Structural Stabilization of Human Transthyretin by Centella asiatica (L.) Urban Extract: Implications for TTR Amyloidosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Purification of huTTR from Plasma
2.2. Preparation of CAB
2.3. Nitroblue Tetrazolium (NBT) Redox-Cycling Assay
2.4. Determination of the Stability of huTTR in the Presence of CAB
2.4.1. Against Urea-Mediated Denaturation
2.4.2. Against Acid-Mediated Denaturation
2.5. Determination of 8,1-ANS Binding Displacement from huTTR by CAB
2.6. Determination of the Influence of CAB on huTTR Fibrillation by Transmission Electron Microscopy (TEM)
2.7. Determination of CAB Antioxidant Properties
2.7.1. DPPH (1,1-Diphenyl-2-picrylhydrazyl) Radical Scavenging Activity
2.7.2. Ferric Reducing Antioxidant Power (FRAP)
2.8. Determination of the Chemical Composition of CAB
2.8.1. Total Phenolic Content
2.8.2. Total Flavonoid Content
2.8.3. Thin-Layer Chromatography (TLC) Profile of CAB
2.8.4. High-Performance Liquid Chromatography-Mass Spectrometry (HPLC-MS) Fingerprint of CAB
2.9. Statistical Analysis
3. Results
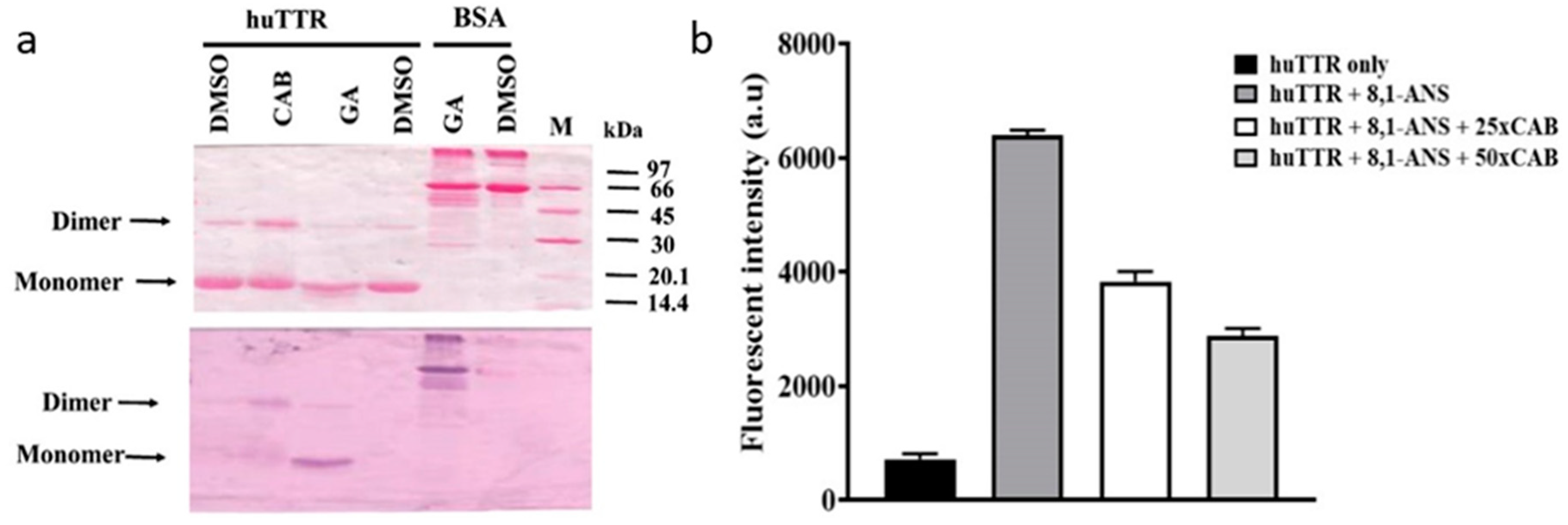
3.1. CAB Directly Binds to huTTR
3.2. CAB Binds to the T4-Binding Sites of huTTR
3.3. CAB Increases huTTR Structural Stability
3.4. CAB Prevents huTTR Fibril Formation
3.5. Antioxidant Activity of CAB
3.6. Chemical Characterization of CAB
3.6.1. Total Phenolic and Flavonoid Contents
3.6.2. TLC Profile of CAB
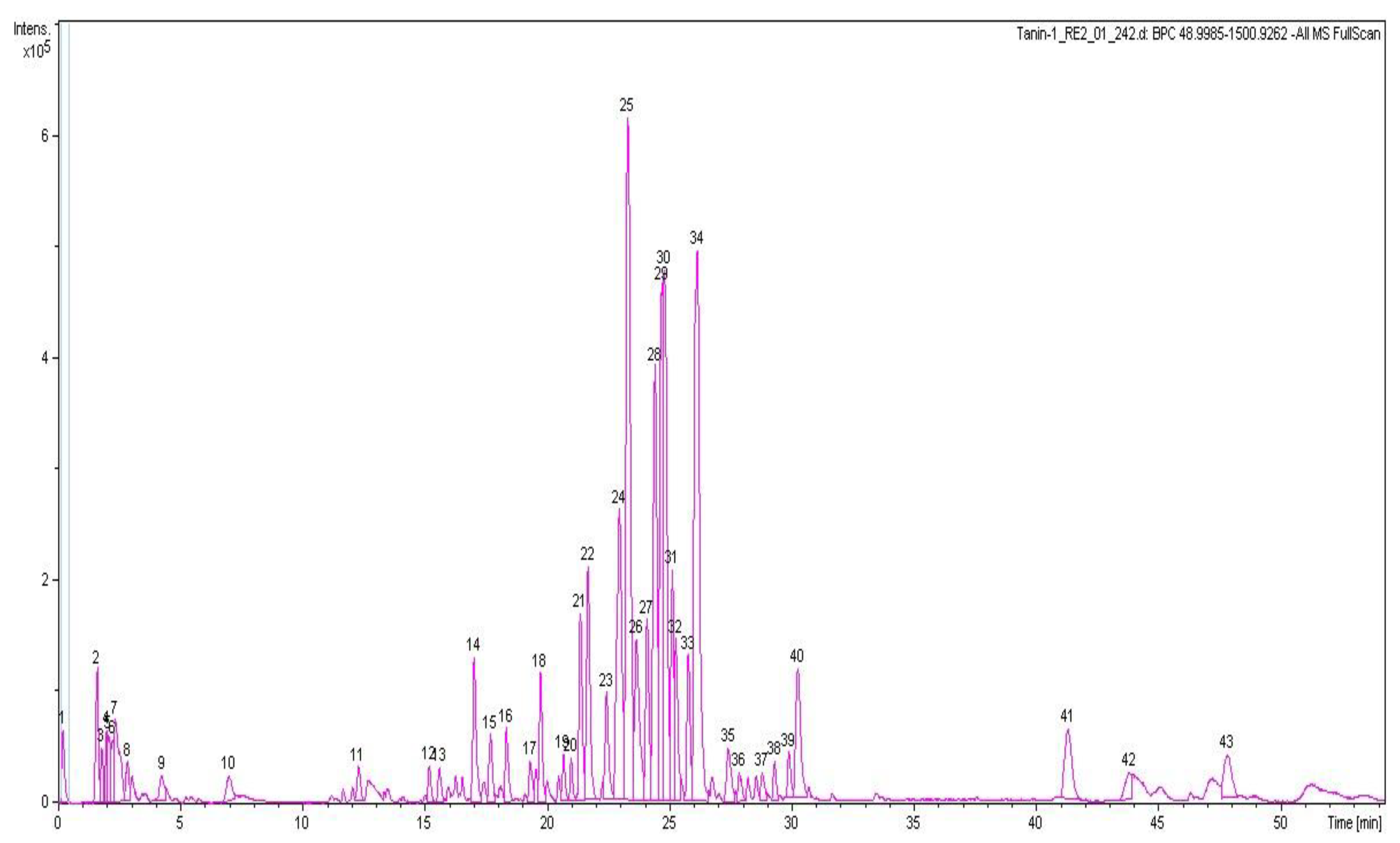
3.6.3. HPLC-QTOF-MS Analysis of CAB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Sipe, J.D.; Benson, M.D.; Buxbaum, J.N.; Ikeda, S.; Merlini, G.; Saraiva, M.J.M.; Westermark, P. Amyloid fibril proteins and amyloidosis: Chemical identification and clinical classification International Society of Amyloidosis 2016 Nomenclature Guidelines. Amyloid 2016, 23, 209–213. [Google Scholar] [CrossRef]
- Reixach, N.; Deechongkit, S.; Jiang, X.; Kelly, J.W.; Buxbaum, J.N. Tissue damage in the amyloidoses: Transthyretin monomers and nonnative oligomers are the major cytotoxic species in tissue culture. Proc. Natl. Acad. Sci. USA 2004, 101, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, M.; Cogan, U.; Mokady, S.; Shinitzky, M. The interaction between retinol-binding proteins and prealbumins studied by fluorescence polarization. Biochim. Biophys. Acta (BBA)-Protein Struct. 1976, 439, 449–460. [Google Scholar] [CrossRef]
