Identification, Classification and Screening for γ-Amino-butyric Acid Production in Lactic Acid Bacteria from Cambodian Fermented Foods

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermented Food Samples and Sampling
2.2. Growth Conditions and LAB Isolation
2.3. LAB Identification by Partial 16S rDNA Sequencing and MALDI-TOF MS
2.3.1. DNA Extraction and Identification of LAB by Partial 16S rDNA Sequencing
2.3.2. Identification of LAB by MALDI-TOF MS
2.4. Fingerprinting and Typing of LAB by (GTG)5-PCR
2.5. Screening LAB Strains for GABA Production Using Thin Layer Chromatography (TLC)
2.6. Quantification of GABA Production
2.7. Statistical Analysis
3. Results and Discussion
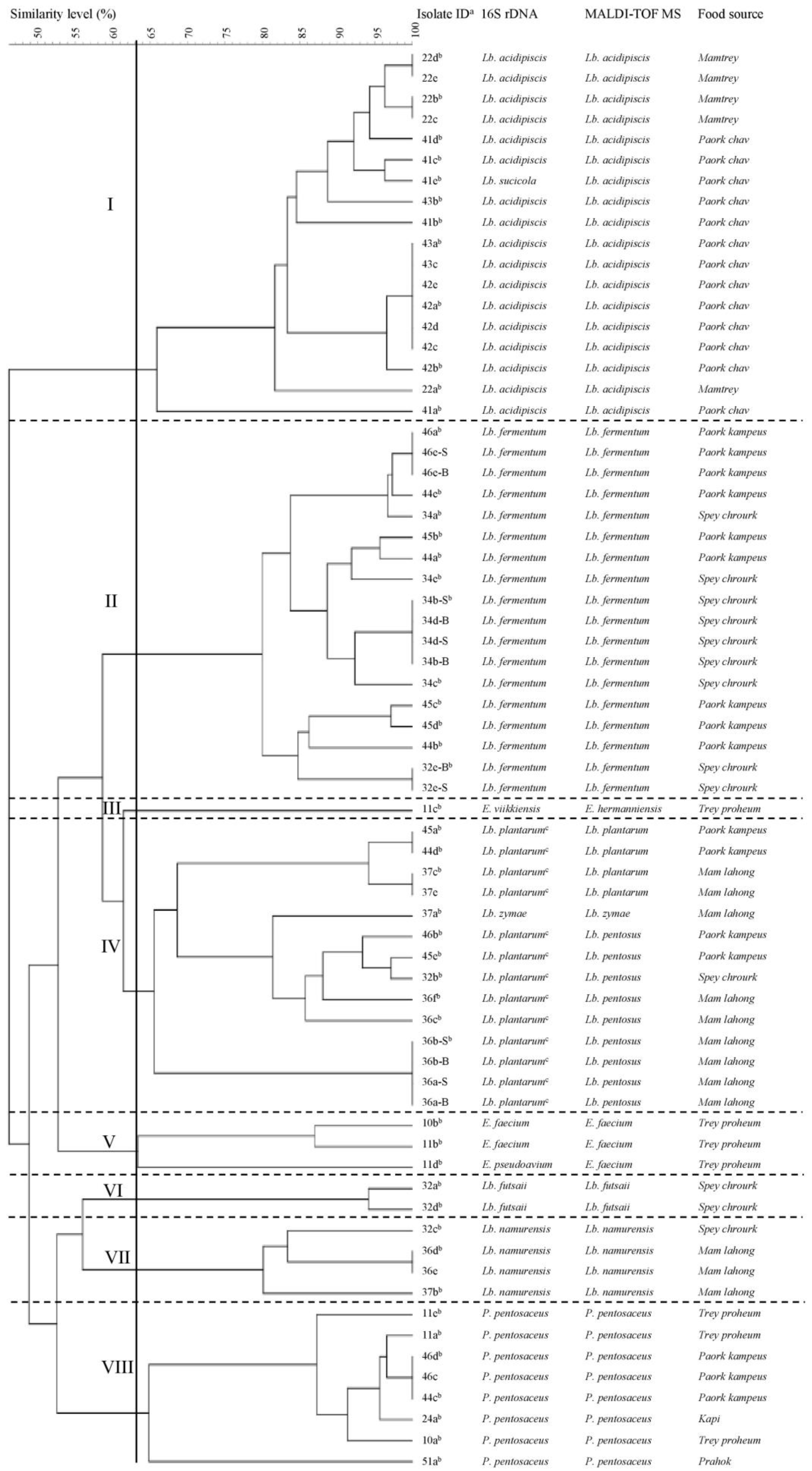
3.1. LAB Identification by Partial 16S rDNA Sequencing and MALDI-TOF MS
3.1.1. Identification of LAB by Partial 16S rDNA Sequencing
3.1.2. Identification of LAB by MALDI-TOF MS
3.2. Fingerprinting and Typing of LAB by (GTG)5-PCR
3.3. Prevalence of LAB in Cambodian Fermented Foods
3.4. Screening LAB Strains for GABA Production and GABA Quantification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ly, D.; Mayrhofer, S.; Domig, K.J. Significance of traditional fermented foods in the lower Mekong subregion: A focus on lactic acid bacteria. Food Biosci. 2018, 26, 113–125. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Koutsoumanis, K.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 8: Suitability of taxonomic units notified to EFSA until March 2018. Efsa J. 2018, 16, 1–42. [Google Scholar] [CrossRef]
- Adams, M. On the safety of lactic acid bacteria from food. Int. J. Food Microbiol. 1995, 27, 263–264. [Google Scholar] [CrossRef]
- Bojanic Rasovic, M.; Mayrhofer, S.; Martinovic, A.; Durr, K.; Domig, K.J. Lactococci of local origin as potential starter cultures for traditional Montenegrin cheese production. Food Technol. Biotechnol. 2017, 55, 55–66. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats: A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and gamma-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. Biomed. Res. Int. 2015, 2015, 625740. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Kook, M.-C.; Cho, S.-C. Production of GABA (gamma amino butyric acid) by lactic acid bacteria. Korean J. Food Sci. Anim. Resour. 2013, 33, 377–389. [Google Scholar] [CrossRef]
- Narayan, V.S.; Nair, P.M. Metabolism, enzymology and possible roles of 4-aminobutyrate in higher plants. Phytochemistry 1990, 29, 367–375. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of GABA (gamma - aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.N.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent advances in gamma-aminobutyric acid (GABA) properties in pulses: An overview. J. Sci. Food Agric. 2017, 97, 2681–2689. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Paik, H.D. Bioconversion using lactic acid bacteria: Ginsenosides, gaba, and phenolic compounds. J. Microbiol. Biotechnol. 2017, 27, 869–877. [Google Scholar] [CrossRef]
