Quantification of Fecal Short Chain Fatty Acids by Liquid Chromatography Tandem Mass Spectrometry—Investigation of Pre-Analytic Stability

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of Human Fecal and Murine Colon Content Homogenates
2.3. Sample Preparation
2.4. Calibrators and Quality Controls
2.5. LC-MS/MS
2.6. Method Validation
2.7. Investigation of Pre-Analytic Stability
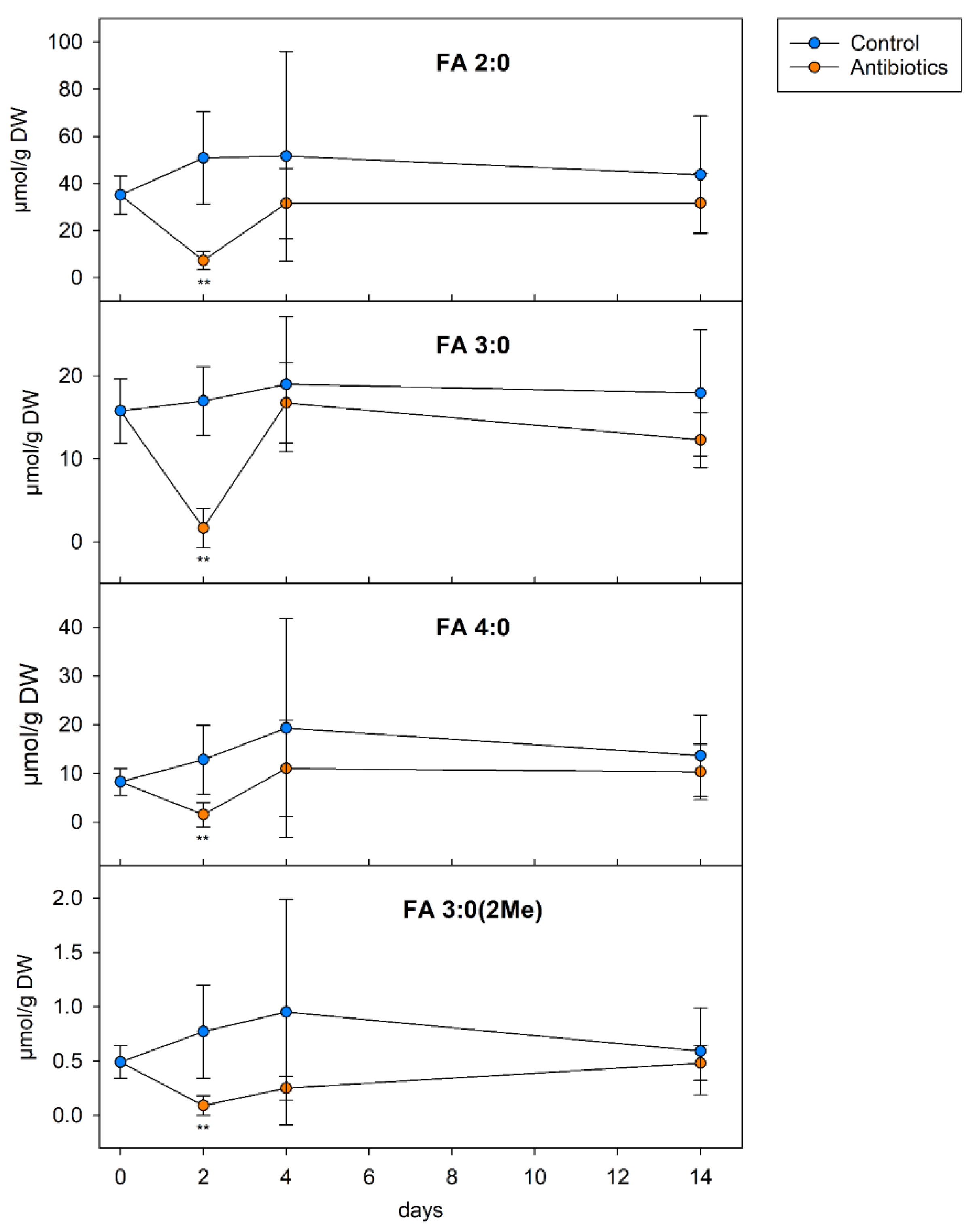
2.8. Mouse Experiments
2.9. Statistical Analyses
3. Results
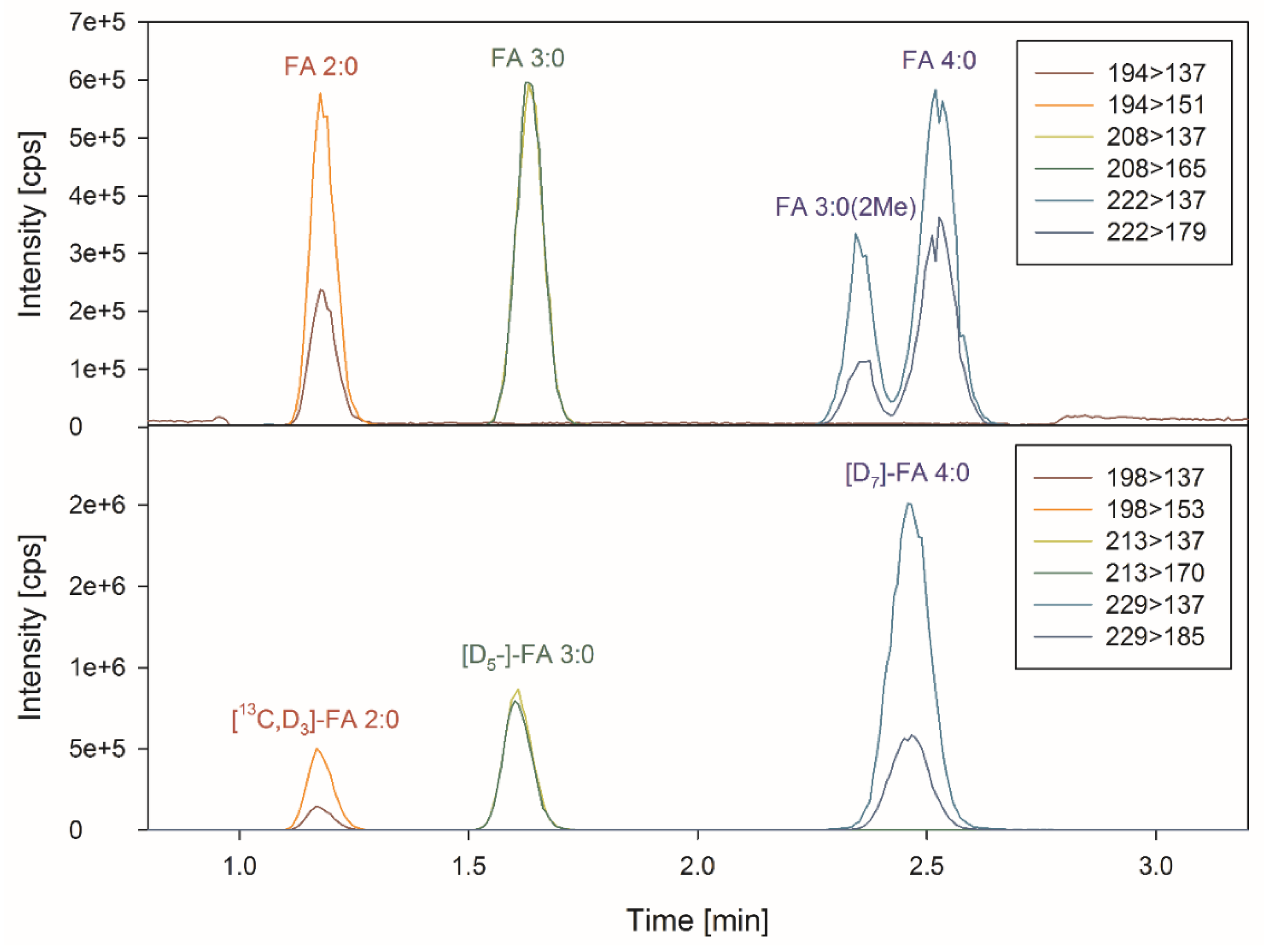
3.1. Sample Preparation and Chromatography
3.2. Specificity and Matrix Effects
3.3. Calibration
3.4. Limit of Detection (LoD) and Reproducibility
3.5. SCFA Concentrations in Human Feces
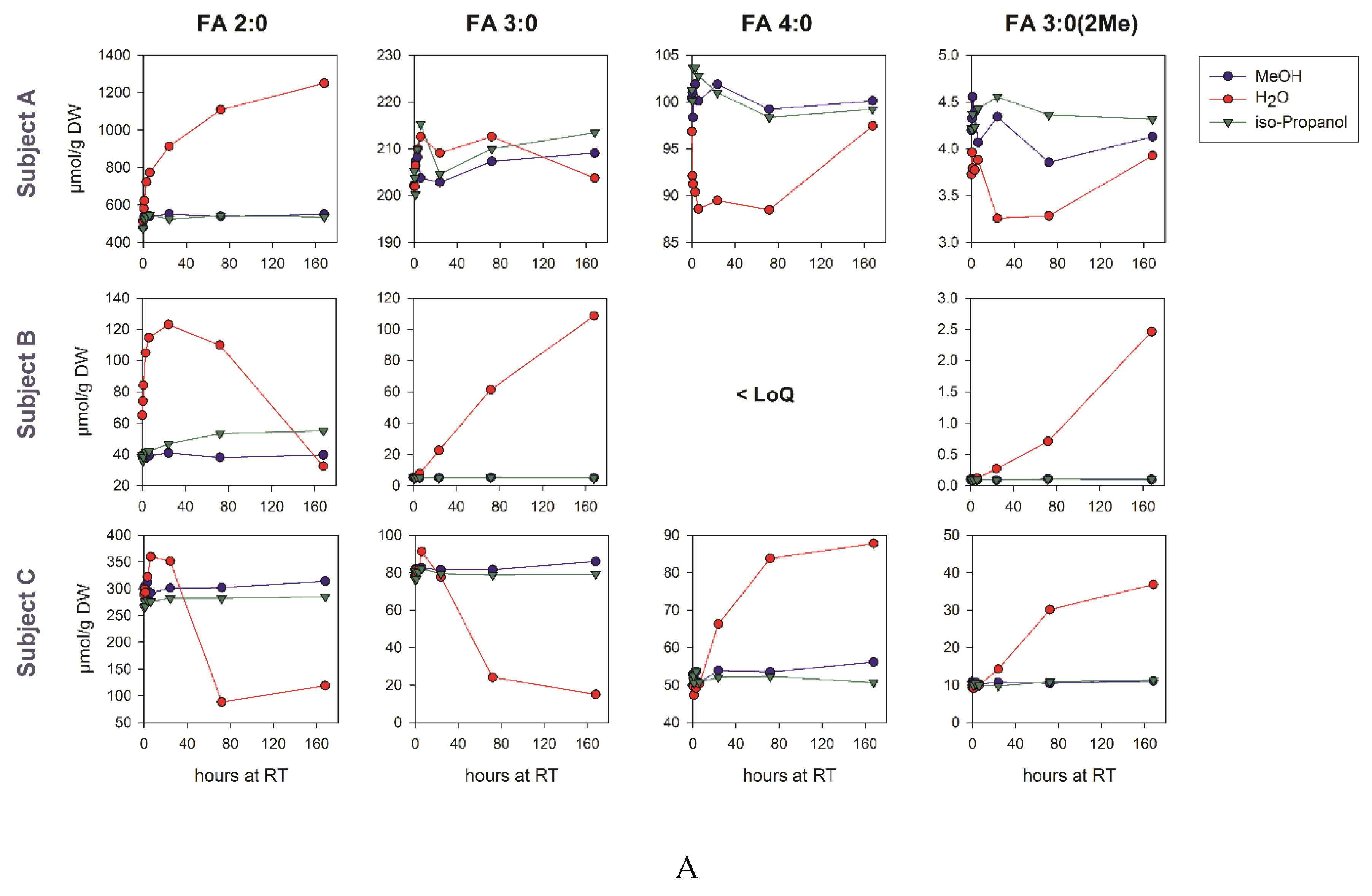
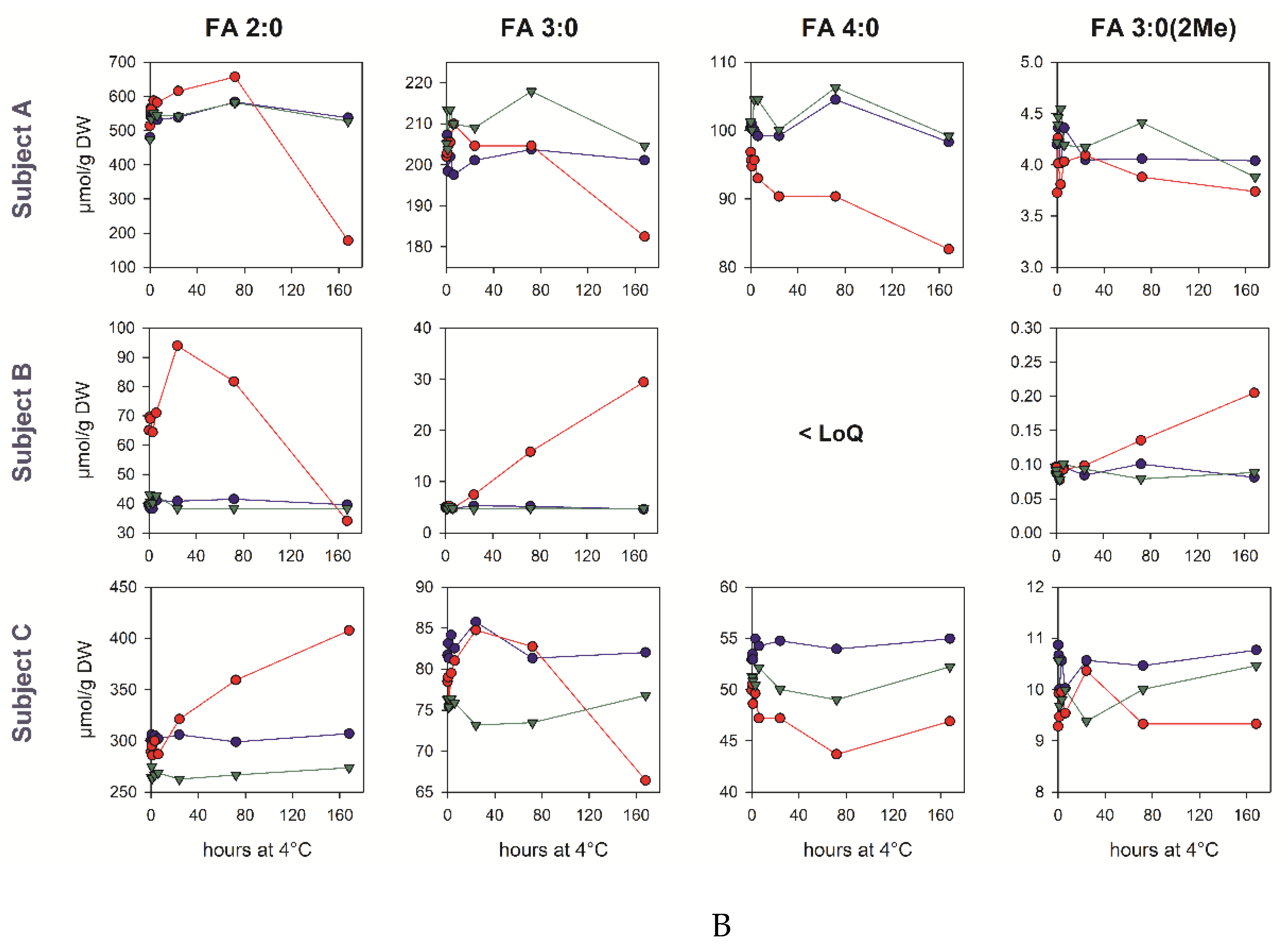
3.6. Pre-Analytic Stability in Human Fecal Samples
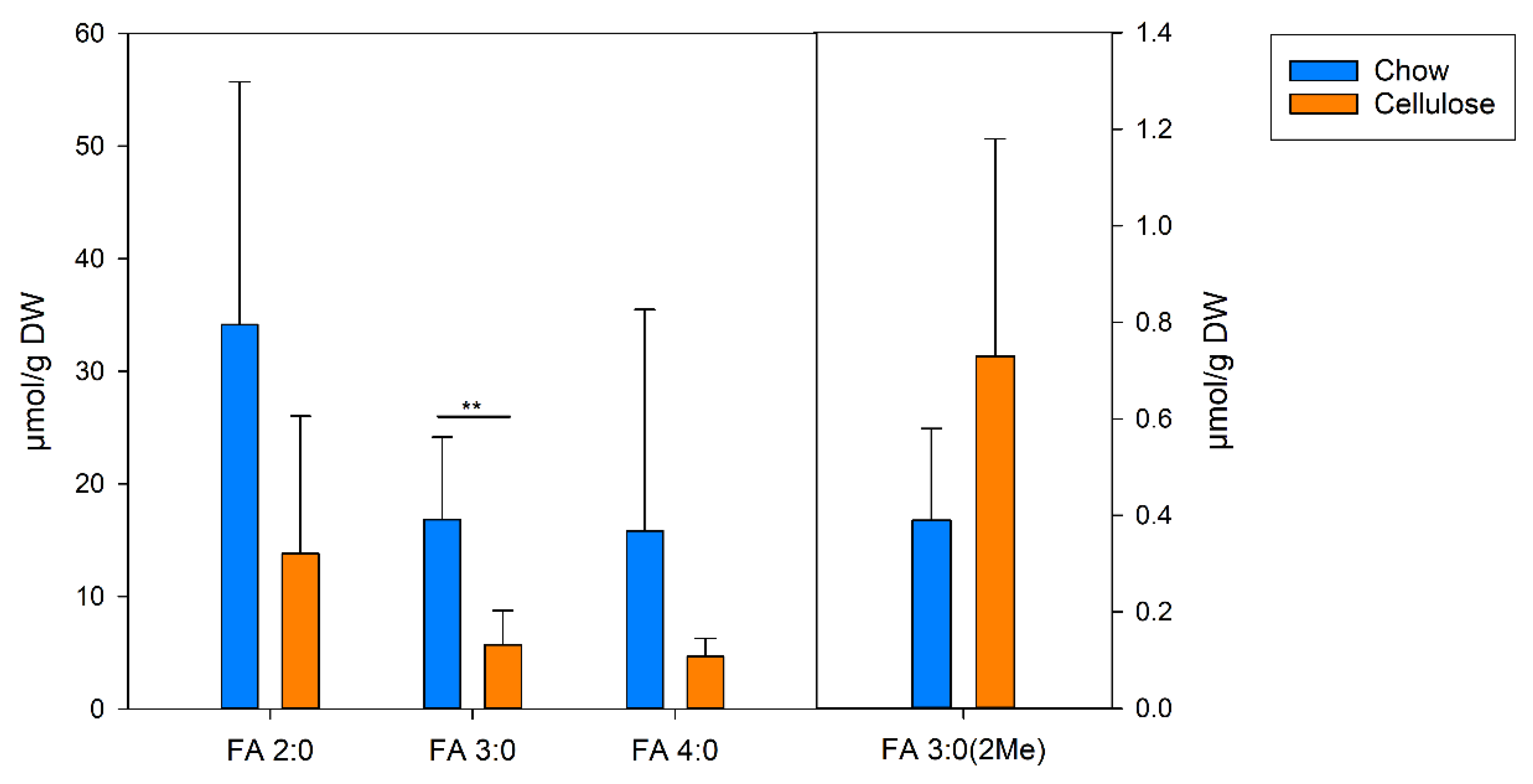
3.7. SCFA in the Colon Content of Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liebisch, G.; Vizcaino, J.A.; Koefeler, H.; Troetzmueller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Cani, P.D.; Van Hul, M.; Lefort, C.; Depommier, C.; Rastelli, M.; Everard, A. Microbial regulation of organismal energy homeostasis. Nat. Metab. 2019, 1, 34–46. [Google Scholar] [CrossRef]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 1. [Google Scholar] [CrossRef] [PubMed]