- González-López, E.; Gallego-Delgado, M.; Guzzo-Merello, G.; de Haro-Del Moral, F.J.; Cobo-Marcos, M.; Robles, C.; Bornstein, B.; Salas, C.; Lara-Pezzi, E.; Alonso-Pulpon, L.; et al. Wild-type transthyretin amyloidosis as a cause of heart failure with preserved ejection fraction. Eur. Heart J. 2015, 36, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Cornwell, G.G.; Murdoch, W.L.; Kyle, R.A.; Westermark, P.; Pitkänen, P. Frequency and distribution of senile cardiovascular amyloid: A clinicopathologic correlation. Am. J. Med. 1983, 75, 618–623. [Google Scholar] [CrossRef]
- Westermark, P.; Sletten, K.; Johansson, B.; Cornwell, G.G. Fibril in senile systemic amyloidosis is derived from normal transthyretin. Proc. Natl. Acad. Sci. USA 1990, 87, 2843–2845. [Google Scholar] [CrossRef]
- Kelly, J.W. Alternative conformations of amyloidogenic proteins govern their behavior. Curr. Opin. Struct. Biol. 1996, 6, 11–17. [Google Scholar] [CrossRef]
- Johnson, S.M.; Wiseman, R.L.; Sekijima, Y.; Green, N.S.; Adamski-Werner, S.L.; Kelly, J.W. Native state kinetic stabilization as a strategy to ameliorate protein misfolding diseases: A focus on the transthyretin amyloidoses. Acc. Chem. Res. 2005, 38, 911–921. [Google Scholar] [CrossRef]
- Schmidt, H.H.-J. Tafamidis for the treatment of transthyretin-associated familial amyloid polyneuropathy. Expert Opin. Orphan Drugs 2013, 1, 837–845. [Google Scholar] [CrossRef]
- Planté-Bordeneuve, V.; Gorram, F.; Salhi, H.; Nordine, T.; Ayache, S.S.; Le Corvoisier, P.; Azoulay, D.; Feray, C.; Damy, T.; Lefaucheur, J.-P. Long-term treatment of transthyretin familial amyloid polyneuropathy with tafamidis: A clinical and neurophysiological study. J. Neurol. 2017, 264, 268–276. [Google Scholar] [CrossRef]
- Lai, Z.; Colón, W.; Kelly, J.W. The acid-mediated denaturation pathway of transthyretin yields a conformational intermediate that can self-assemble into amyloid. Biochemistry 1996, 35, 6470–6482. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Smith, C.S.; Petrassi, H.M.; Hammarström, P.; White, J.T.; Sacchettini, J.C.; Kelly, J.W. An Engineered Transthyretin Monomer that Is Nonamyloidogenic, Unless It Is Partially Denatured. Biochemistry 2001, 40, 11442–11452. [Google Scholar] [CrossRef]
- Obici, L.; Merlini, G. An overview of drugs currently under investigation for the treatment of transthyretin-related hereditary amyloidosis. Expert Opin. Investig. Drugs 2014, 23, 1239–1251. [Google Scholar] [CrossRef]
- Lashuel, H.A.; Wurth, C.; Woo, L.; Kelly, J.W. The most pathogenic transthyretin variant, L55P, forms amyloid fibrils under acidic conditions and protofilaments under physiological conditions. Biochemistry 1999, 38, 13560–13573. [Google Scholar] [CrossRef]