- Xu, N.; Wei, L.; Liu, J. Biotechnological advances and perspectives of gamma-aminobutyric acid production. World J. Microbiol. Biotechnol. 2017, 33, 64. [Google Scholar] [CrossRef] [PubMed]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of γ-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Dec, M.; Puchalski, A.; Urban-Chmiel, R.; Wernicki, A. 16S-ARDRA and MALDI-TOF mass spectrometry as tools for identification of Lactobacillus bacteria isolated from poultry. Bmc Microbiol. 2016, 16, 105. [Google Scholar] [CrossRef]
- Donelli, G.; Vuotto, C.; Mastromarino, P. Phenotyping and genotyping are both essential to identify and classify a probiotic microorganism. Microb. Ecol. Health Dis. 2013, 24. [Google Scholar] [CrossRef]
- Kitahara, K.; Miyazaki, K. Revisiting bacterial phylogeny: Natural and experimental evidence for horizontal gene transfer of 16S rRNA. Mob. Genet. Elem. 2013, 3, e24210. [Google Scholar] [CrossRef]
- Nguyen, D.T.L.; Van Hoorde, K.; Cnockaert, M.; Brandt, E.D.; Aerts, M.; Binh Thanh, L.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Švec, P.; Sedlacek, I.; Zackova, L.; Novakova, D.; Kukletova, M. Lactobacillus spp. associated with early childhood caries. Folia Microbiol. 2009, 54, 53–58. [Google Scholar] [CrossRef]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Sanchart, C.; Rattanaporn, O.; Haltrich, D.; Phukpattaranont, P.; Maneerat, S. Lactobacillus futsaii CS3, a new GABA-producing strain isolated from Thai fermented shrimp (Kung-Som). Indian J. Microbiol. 2017, 57, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Seo, D.H.; Cha, I.T.; Lee, H.; Nam, Y.D.; Seo, M.J. Expression and characterization of glutamate decarboxylase from Lactobacillus brevis HYE1 isolated from kimchi. World J. Microbiol. Biotechnol. 2018, 34, 44. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Quesnes, G.; Trieu-Cuot, P. Sequencing the gene encoding manganese-dependent superoxide dismutase for rapid species identification of enterococci. J. Clin. Microbiol. 2000, 38, 415–418. [Google Scholar] [PubMed]
- Devriese, L.A.; Vancanneyt, M.; Descheemaeker, P.; Baele, M.; Van Landuyt, H.W.; Gordts, B.; Butaye, P.; Swings, J.; Haesebrouck, F. Differentiation and identification of Enterococcus durans, E. hirae and E. villorum. J. Appl. Microbiol. 2002, 92, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Rahkila, R.; Johansson, P.; Sade, E.; Bjorkroth, J. Identification of enterococci from broiler products and a broiler processing plant and description of Enterococcus viikkiensis sp. nov. Appl. Environ. Microbiol. 2011, 77, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Naser, S.M.; Thompson, F.L.; Hoste, B.; Gevers, D.; Dawyndt, P.; Vancanneyt, M.; Swings, J. Application of multilocus sequence analysis (MLSA) for rapid identification of Enterococcus species based on rpoA and pheS genes. Microbiology 2005, 151, 2141–2150. [Google Scholar] [CrossRef]
- Moore, D.F.; Zhowandai, M.H.; Ferguson, D.M.; McGee, C.; Mott, J.B.; Stewart, J.C. Comparison of 16S rRNA sequencing with conventional and commercial phenotypic techniques for identification of enterococci from the marine environment. J. Appl. Microbiol. 2006, 100, 1272–1281. [Google Scholar] [CrossRef]