- Kindt, A.; Liebisch, G.; Clavel, T.; Haller, D.; Hoermannsperger, G.; Yoon, H.; Kolmeder, D.; Sigruener, A.; Krautbauer, S.; Seeliger, C.; et al. The gut microbiota promotes hepatic fatty acid desaturation and elongation in mice. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primec, M.; Micetic-Turk, D.; Langerholc, T. Analysis of short-chain fatty acids in human feces: A scoping review. Anal. Biochem. 2017, 526, 9–21. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, Y.; Shrestha, R.; Gao, Z.; Zhao, Y.; Miura, Y.; Tamakoshi, A.; Chiba, H.; Hui, S.P. Determination of Total, Free, and Esterified Short-Chain Fatty Acid in Human Serum by LC-MS/MS. Ann. Clin. Biochem. 2018, 56, 190–197. [Google Scholar] [CrossRef]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An isotope-labeled chemical derivatization method for the quantitation of short-chain fatty acids in human feces by liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Cao, H. Fast quantification of short chain fatty acids and ketone bodies by liquid chromatography-tandem mass spectrometry after facile derivatization coupled with liquid-liquid extraction. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1083, 137–145. [Google Scholar] [CrossRef]
- Chan, J.C.; Kioh, D.Y.; Yap, G.C.; Lee, B.W.; Chan, E.C. A novel LCMSMS method for quantitative measurement of short-chain fatty acids in human stool derivatized with (12)C- and (13)C-labelled aniline. J. Pharm. Biomed. Anal. 2017, 138, 43–53. [Google Scholar] [CrossRef]
- Schott, H.-F.; Krautbauer, S.; Hoering, M.; Liebisch, G.; Matysik, S. A Validated, Fast Method for Quantification of Sterols and Gut Microbiome Derived 5 alpha/beta-Stanols in Human Feces by Isotope Dilution LC-High-Resolution MS. Anal. Chem. 2018, 90, 8487–8494. [Google Scholar] [CrossRef] [PubMed]
- Booth, B.; Arnold, M.E.; DeSilva, B.; Amaravadi, L.; Dudal, S.; Fluhler, E.; Gorovits, B.; Haidar, S.H.; Kadavil, J.; Lowes, S.; et al. Workshop report: Crystal City V—Quantitative bioanalytical method validation and implementation: The 2013 revised FDA guidance. AAPS J. 2015, 17, 277–288. [Google Scholar] [CrossRef]
- European Medicines Agency. Committee for Medicinal Products for Human, U. Guideline on Bioanalytical Method Validation; European Medicines Agency: London, UK, 2014. [Google Scholar]