- Saito, S.; Ando, Y.; Nakamura, M.; Ueda, M.; Kim, J.; Ishima, Y.; Akaike, T.; Otagiri, M. Effect of nitric oxide in amyloid fibril formation on transthyretin-related amyloidosis. Biochemistry 2005, 44, 11122–11129. [Google Scholar] [CrossRef]
- Zhao, L.; Buxbaum, J.N.; Reixach, N. Age-related oxidative modifications of transthyretin modulate its amyloidogenicity. Biochemistry 2013, 52, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Ortore, G.; Orlandini, E.; Braca, A.; Ciccone, L.; Rossello, A.; Martinelli, A.; Nencetti, S. Targeting Different Transthyretin Binding Sites with Unusual Natural Compounds. ChemMedChem 2016, 11, 1865–1874. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Kosaka, Y.; Mizuguchi, M. Inhibitory activities of propolis and its promising component, caffeic acid phenethyl ester, against amyloidogenesis of human transthyretin. J. Med. Chem. 2014, 57, 8928–8935. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Kim, M.-S.; Park, S.-Y. Effect of natural antioxidants on the aggregation and disaggregation of beta-amyloid. Trop. J. Pharm. Res. 2017, 16, 2629–2635. [Google Scholar] [CrossRef]
- Gohil, K.J.; Patel, J.A.; Gajjar, A.K. Pharmacological Review on Centella asiatica: A Potential Herbal Cure-all. Indian J. Pharm. Sci. 2010, 72, 546–556. [Google Scholar] [CrossRef]
- Alqahtani, A.; Tongkao-on, W.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K.; Li, G.Q. Seasonal Variation of Triterpenes and Phenolic Compounds in Australian Centella asiatica (L.) Urb. Phytochem. Anal. 2015, 26, 436–443. [Google Scholar] [CrossRef]
- Berrocal, R.; Vasudevaraju, P.; Indi, S.S.; Sambasiva Rao, K.R.S.; Rao, K.S. In vitro evidence that an aqueous extract of Centella asiatica modulates α-synuclein aggregation dynamics. J. Alzheimers Dis. 2014, 39, 457–465. [Google Scholar] [CrossRef]
- Gray, N.E.; Morré, J.; Kelley, J.; Maier, C.S.; Stevens, J.F.; Quinn, J.F.; Soumyanath, A. Caffeoylquinic acids in Centella asiatica protect against β-amyloid toxicity. J. Alzheimers Dis. 2014, 40, 359–373. [Google Scholar] [CrossRef]
- Prapunpoj, P.; Leelawatwatana, L.; Schreiber, G.; Richardson, S.J. Change in structure of the N-terminal region of transthyretin produces change in affinity of transthyretin to T4 and T3. FEBS J. 2006, 273, 4013–4023. [Google Scholar] [CrossRef]
- Leelawatwattana, L.; Praphanphoj, V.; Prapunpoj, P. Effect of the N-terminal sequence on the binding affinity of transthyretin for human retinol-binding protein. FEBS J. 2011, 278, 3337–3347. [Google Scholar] [CrossRef]
- Ernst, O.; Zor, T. Linearization of the Bradford Protein Assay. J. Vis. Exp. 2010, 38. [Google Scholar] [CrossRef]