- Huang, C.H.; Chang, M.T.; Huang, L. Cloning of a novel specific SCAR marker for species identification in Lactobacillus pentosus. Mol. Cell. Probes 2014, 28, 192–194. [Google Scholar] [CrossRef]
- Anderson, A.C.; Sanunu, M.; Schneider, C.; Clad, A.; Karygianni, L.; Hellwig, E.; Al-Ahmad, A. Rapid species-level identification of vaginal and oral lactobacilli using MALDI-TOF MS analysis and 16S rDNA sequencing. BMC Microbiol. 2014, 14, 312. [Google Scholar] [CrossRef]
- Chavagnat, F.; Haueter, M.; Jimeno, J.; Casey, M.G. Comparison of partial tuf gene sequences for the identification of lactobacilli. Fems Microbiol. Lett. 2002, 217, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stępień-Pyśniak, D.; Hauschild, T.; Różański, P.; Marek, A. MALDI-TOF mass spectrometry as a useful tool for identification of Enterococcus spp. From wild birds and differentiation of closely related species. J. Microbiol. Biotechnol. 2017, 27, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Urban-Chmiel, R.; Gnat, S.; Puchalski, A.; Wernicki, A. Identification of Lactobacillus strains of goose origin using MALDI-TOF mass spectrometry and 16S-23S rDNA intergenic spacer PCR analysis. Res. Microbiol. 2014, 165, 190–201. [Google Scholar] [CrossRef]
- Dušková, M.; Šedo, O.; Kšicová, K.; Zdráhal, Z.; Karpíšková, R. Identification of lactobacilli isolated from food by genotypic methods and MALDI-TOF MS. Int. J. Food Microbiol. 2012, 159, 107–114. [Google Scholar] [CrossRef]
- Sato, H.; Torimura, M.; Kitahara, M.; Ohkuma, M.; Hotta, Y.; Tamura, H. Characterization of the Lactobacillus casei group based on the profiling of ribosomal proteins coded in S10-spc-alpha operons as observed by MALDI-TOF MS. Syst. Appl. Microbiol. 2012, 35, 447–454. [Google Scholar] [CrossRef]
- Kern, C.C.; Vogel, R.F.; Behr, J. Differentiation of Lactobacillus brevis strains using Matrix-Assisted-Laser-Desorption-Ionization-Time-of-Flight Mass Spectrometry with respect to their beer spoilage potential. Food Microbiol. 2014, 40, 18–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Ma, Q.; Song, Y.; Zhang, Q.; Wang, X.; Chen, F. Identification of Lactobacillus from the saliva of adult patients with caries using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. PLoS ONE 2014, 9, e106185. [Google Scholar] [CrossRef]
- Doan, N.T.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Le Thanh, B.; Vandamme, P. Validation of MALDI-TOF MS for rapid classification and identification of lactic acid bacteria, with a focus on isolates from traditional fermented foods in northern Vietnam. Lett. Appl. Microbiol. 2012, 55, 265–273. [Google Scholar] [CrossRef]
- Nacef, M.; Chevalier, M.; Chollet, S.; Drider, D.; Flahaut, C. MALDI-TOF mass spectrometry for the identification of lactic acid bacteria isolated from a French cheese: The Maroilles. Int. J. Food Microbiol. 2017, 247, 2–8. [Google Scholar] [CrossRef]
