An Insight into Storage Lipid Synthesis by Yarrowia lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Conditions
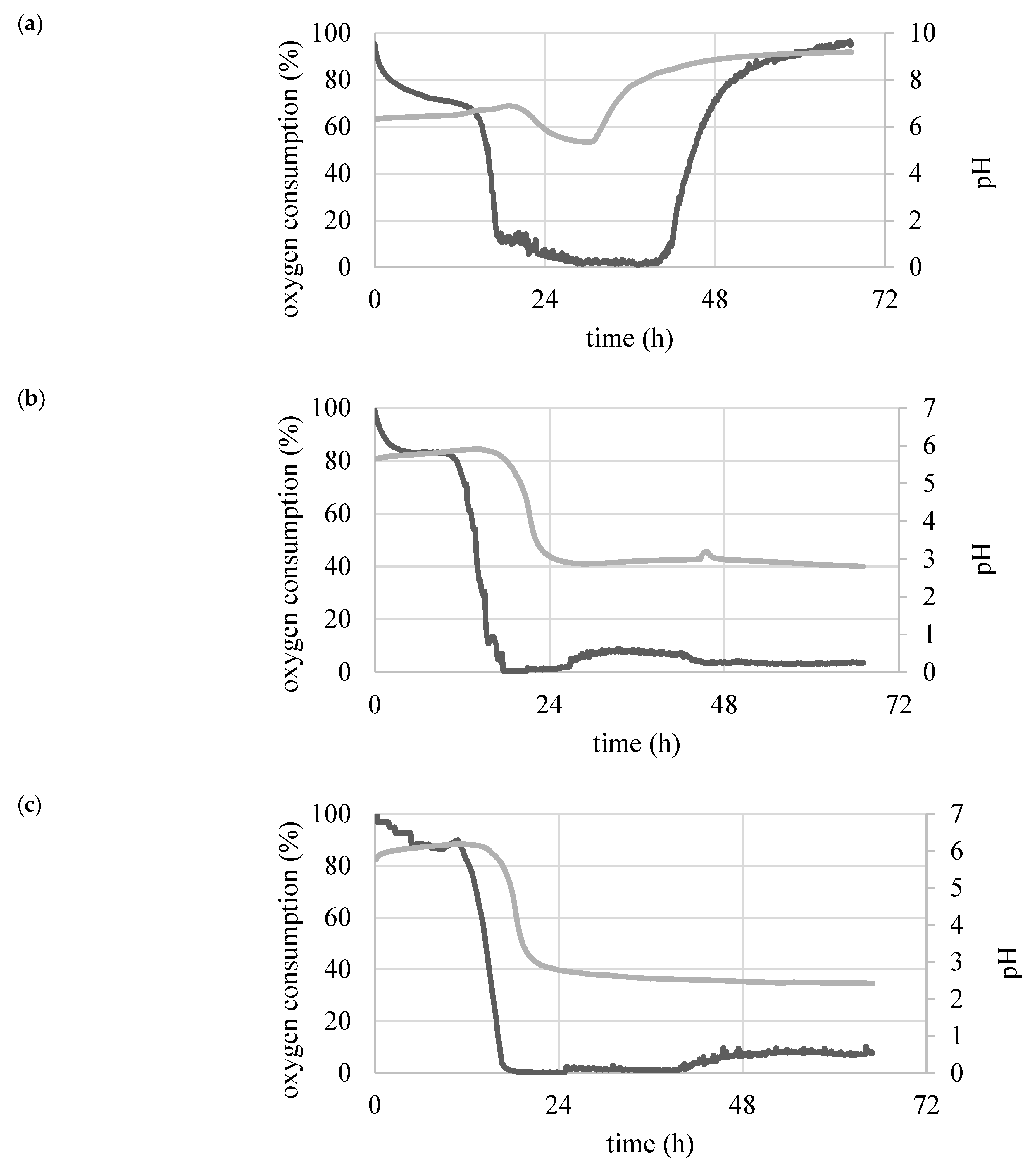
2.2. Determination of Biomass Yield, Glucose Concentration, pH, and Oxygen Consumption during Batch Culture
2.3. Lipid Extraction and Analysis
2.4. Determination of the mRNA Transcription Profiles of POX2 and YALI0 Genes
2.4.1. RNA Isolation
2.4.2. Reverse Transcription and Real Time PCR
2.5. Statistical Analysis
3. Results
3.1. Impact of Carbon Source on Synthesis of Lipids in Yeast Cells
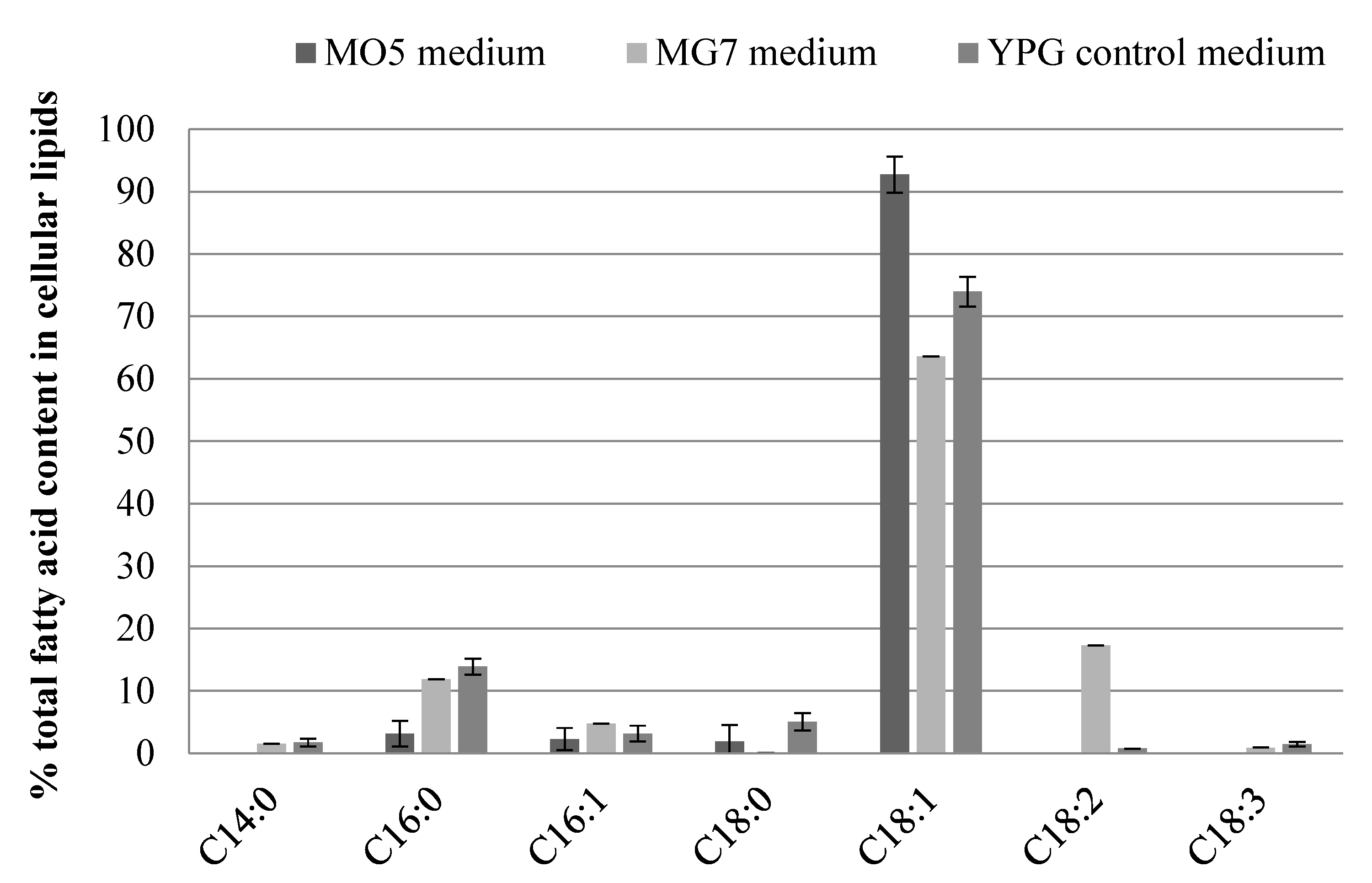
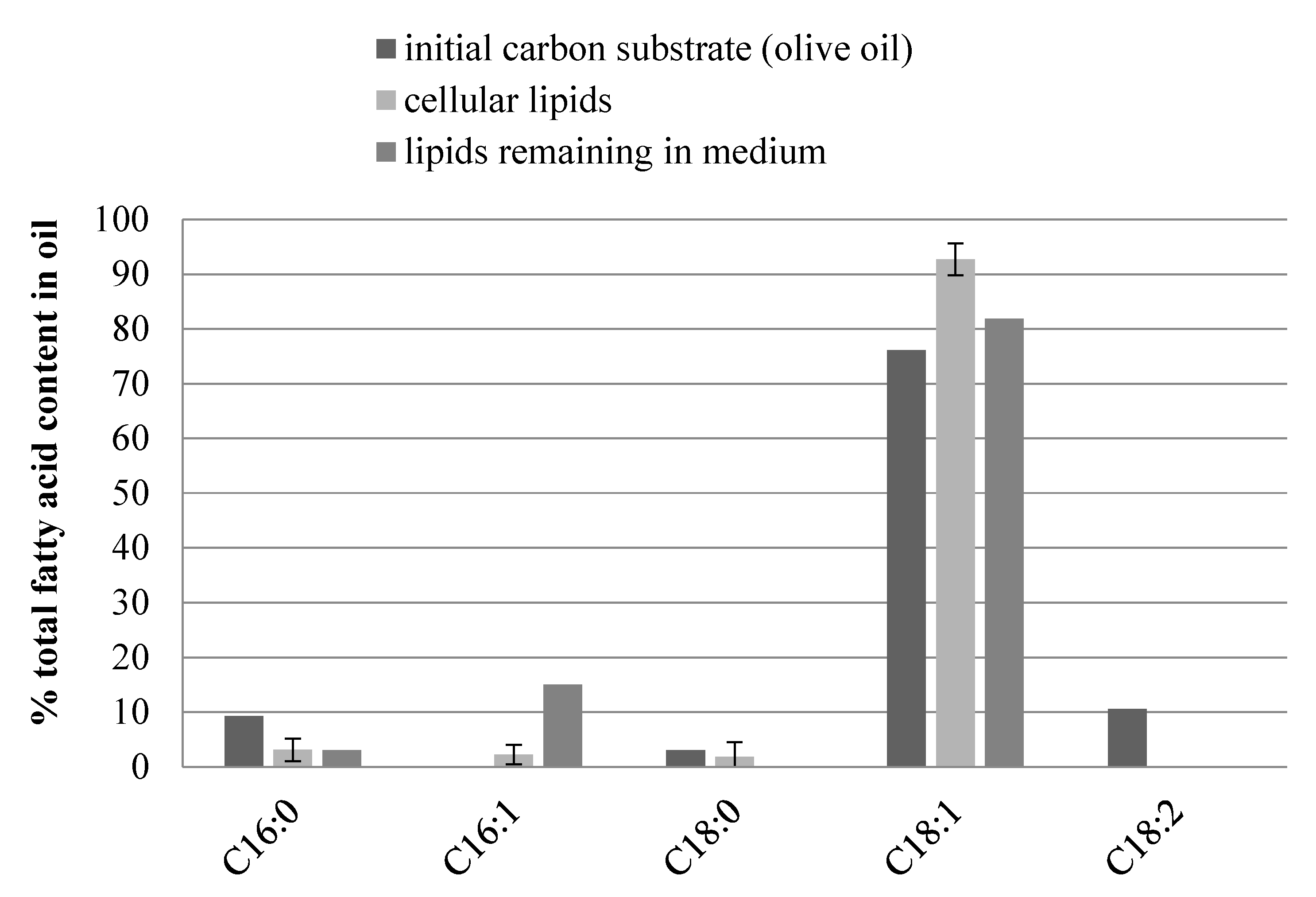
3.2. Fatty Acid Composition in Microbial Oil from Y. lipolytica
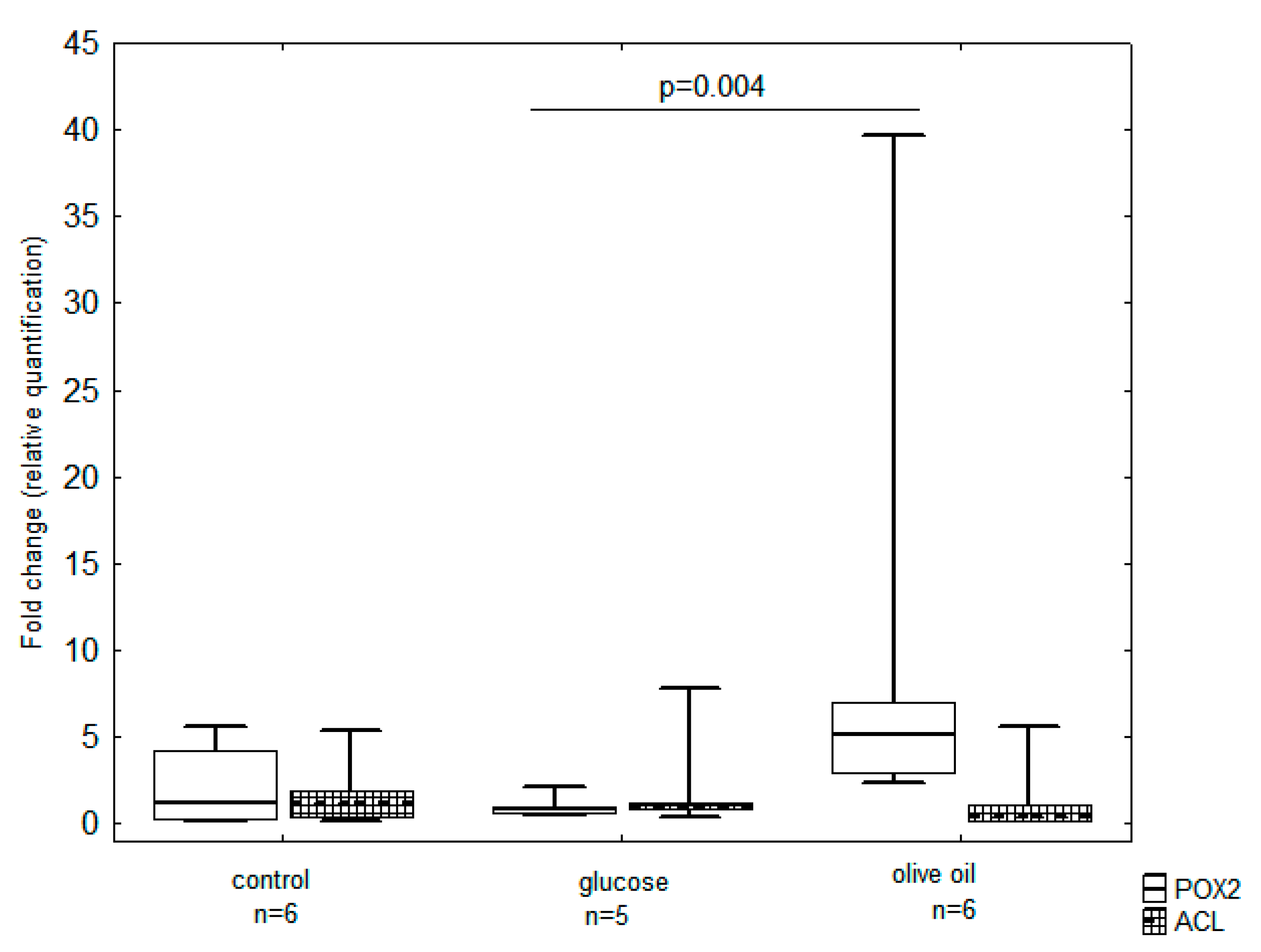
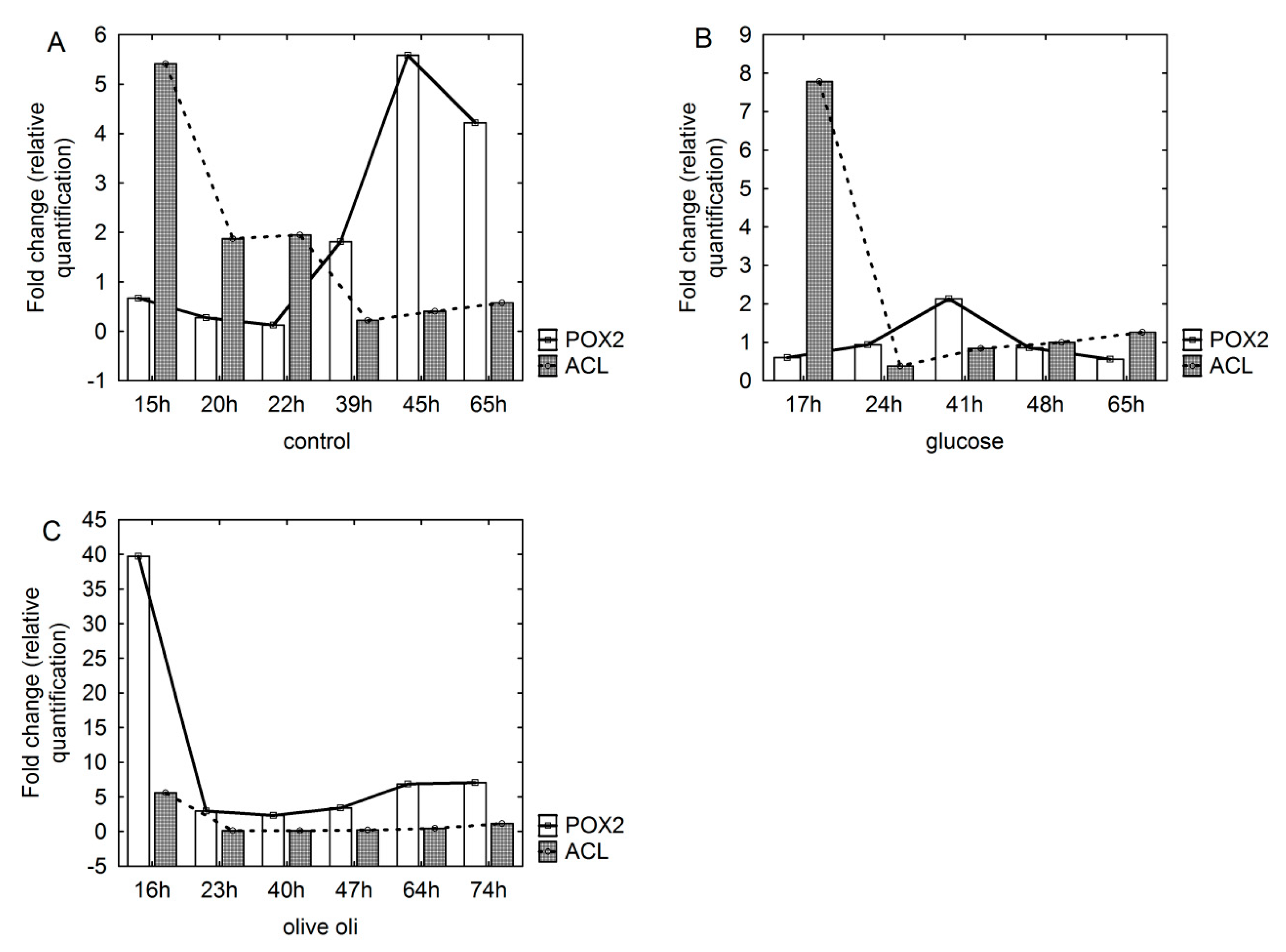
3.3. Transcription Level of Genes Involved in De Novo and Ex Novo Storage Lipid Synthesis
4. Discussion
4.1. Single Cell Oil Yield and its Quality in Dependence with Carbon Substrate