- Hellström, J.K.; Mattila, P.H. HPLC determination of extractable and unextractable proanthocyanidins in plant materials. J. Agric. Food Chem. 2008, 56, 7617–7624. [Google Scholar] [CrossRef]
- Paz, M.A.; Flückiger, R.; Boak, A.; Kagan, H.M.; Gallop, P.M. Specific detection of quinoproteins by redox-cycling staining. J. Biol. Chem. 1991, 266, 689–692. [Google Scholar]
- Quintas, A.; Saraiva, M.J.M.; Brito, R.M.M. The Tetrameric Protein Transthyretin Dissociates to a Non-native Monomer in Solution a novel model for amyloidogenesis. J. Biol. Chem. 1999, 274, 32943–32949. [Google Scholar] [CrossRef]
- Mu, Y.; Jin, S.; Shen, J.; Sugano, A.; Takaoka, Y.; Qiang, L.; Imbimbo, B.P.; Yamamura, K.; Li, Z. CHF5074 (CSP-1103) stabilizes human transthyretin in mice humanized at the transthyretin and retinol-binding protein loci. FEBS Lett. 2015, 589, 849–856. [Google Scholar] [CrossRef]
- Nilsson, L.; Larsson, A.; Begum, A.; Iakovleva, I.; Carlsson, M.; Brännström, K.; Sauer-Eriksson, A.E.; Olofsson, A. Modifications of the 7-Hydroxyl Group of the Transthyretin Ligand Luteolin Provide Mechanistic Insights into Its Binding Properties and High Plasma Specificity. PLoS ONE 2016, 11, e0153112. [Google Scholar] [CrossRef]
- Pullakhandam, R.; Srinivas, P.N.B.S.; Nair, M.K.; Reddy, G.B. Binding and stabilization of transthyretin by curcumin. Arch. Biochem. Biophys. 2009, 485, 115–119. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Pages, R.A.; Saroff, H.A.; Edelhoch, H.; Robbins, J. Analysis of thyroid hormone binding to human serum prealbumin by 8-anilinonaphthalene-1-sulfonate fluorescence. Biochemistry 1977, 16, 3707–3713. [Google Scholar] [CrossRef]
- Ferreira, N.; Pereira-Henriques, A.; Almeida, M.R. Transthyretin chemical chaperoning by flavonoids: Structure-activity insights towards the design of potent amyloidosis inhibitors. Biochem. Biophys. Rep. 2015, 3, 123–133. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Pintać, D.; Majkić, T.; Torović, L.; Orčić, D.; Beara, I.; Simin, N.; Mimica–Dukić, N.; Lesjak, M. Solvent selection for efficient extraction of bioactive compounds from grape pomace. Ind. Crops Prod. 2018, 111, 379–390. [Google Scholar] [CrossRef]
- Long, H.S.; Stander, M.A.; Van Wyk, B.-E. Notes on the occurrence and significance of triterpenoids (asiaticoside and related compounds) and caffeoylquinic acids in Centella species. S. Afr. J. Bot. 2012, 82, 53–59. [Google Scholar] [CrossRef]
- Iwamoto, N.; Sumi, D.; Ishii, T.; Uchida, K.; Cho, A.K.; Froines, J.R.; Kumagai, Y. Chemical knockdown of protein-tyrosine phosphatase 1B by 1,2-naphthoquinone through covalent modification causes persistent transactivation of epidermal growth factor receptor. J. Biol. Chem. 2007, 282, 33396–33404. [Google Scholar] [CrossRef]