- Armbruster, D.A.; Pry, T. Limit of blank, limit of detection and limit of quantitation. Clin. Biochem. Rev. 2008, 29 (Suppl. 1), S49–S52. [Google Scholar]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krautbauer, S.; Buechler, C.; Liebisch, G. Relevance in the Use of Appropriate Internal Standards for Accurate Quantification Using LC-MS/MS: Tauro-Conjugated Bile Acids as an Example. Anal. Chem. 2016, 88, 10957–10961. [Google Scholar] [CrossRef] [PubMed]
- Ecker, J.; Liebisch, G. Application of stable isotopes to investigate the metabolism of fatty acids, glycerophospholipid and sphingolipid species. Prog. Lipid Res. 2014, 54, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Ecker, J.; Liebisch, G.; Englmaier, M.; Grandl, M.; Robenek, H.; Schmitz, G. Induction of fatty acid synthesis is a key requirement for phagocytic differentiation of human monocytes. Proc. Natl. Acad. Sci. USA 2010, 107, 7817–7822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, S.F.; Hermansson, M.; Liebisch, G.; Hodson, L.; Ejsing, C.S. Total Fatty Acid Analysis of Human Blood Samples in One Minute by High-Resolution Mass Spectrometry. Biomolecules 2018, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Wenzl, H.H.; Fine, K.D.; Schiller, L.R.; Fordtran, J.S. Determinants of decreased fecal consistency in patients with diarrhea. Gastroenterology 1995, 108, 1729–1738. [Google Scholar] [CrossRef]