- Foschi, C.; Laghi, L.; Parolin, C.; Giordani, B.; Compri, M.; Cevenini, R.; Marangoni, A.; Vitali, B. Novel approaches for the taxonomic and metabolic characterization of lactobacilli: Integration of 16S rRNA gene sequencing with MALDI-TOF MS and 1H-NMR. PLoS ONE 2017, 12, e0172483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, E.F.; Luciano, W.A.; Xavier, D.E.; da Costa, W.C.; de Sousa Oliveira, K.; Franco, O.L.; de Morais Junior, M.A.; Lucena, B.T.; Picao, R.C.; Magnani, M.; et al. Identification of lactic acid bacteria in fruit pulp processing byproducts and potential probiotic properties of selected Lactobacillus strains. Front. Microbiol. 2016, 7, 1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Kim, E.; Lee, Y.; Han, S.K.; Kim, C.G.; Choo, D.W.; Kim, Y.R.; Kim, H.Y. Rapid and accurate identification of species of the genus Pediococcus isolated from Korean fermented foods by matrix-assisted laser desorption/ionization time-of-flight MS with local database extension. Int. J. Syst. Evol. Microbiol. 2017, 67, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Alatoom, A.A.; Cunningham, S.A.; Ihde, S.M.; Mandrekar, J.; Patel, R. Comparison of direct colony method versus extraction method for identification of gram-positive cocci by use of Bruker Biotyper matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2011, 49, 2868–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, N.; Matsuda, M.; Notake, S.; Yokokawa, H.; Kawamura, Y.; Hiramatsu, K.; Kikuchi, K. Evaluation of a simple protein extraction method for species identification of clinically relevant staphylococci by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2012, 50, 3862–3866. [Google Scholar] [CrossRef] [Green Version]
- Sanchart, C.; Rattanaporn, O.; Haltrich, D.; Phukpattaranont, P.; Maneerat, S. Technological and safety properties of newly isolated GABA-producing Lactobacillus futsaii strains. J. Appl. Microbiol. 2016, 121, 734–745. [Google Scholar] [CrossRef]
- Švec, P.; Sedlacek, I.; Chrapava, M.; Vandamme, P. (GTG)5-PCR fingerprinting of lactobacilli isolated from cervix of healthy women. Folia Microbiol. 2011, 56, 80–83. [Google Scholar] [CrossRef]
- Gevers, D.; Huys, G.; Swings, J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. Fems Microbiol. Lett. 2001, 205, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H.; Wood, B.J.B. (Eds.) The genus Lactobacillus. In Lactic Acid Bacteria: Biodiversity and Taxonomy; Wiley Blackwell: Chennai, India, 2014; pp. 249–353. [Google Scholar]
- Kazou, M.; Alexandraki, V.; Pot, B.; Tsakalidou, E.; Papadimitriou, K. Complete genome sequence of the sourdough isolate Lactobacillus zymae ACA-DC 3411. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.G.; Ye, Z.Q.; Yu, L.; Shi, P. Phylogenomic reconstruction of lactic acid bacteria: An update. Bmc Evol. Biol. 2011, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; van Sinderen, D.; O’Toole, P.W. Lactobacillus phylogenomics--towards a reclassification of the genus. Int. J. Syst. Evol. Microbiol. 2008, 58, 2945–2954. [Google Scholar] [CrossRef] [PubMed]