- McCutchen, S.L.; Kelly, J.W. Intermolecular disulfide linkages are not required for transthyretin amyloid fibril formation in vitro. Biochem. Biophys. Res. Commun. 1993, 197, 415–421. [Google Scholar] [CrossRef]
- Schneider, F.; Hammarström, P.; Kelly, J.W. Transthyretin slowly exchanges subunits under physiological conditions: A convenient chromatographic method to study subunit exchange in oligomeric proteins. Protein Sci. 2001, 10, 1606–1613. [Google Scholar] [CrossRef]
- Miranker, A.D.; Dobson, C.M. Collapse and cooperativity in protein folding. Curr. Opin. Struct. Biol. 1996, 6, 31–42. [Google Scholar] [CrossRef]
- Yang, D.T.; Joshi, G.; Cho, P.Y.; Johnson, J.A.; Murphy, R.M. Transthyretin as both a sensor and a scavenger of β-amyloid oligomers. Biochemistry 2013, 52, 2849–2861. [Google Scholar] [CrossRef]
- Lashuel, H.A.; Lai, Z.; Kelly, J.W. Characterization of the transthyretin acid denaturation pathways by analytical ultracentrifugation: Implications for wild-type, V30M, and L55P amyloid fibril formation. Biochemistry 1998, 37, 17851–17864. [Google Scholar] [CrossRef]
- Fernandes, R.P.P.; Trindade, M.A.; Tonin, F.G.; Lima, C.G.; Pugine, S.M.P.; Munekata, P.E.S.; Lorenzo, J.M.; de Melo, M.P. Evaluation of antioxidant capacity of 13 plant extracts by three different methods: Cluster analyses applied for selection of the natural extracts with higher antioxidant capacity to replace synthetic antioxidant in lamb burgers. J. Food Sci. Technol. 2016, 53, 451–460. [Google Scholar] [CrossRef]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef]
- Wagner, H.; Bladt, S. Saponin Drugs. In Plant Drug Analysis: A Thin Layer Chromatography Atlas, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Maulidiani; Abas, F.; Khatib, A.; Shaari, K.; Lajis, N.H. Chemical characterization and antioxidant activity of three medicinal Apiaceae species. Ind. Crops Prod. 2014, 55, 238–247. [Google Scholar] [CrossRef]
- Colon, W.; Kelly, J.W. Partial denaturation of transthyretin is sufficient for amyloid fibril formation in vitro. Biochemistry 1992, 31, 8654–8660. [Google Scholar] [CrossRef]
- Kelly, J.W.; Colon, W.; Lai, Z.; Lashuel, H.A.; McCulloch, J.; McCutchen, S.L.; Miroy, G.J.; Peterson, S.A. Transthyretin quaternary and tertiary structural changes facilitate misassembly into amyloid. Adv. Protein Chem. 1997, 50, 161–181. [Google Scholar]
- Hammarström, P.; Jiang, X.; Deechongkit, S.; Kelly, J.W. Anion shielding of electrostatic repulsions in transthyretin modulates stability and amyloidosis: Insight into the chaotrope unfolding dichotomy. Biochemistry 2001, 40, 11453–11459. [Google Scholar] [CrossRef]