4.2. POX and ACL Genes as Markers of the Ex Novo and De Novo Routes for SCO Synthesis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.-L.; Molina-Jouve, C.; Nicaud, J.-M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Du, W.; Liu, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Papanikolau, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Clin. Chem. 2002, 51, 1–52. [Google Scholar]
- Tan, H.; Zhao, Z.K.; Tang, W.; Zhang, S.; Wang, Q. The isocitrate dehydrogenase gene of oleaginous yeast Lipomyces starkeyi is linked to lipid accumulation. Can. J. Microbiol. 2009, 55, 1062–1069. [Google Scholar]
- Wynn, J.P.; Ratledge, C.; Hamid, A.A.; Li, Y. Biochemical events leading to the diversion of carbon into storage lipids in the oleaginous fungi Mucor circinelloides and Mortierella alpina. Microbiology 2001, 147, 2857–2864. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-S.; Nicaud, J.-M.; Van Veldhoven, P.P.; Chardot, T. The acyl–CoA oxidases from the yeast Yarrowia lipolytica: Characterization of Aox2p. Arch. Biochem. Biophys. 2002, 407, 32–38. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Fabiszewska, A.; Pielińska, A.; Mazurczak, P.; Zieniuk, B.; Wołoszynowska, M. Impact of the selected factors on extraction yield and composition of fatty acids of microbial oil produced by the yeast cells of Yarrowia lipolytica. Żywność Nauka Technol. Jakość 2017, 24, 58–68. [Google Scholar]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitive PCR and the 2−ΔΔCT. Method 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Bommareddy, R.R.; Maheshwari, G.; Papanikolaou, S.; Zeng, A.-P. Substrates and oxygen dependent citric acid production by Yarrowia lipolytica: Insights through transcriptome and fluxome analyses. Microb. Cell Factories 2017, 16, 78. [Google Scholar] [CrossRef] [PubMed]
- Najjar, A.; Robert, S.; Guerin, C.; Violet-Asther, M.; Carriere, F. Quantitative study of lipase secretion, extracellular lipolysis, and lipid storage in the yeast Yarrowia lipolytica grown in the presence of olive oil: Analogies with lipolysis in humans. Appl. Microbiol. Biotechnol. 2011, 89, 1947–1962. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Cheng, S.; Zheng, L. Oleaginous yeast Yarrowia lipolytica culture with synthetic and food waste-derived volatile fatty acids for lipid production. Biotechnol. Biofuels 2017, 10, 247. [Google Scholar] [CrossRef]
- El Bialy, H.; Gomaa, O.M.; Azab, K.S. Conversion of oil waste to valuable fatty acids using Oleaginous yeast. World J. Microbiol. Biotechnol. 2011, 27, 2791–2798. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Influence of Glucose and Saturated Free-Fatty Acid Mixtures on Citric Acid and Lipid Production by Yarrowia lipolytica. Curr. Microbiology 2006, 52, 134–142. [Google Scholar] [CrossRef]
- Saygün, A.; Şahin-Yeşilçubuk, N.; Aran, N. Effects of Different Oil Sources and Residues on Biomass and Metabolite Production by Yarrowia lipolytica YB 423-12. J. Am. Oil Chem. Soc. 2014, 91, 1521–1530. [Google Scholar] [CrossRef]
- Nicaud, J.-M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, H.M.; Kalscheuer, R.; Steinbüchel, A. Accumulation of storage lipids in species of Rhodococcus and Nocardia and effect of inhibitors and polyethylene glycol. Fette. Seifen. Anstrichm. 