- Tanasupawat, S.; Shida, O.; Okada, S.; Komagata, K. Lactobacillus acidipiscis sp. nov. and Weissella thailandensis sp. nov., isolated from fermented fish in Thailand. Int. J. Syst. Evol. Microbiol. 2000, 50(Pt. 4), 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Luo, J.; Li, P.; Yu, H.; Huang, J.; Luo, L. Microbial diversity and flavor formation in onion fermentation. Food Funct. 2014, 5, 2338–2347. [Google Scholar] [CrossRef] [PubMed]
- Kopermsub, P.; Yunchalard, S. Identification of lactic acid bacteria associated with the production of plaa-som, a traditional fermented fish product of Thailand. Int. J. Food Microbiol. 2010, 138, 200–204. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Balcázar, J.L.; Daniels, C.; Zhou, Z.; Carnevali, O.; Sun, Y.Z.; Hoseinifar, S.H.; Ringø, E. Indigenous lactic acid bacteria in fish and crustaceans. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Merrifield, D., Ringø, E., Eds.; Wiley-Blackwell: Oxford, UK, 2014; pp. 128–168. [Google Scholar] [CrossRef]
- Bucio, A.; Hartemink, R.; Schrama, J.W.; Verreth, J.; Rombouts, F.M. Presence of lactobacilli in the intestinal content of freshwater fish from a river and from a farm with a recirculation system. Food Microbiol. 2006, 23, 476–482. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozes, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef]
- Scheirlinck, I.; Van der Meulen, R.; Van Schoor, A.; Cleenwerck, I.; Huys, G.; Vandamme, P.; De Vuyst, L.; Vancanneyt, M. Lactobacillus namurensis sp. nov., isolated from a traditional Belgian sourdough. Int. J. Syst. Evol. Microbiol. 2007, 57, 223–227. [Google Scholar] [CrossRef]
- George, F.; Daniel, C.; Thomas, M.; Singer, E.; Guilbaud, A.; Tessier, F.J.; Revol-Junelles, A.M.; Borges, F.; Foligne, B. Occurrence and dynamism of lactic acid bacteria in distinct ecological niches: A multifaceted functional health perspective. Front. Microbiol. 2018, 9, 2899. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Toh, H.; Sakamoto, N.; Mori, K.; Tashiro, K.; Hibi, N.; Sonomoto, K.; Nakayama, J. Draft genome sequence of Lactobacillus namurensis Chizuka 01, isolated from Nukadoko, a pickling bed of fermented rice bran. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Jeong, S.J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) gene from Lactobacillus zymae. Biotechnol. Lett. 2014, 36, 1791–1799. [Google Scholar] [CrossRef]
- Chao, S.H.; Kudo, Y.; Tsai, Y.C.; Watanabe, K. Lactobacillus futsaii sp. nov., isolated from fu-tsai and suan-tsai, traditional Taiwanese fermented mustard products. Int. J. Syst. Evol. Microbiol. 2012, 62, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Cho, Y.; Kim, E.; Kim, H.J.; Kim, H.Y. Identification of lactic acid bacteria in Galchi- and Myeolchi-Jeotgal by 16S rRNA gene sequencing, MALDI-TOF mass spectrometry, and PCR-DGGE. J. Microbiol. Biotechnol. 