- Fischer, B.; Hoh, S.; Wehler, M.; Hahn, E.G.; Schneider, H.T. Faecal elastase-1: Lyophilization of stool samples prevents false low results in diarrhoea. Scand. J. Gastroenterol. 2001, 36, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The Characterization of Feces and Urine: A Review of the Literature to Inform Advanced Treatment Technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef] [Green Version]
- Hoverstad, T.; Fausa, O.; Bjorneklett, A.; Bohmer, T. Short-chain fatty acids in the normal human feces. Scand. J. Gastroenterol. 1984, 19, 375–381. [Google Scholar] [CrossRef]
- Gardana, C.; Del Bo, C.; Simonetti, P. Validation and application of an ultrahigh-performance liquid chromatographic-Orbitrap mass spectrometric method for the simultaneous detection and quantification of volatile and non-volatile organic acids in human faecal samples. J. Pharm. Biomed. Anal. 2017, 141, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalba, R.; Gimenez-Bastida, J.A.; Garcia-Conesa, M.T.; Tomas-Barberan, F.A.; Carlos Espin, J.; Larrosa, M. Alternative method for gas chromatography-mass spectrometry analysis of short-chain fatty acids in faecal samples. J. Sep. Sci. 2012, 35, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Burla, B.; Arita, M.; Arita, M.; Bendt, A.K.; Cazenave-Gassiot, A.; Dennis, E.A.; Ekroos, K.; Han, X.; Ikeda, K.; Liebisch, G.; et al. MS-based lipidomics of human blood plasma: A community-initiated position paper to develop accepted guidelines. J. Lipid Res. 2018, 59, 2001–2017. [Google Scholar] [CrossRef]
- Matysik, S.; Le Roy, C.I.; Liebisch, G.; Claus, S.P. Metabolomics of fecal samples: A practical consideration. Trends Food Sci. Technol. 2016, 57, 244–255. [Google Scholar] [CrossRef]
- Gratton, J.; Phetcharaburanin, J.; Mullish, B.H.; Williams, H.R.; Thursz, M.; Nicholson, J.K.; Holmes, E.; Marchesi, J.R.; Li, J.V. Optimized Sample Handling Strategy for Metabolic Profiling of Human Feces. Anal. Chem. 2016, 88, 4661–4668. [Google Scholar] [CrossRef]
- Torii, T.; Kanemitsu, K.; Wada, T.; Itoh, S.; Kinugawa, K.; Hagiwara, A. Measurement of short-chain fatty acids in human faeces using high-performance liquid chromatography: Specimen stability. Ann. Clin. Biochem. 2010, 47, 447–452. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyt | RT (min) | Quantifier (m/z) | Qualifier (m/z) | DP (V) | CE (V) | LoD (µmol/g DW) | Highest Calibrator (µmol/g DW) |