1997, 99, 239–246. [Google Scholar]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Accumulation of a Cocoa-Butter-Like Lipid by Yarrowia lipolytica Cultivated on Agro-Industrial Residues. Curr. Microbiol. 2003, 46, 124–130. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Medium | ||
|---|---|---|---|---|
| MG7 | MO5 | YPG | ||
| Initial concentration of carbon source (S) | g/L | 70 | 50 | 20 |
| Time* (t) | h | 65 | 23 (64) | 65 |
| Biomass yield (x) | gd.w./L | 8.60 | 7.09 (10.890) | 13.53 |
| Maximum concentration of lipids produced (Lmax) | g/L | 0.994 | 1.468 (1.200) | 0.057 |
| Conversion yield of biomass per carbon substrate (YX/S) | gd.w./g | 0.123 | 0.142 (0.218) | 0.677 |
| Conversion yield of storage lipids per biomass formed (YL/X) | g/gd.w. | 0.116 | 0.207 (0.110) | 0.004 |
| Conversion yield of storage lipids per carbon substrate (YL/S) | g/g | 0.014 | 0.029 (0.024) | 0.003 |
| Volumetric rate of storage lipids production (qLv) | g/L/h | 0.015 | 0.064 (0.019) | 0.001 |
| Specific rate of storage lipid production (qL) | g/gd.w./h | 0.0018 | 0.0090 (0.0017) | 0.00006 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabiszewska, A.; Misiukiewicz-Stępień, P.; Paplińska-Goryca, M.; Zieniuk, B.; Białecka-Florjańczyk, E. An Insight into Storage Lipid Synthesis by Yarrowia lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism. Biomolecules 2019, 9, 685. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110685
Fabiszewska A, Misiukiewicz-Stępień P, Paplińska-Goryca M, Zieniuk B, Białecka-Florjańczyk E. An Insight into Storage Lipid Synthesis by Yarrowia lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism. Biomolecules. 2019; 9(11):685. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110685
Chicago/Turabian StyleFabiszewska, Agata, Paulina Misiukiewicz-Stępień, Magdalena Paplińska-Goryca, Bartłomiej Zieniuk, and Ewa Białecka-Florjańczyk. 2019. "An Insight into Storage Lipid Synthesis by Yarrowia lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism" Biomolecules 9, no. 11: 685. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110685