo Calderon, L.; Silva, A.E.A.; Ciancaglini, P.; Stabeli, R.G. Antimicrobial peptides from Phyllomedusa frogs: From biomolecular diversity to potential nanotechnologic medical applications. Amino Acids 2011, 40, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic alpha helical antimicrobial peptides. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Wiradharma, N.; Sng, M.Y.; Khan, M.; Ong, Z.Y.; Yang, Y.Y. Rationally designed α-helical broad-spectrum antimicrobial peptides with idealized facial amphiphilicity. Macromol. Rapid. Commun. 2013, 34, 74–80. [Google Scholar] [CrossRef]
- Wang, K.; Yan, J.; Dang, W.; Liu, X.; Chen, R.; Zhang, J.; Zhang, B.; Zhang, W.; Kai, M.; Yan, W.; et al. Membrane active antimicrobial activity and molecular dynamics study of a novel cationic antimicrobial peptide polybia-MPI, from the venom of Polybia paulista. Peptides 2013, 39, 80–88. [Google Scholar] [CrossRef]
- Kustanovich, I.; Shalev, D.E.; Mikhlin, M.; Gaidukov, L.; Mor, A. Structural requirements for potent versus selective cytotoxicity for antimicrobial dermaseptin S4 derivatives. J. Biol. Chem. 2002, 277, 16941–16951. [Google Scholar] [CrossRef]
- Du, D.; Bunagan, M.R.; Gai, F. The effect of charge-charge interactions on the kinetics of alpha-helix formation. Biophys. J. 2007, 93, 4076–4082. [Google Scholar] [CrossRef]
- Gaidukov, L.; Fish, A.; Mor, A. Analysis of membrane-binding properties of dermaseptin analogues: Relationships between binding and cytotoxicity. Biochemistry 2003, 42, 12866–12874. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, F. Cationic amphiphilic peptides with cancer-selective toxicity. Eur. J. Pharmacol. 2009, 625, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Van Zoggel, H.; Carpentier, G.; Dos Santos, C.; Hamma-Kourbali, Y.; Courty, J.; Amiche, M.; Delbe, J. Antitumor and angiostatic activities of the antimicrobial peptide dermaseptin B2. PLoS ONE 2012, 7, e44351. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Li, L.; Wang, H.; Li, M.; Wang, L.; Zhou, M.; Su, Q.; Chen, T.; Wu, Y. Novel peptide dermaseptin-PS1 exhibits anticancer activity via induction of intrinsic apoptosis signalling. J. Cell. Mol. Med. 2019, 23, 1300–1312. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPT9 | K8, 23-DPT9 | DPH a | DCA1 b | DDU1 b | DPS3 c | DPS4 d | |
|---|---|---|---|---|---|---|---|
| Molecular weight (Da) | 2605.10 | 2617.24 | 2535.78 | 3024.62 | 2911.46 | 2550.00 | 2948.53 |
| Hydrophobicity | 0.348 | 0.326 | 0.333 | 0.291 | 0.331 | 0.373 | 0.368 |
| Hydrophobic moment | 0.447 | 0.462 | 0.427 | 0.263 | 0.255 | 0.437 | 0.226 |
| Net charge | +2 | +6 | +1 | +3 | +3 | +2 | +3 |
| α-helix (%) | 24.18 | 46.14 | 35 | 25.3 | 27.9 | 44.9 | 67.6 |
| Microorganisms | MIC/MBC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| MIC/MBC (µg/mL) | ||||||||
| DPT9 | K8, 23-DPT9 | DPH | DCA1 | DDU1 | DPS3 a | DPS4 a | Melittin | |
| S. aureus NCTC 10788 | 16/32 | 2/4 | 32/64 | 4/16 | 4/16 | 256 | 4 | 1/2 |
| 41.7/83.4 | 5.2/10.4 | 81.1/162.2 | 12.1/48.4 | 11.6/46.4 | 652.5 | 11.8 | 2.8/5.7 | |
| Methicillin-resistant Staphylococcus aureus (MRSA) NCTC 12493 | 32/64 | 2/4 | >512/>512 | 8/32 | 4/16 | NA | 8 | 1/4 |
| 83.4/166.7 | 5.2/10.4 | >1298.4/>1298.4 | 24.2/96.8 | 11.6/46.4 | 23.6 | 2.8/11.4 | ||
| E. faecalis NCTC 12697 | 16/32 | 2/4 | NA | 128/256 | 64/128 | NA | 32 | 1/2 |
| 41.7/83.4 | 5.2/10.4 | 387.1/774.1 | 185.6/371.2 | 94.3 | 2.8/5.7 | |||
| E. coli NCTC 10418 | 8/16 | 2/4 | 16/16 | 4/16 | 4/16 | 8 | 8 | 2/4 |
| 20.8/41.7 | 5.2/10.4 | 40.6/40.6 | 12.1/48.4 | 11.6/46.4 | 20.4 | 23.6 | 5.7/11.4 | |
| P. aeruginosa ATCC 27853 | 16/32 | 4/8 | 64>512 | 8/32 | 4/16 | NA | 16 | 16/32 |
| 41.7/83.4 | 10.4/20.8 | 162.2/>1298.4 | 24.2/96.8 | 11.6/46.4 | 47.1 | 45.5/91.1 | ||
| K. pneumoniae ATCC 43816 | 8/16 | 2/4 | NA | 8/128 | 4/64 | NA | NA | 2/8 |
| 20.8/41.7 | 5.2/10.4 | 24.2/387.1 | 11.6/185.6 | 5.7/22.8 | ||||
| C. albicans NCYC 1467 | 64/128 | 4/8 | 16/64 | 4/16 | 4/16 | 4 | 4 | 2/4 |
| 166.7/333.4 | 10.4/20.8 | 40.6/162.2 | 12.1/48.4 | 11.6/46.4 | 10.2 | 11.8 | 5.7/11.4 | |
| Microorganisms | MBIC/MBEC (μM) a | |
|---|---|---|
| DPT9 | K8, 23-DPT9 | |
| S. aureus NCTC 10788 | 16/32 | 2/4 |
| MRSA NCTC 12493 | 32/64 | 2/4 |
| E. coli NCTC 10418 | 8/16 | 2/4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Xi, X.; Ma, C.; Chen, X.; Zhou, M.; Burrows, J.F.; Chen, T.; Wang, L. A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities. Biomolecules 2019, 9, 628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100628
Li M, Xi X, Ma C, Chen X, Zhou M, Burrows JF, Chen T, Wang L. A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities. Biomolecules. 2019; 9(10):628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100628
Chicago/Turabian StyleLi, Miaoran, Xinping Xi, Chengbang Ma, Xiaoling Chen, Mei Zhou, James F. Burrows, Tianbao Chen, and Lei Wang. 2019. "A Novel Dermaseptin Isolated from the Skin Secretion of Phyllomedusa tarsius and Its Cationicity-Enhanced Analogue Exhibiting Effective Antimicrobial and Anti-Proliferative Activities" Biomolecules 9, no. 10: 628. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100628