2018, 28, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Park, J.Y.; Sa, H.D.; Jeong, J.H.; Jin, D.E.; Heo, H.J.; Kim, J.H. Probiotic properties of Pediococcus strains isolated from jeotgals, salted and fermented Korean seafood. Anaerobe 2014, 28, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Simpson, W.J.; Taguchi, H. The genus Pediococcus, with notes on the genera Tetratogenococcus and Aerococcus. In The Genera of Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer: Boston, MA, USA, 1995; pp. 125–172. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koort, J.; Coenye, T.; Vandamme, P.; Sukura, A.; Bjorkroth, J. Enterococcus hermanniensis sp. nov., from modified-atmosphere-packaged broiler meat and canine tonsils. Int. J. Syst. Evol. Microbiol. 2004, 54, 1823–1827. [Google Scholar] [CrossRef]
- Devi, K.R.; Deka, M.; Jeyaram, K. Bacterial dynamics during yearlong spontaneous fermentation for production of ngari, a dry fermented fish product of Northeast India. Int. J. Food Microbiol. 2015, 199, 62–71. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, S.; Lee, R.S.; Lim, S.D.; Kim, H.J.; Lee, M.K. Selection and characteristics of fermented salted seafood (jeotgal)-originated strains with excellent S-adenosyl-L-methionine (SAM) production and probiotics efficacy. Korean J. Food Sci. Anim. Resour. 2014, 34, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Kim, K.-S. Isolation and identification of γ-aminobutyric acid (GABA)-producing lactic acid bacteria from Kimchi. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 777–785. [Google Scholar] [CrossRef]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of gamma-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [Green Version]
- Kantachote, D.; Ratanaburee, A.; Sukhoom, A.; Sumpradit, T.; Asavaroungpipop, N. Use of γ-aminobutyric acid producing lactic acid bacteria as starters to reduce biogenic amines and cholesterol in Thai fermented pork sausage (nham) and their distribution during fermentation. Lwt-Food Sci. Technol. 2016, 70, 171–177. [Google Scholar] [CrossRef]
- Ratanaburee, A.; Kantachote, D.; Charernjiratrakul, W.; Sukhoom, A. Selection of γ-aminobutyric acid-producing lactic acid bacteria and their potential as probiotics for use as starter cultures in Thai fermented sausages (Nham). Int. J. Food Sci. Tech. 2013, 48, 1371–1382. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Woraharn, S.; Sivamaruthi, B.S.; Lailerd, N.; Kesika, P.; Peerajan, S. Lactobacillus fermentum HP3-Mediated fermented Hericium erinaceus juice as a health promoting food supplement to manage diabetes mellitus. J. Evid. Based Integr. Med. 2018, 23, 2515690X18765699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carafa, I.; Nardin, T.; Larcher, R.; Viola, R.; Tuohy, K.; Franciosi, E. Identification and characterization of wild lactobacilli and pediococci from spontaneously fermented Mountain cheese. Food Microbiol. 2015, 48, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Cha, I.-T.; Lee, H.; Seo, M.-J. Optimization of γ-aminobutyric acid production by Enterococcus faecium JK29 isolated from a traditional fermented foods. Microbiol. Biotechnol. Lett. 2016, 44, 26–33. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Yao, Z.; Kim, J.A.; Kim, H.J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) from Enterococcus avium M5 isolated from Jeotgal, a Korean fermented seafood. J. Microbiol. Biotechnol. 2017, 27, 1216–1222. [Google Scholar] [CrossRef]