|---|---|---|---|---|---|---|---|
| FA 2:0 | 1.18 | 194.1 > 151.1 | 194.1 > 137.1 | −50 | −17 | 1.9 | 1020 |
| [13C,D3]-FA 2:0 | 1.17 | 198.1 > 153.1 | 198.1 > 137.1 | −50 | −17 | - | - |
| FA 3:0 | 1.63 | 208.1 > 165.1 | 208.1 > 137.1 | −60 | −20 | 0.2 | 446 |
| [D5]-FA 3:0 | 1.61 | 213.1 > 170.1 | 213.1 > 137.1 | −60 | −20 | - | - |
| FA 4:0 | 2.53 | 222.1 > 179.1 | 222.1 > 137.1 | −60 | −20 | 0.06 | 440 |
| FA 3:0 (2Me) | 2.36 | 222.1 > 179.1 | 222.1 > 137.1 | −60 | −20 | 0.03 | 235 |
| [D7]-FA 4:0 | 2.47 | 229.1 > 185.1 | 229.1 > 137.1 | −60 | −20 | - | - |
| FA 2:0 | FA 3:0 | FA 4:0 | FA 3:0(2Me) | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | CV | Mean | CV | Mean | CV | Mean | CV | |
| intra-day (n = 5) | ||||||||
| low | 16 | 5.1% | 4.6 | 6.1% | 4.2 | 1.9% | 1.3 | 5.1% |
| medium | 203 | 3.8% | 55 | 2.8% | 65 | 1.3% | 12.5 | 2.3% |
| high | 628 | 2.6% | 141 | 4.1% | 122 | 2.1% | 11.7 | 1.9% |
| inter-day (n = 6) | ||||||||
| low | 16 | 3.5% | 4.5 | 6.8% | 4.1 | 7.5% | 1.4 | 9.6% |
| medium | 190 | 5.9% | 53 | 6.5% | 62 | 4.7% | 13.8 | 3.5% |
| high | 561 | 3.9% | 139 | 4.8% | 116 | 5.8% | 12.6 | 4.8% |
| SCFA (µmol/g DW) | Mean ± SD | Median | Min | Max |
|---|---|---|---|---|
| FA 2:0 | 399 ± 270 | 341 | 102 | 1210 |