- Shan, Y.; Man, C.X.; Han, X.; Li, L.; Guo, Y.; Deng, Y.; Li, T.; Zhang, L.W.; Jiang, Y.J. Evaluation of improved gamma-aminobutyric acid production in yogurt using Lactobacillus plantarum NDC75017. J. Dairy Sci. 2015, 98, 2138–2149. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Shah, N.P. High gamma-aminobutyric acid production from lactic acid bacteria: Emphasis on Lactobacillus brevis as a functional dairy starter. Crit. Rev. Food Sci. Nutr. 2016, 57, 3661–3672. [Google Scholar] [CrossRef] [Green Version]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary neurotransmitters: A narrative review on current knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef] [Green Version]
- Gan, R.-Y.; Lui, W.-Y.; Wu, K.; Chan, C.-L.; Dai, S.-H.; Sui, Z.-Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Akihiro, T.; Koike, S.; Tani, R.; Tominaga, T.; Watanabe, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Ashihara, H.; Matsukura, C.; et al. Biochemical mechanism on GABA accumulation during fruit development in tomato. Plant Cell Physiol. 2008, 49, 1378–1389. [Google Scholar] [CrossRef] [Green Version]
- Perucho, J.; Gonzalo-Gobernado, R.; Bazan, E.; Casarejos, M.J.; Jimenez-Escrig, A.; Asensio, M.J.; Herranz, A.S. Optimal excitation and emission wavelengths to analyze amino acids and optimize neurotransmitters quantification using precolumn OPA-derivatization by HPLC. Amino Acids 2015, 47, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.A.; Florez, A.B.; Vazquez, L.; Vasek, O.M.; Mayo, B. Production of gamma-aminobutyric acid (GABA) by lactic acid bacteria strains isolated from traditional, starter-free dairy products made of raw milk. Benef. Microbes 2019, 10, 579–587. [Google Scholar] [CrossRef]
- Sanchart, C.; Rattanaporn, O.; Haltrich, D.; Phukpattaranont, P.; Maneerat, S. Enhancement of gamma-aminobutyric acid (GABA) levels using an autochthonous Lactobacillus futsaii CS3 as starter culture in Thai fermented shrimp (Kung-Som). World J. Microbiol. Biotechnol. 2017, 33, 152. [Google Scholar] [CrossRef]


{kind=link}
| No. | Local Name (Number of Samples) | English Name | Ingredients | Usage | Market Origin |
|---|---|---|---|---|---|
| 1 | Prahok (n = 1) | Fish paste | Freshwater fish, salt | Main dish, side-dish, condiment, seasoning | Chamkadaung |
| 2 | Paork chav (n = 3) | Fermented fish | Freshwater fish, brown glutinous rice, salt | Main dish, side-dish | Oreusey |
| 3 | Mam trey (n = 1) | Fermented fish | Freshwater fish, palm sugar, salt | Main dish, side-dish | Thmey |
| 4 | Trey proheum (n = 2) | Salted fish | Freshwater fish, salt | Main dish, seasoning | Thmey |
| 5 | Kapi (n = 1) | Shrimp paste | Tiny marine shrimp, salt | Side-dish, condiment, seasoning | Chas |
| 6 | Paork kampeus (n = 3) | Fermented tiny freshwater shrimp | Tiny freshwater shrimp, salt, roasted rice, slightly green papaya, galangal | Side-dish | Phumreusey, Limcheanghak |
| 7 | Mam lahong (n = 2) | Fermented green papaya | Green papaya, slightly tiny fermented fish, salt, roasted rice, galangal | Side-dish | Limcheanghak |
| 8 | Spey chrourk (n = 2) | Fermented mustard | Chinese mustard, salt | Side-dish | Phumreusey, Limcheanghak |
| Food Samples | Total Number of Isolates | Identification Technique | |||
|---|---|---|---|---|---|
| Partial 16S rDNA | Number of Identified Isolates | MALDI-TOF MS (Bruker Biotyper) | Number of Identified Isolates | ||
| Prahok (n = 1) | 1 | P. pentosaceus | 1 | P. pentosaceus | 1 |
| Paork chav (n = 3) | 13 | Lb. acidipiscis Lb. sucicola | 12 1 | Lb. acidipiscis – | 13 0 |
| Mam trey (n = 1) | 5 | Lb. acidipiscis | 5 | Lb. acidipiscis | 5 |
| Trey proheum (n = 2) | 7 | P. pentosaceus E. faecium – E. viikkiensis/E. durans/E. malodoratus/E. pseudoavium E. pseudoavium/E. avium | 3 2 0 1 1 | P. pentosaceus E. faecium E. hermanniensis – – | 3 3 1 0 0 |
| Kapi (n = 1) | 1 | P. pentosaceus | 1 | P. pentosaceus | 1 |
| Paork kampeus (n = 3) | 16 | Lb. fermentum Lb. plantarum/Lb. paraplantarum/Lb. pentosus P. pentosaceus | 9 4 3 | Lb. fermentum Lb. pentosus Lb. plantarum P. pentosaceus | 9 2 2 3 |
| Mam lahong (n = 2) | 12 | Lb. plantarum/Lb. paraplantarum/Lb. pentosus Lb. namurensis Lb. zymae | 8 3 1 | Lb. pentosus Lb. plantarum Lb. namurensis Lb. zymae | 6 2 3 1 |
| Spey chrourk (n = 2) | 13 | Lb. fermentum Lb. plantarum/Lb. paraplantarum/Lb. pentosus Lb. namurensis Lb. futsaii | 9 1 1 2 | Lb. fermentum Lb. pentosus Lb. namurensis Lb. futsaii | 9 1 1 2 |
| Strain | LAB Species Identified by MALDI-TOF MS (Bruker Biotyper) | Corresponding Lanes of TLC Analysis | Rf (cm) a | GABA (mM) b Production |
|---|---|---|---|---|
| 45a | Lb. plantarum |  | 0.27 | 20.34 ± 1.41 |
| 44d | Lb. plantarum |  | 0.27 | 16.47 ± 1.91 |
| 37e | Lb. plantarum |  | 0.27 | 5.63 ± 0.68 |
| 32d | Lb. futsaii |  | 0.27 | 4.68 ± 0.87 |
| 37b | Lb. namurensis |  | 0.27 | 1.62 ± 0.43 |
| 32c | Lb. namurensis |  | 0.27 | 1.19 ± 0.66 |
| - | GABA-negative strain c |  | 0.16 | n.d. |
| - | GABA standard |  | 0.27 | n.d. |
| - | MSG d |  | 0.16 | n.d. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ly, D.; Mayrhofer, S.; Agung Yogeswara, I.B.; Nguyen, T.-H.; Domig, K.J. Identification, Classification and Screening for γ-Amino-butyric Acid Production in Lactic Acid Bacteria from Cambodian Fermented Foods. Biomolecules 2019, 9, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120768
Ly D, Mayrhofer S, Agung Yogeswara IB, Nguyen T-H, Domig KJ. Identification, Classification and Screening for γ-Amino-butyric Acid Production in Lactic Acid Bacteria from Cambodian Fermented Foods. Biomolecules. 2019; 9(12):768. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120768
Chicago/Turabian StyleLy, Dalin, Sigrid Mayrhofer, I. B. Agung Yogeswara, Thu-Ha Nguyen, and Konrad J. Domig. 2019. "Identification, Classification and Screening for γ-Amino-butyric Acid Production in Lactic Acid Bacteria from Cambodian Fermented Foods" Biomolecules 9, no. 12: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120768