| FA 3:0 | 155 ± 97 | 121 | 19 | 340 |
| FA 4:0 | 151 ± 95 | 137 | 18 | 370 |
| FA 3:0(2Me) | 22 ± 9 | 19 | 10 | 41 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebisch, G.; Ecker, J.; Roth, S.; Schweizer, S.; Öttl, V.; Schött, H.-F.; Yoon, H.; Haller, D.; Holler, E.; Burkhardt, R.; et al. Quantification of Fecal Short Chain Fatty Acids by Liquid Chromatography Tandem Mass Spectrometry—Investigation of Pre-Analytic Stability. Biomolecules 2019, 9, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040121
Liebisch G, Ecker J, Roth S, Schweizer S, Öttl V, Schött H-F, Yoon H, Haller D, Holler E, Burkhardt R, et al. Quantification of Fecal Short Chain Fatty Acids by Liquid Chromatography Tandem Mass Spectrometry—Investigation of Pre-Analytic Stability. Biomolecules. 2019; 9(4):121. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040121
Chicago/Turabian StyleLiebisch, Gerhard, Josef Ecker, Sebastian Roth, Sabine Schweizer, Veronika Öttl, Hans-Frieder Schött, Hongsup Yoon, Dirk Haller, Ernst Holler, Ralph Burkhardt, and et al. 2019. "Quantification of Fecal Short Chain Fatty Acids by Liquid Chromatography Tandem Mass Spectrometry—Investigation of Pre-Analytic Stability" Biomolecules 9, no. 4: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040121