- Sekijima, Y.; Kelly, J.W.; Ikeda, S. Pathogenesis of and therapeutic strategies to ameliorate the transthyretin amyloidoses. Curr. Pharm. Des. 2008, 14, 3219–3230. [Google Scholar] [CrossRef]
- Ferreira, N.; Gonçalves, N.P.; Saraiva, M.J.; Almeida, M.R. Curcumin: A multi-target disease-modifying agent for late-stage transthyretin amyloidosis. Sci. Rep. 2016, 6, 26623. [Google Scholar] [CrossRef]
- Florio, P.; Folli, C.; Cianci, M.; Del Rio, D.; Zanotti, G.; Berni, R. Transthyretin Binding Heterogeneity and Anti-amyloidogenic Activity of Natural Polyphenols and Their Metabolites. J. Biol. Chem. 2015, 290, 29769–29780. [Google Scholar] [CrossRef]
- Hamilton, J.A.; Benson, M.D. Transthyretin: A review from a structural perspective. Cell. Mol. Life Sci. 2001, 58, 1491–1521. [Google Scholar] [CrossRef]
- Ishii, T.; Mori, T.; Tanaka, T.; Mizuno, D.; Yamaji, R.; Kumazawa, S.; Nakayama, T.; Akagawa, M. Covalent modification of proteins by green tea polyphenol (-)-epigallocatechin-3-gallate through autoxidation. Free Radic. Biol. Med. 2008, 45, 1384–1394. [Google Scholar] [CrossRef]
- Tomar, D.; Khan, T.; Singh, R.R.; Mishra, S.; Gupta, S.; Surolia, A.; Salunke, D.M. Crystallographic Study of Novel Transthyretin Ligands Exhibiting Negative-Cooperativity between Two Thyroxine Binding Sites. PLoS ONE 2012, 7, e43522. [Google Scholar] [CrossRef]
- Ferreira, N.; Saraiva, M.J.; Almeida, M.R. Natural polyphenols inhibit different steps of the process of transthyretin (TTR) amyloid fibril formation. FEBS Lett. 2011, 585, 2424–2430. [Google Scholar] [CrossRef]
- Lakey-Beitia, J.; Berrocal, R.; Rao, K.S.; Durant, A.A. Polyphenols as therapeutic molecules in Alzheimer’s disease through modulating amyloid pathways. Mol. Neurobiol. 2015, 51, 466–479. [Google Scholar] [CrossRef]
- Manral, P.; Reixach, N. Amyloidogenic and non-amyloidogenic transthyretin variants interact differently with human cardiomyocytes: Insights into early events of non-fibrillar tissue damage. Biosci. Rep. 2015, 35, e00172. [Google Scholar] [CrossRef]
- Kugimiya, T.; Jono, H.; Saito, S.; Maruyama, T.; Kadowaki, D.; Misumi, Y.; Hoshii, Y.; Tasaki, M.; Su, Y.; Ueda, M.; et al. Loss of functional albumin triggers acceleration of transthyretin amyloid fibril formation in familial amyloidotic polyneuropathy. Lab. Investig. 2011, 91, 1219–1228. [Google Scholar] [CrossRef]
- Macedo, B.; Magalhães, J.; Batista, A.R.; Saraiva, M.J. Carvedilol treatment reduces transthyretin deposition in a familial amyloidotic polyneuropathy mouse model. Pharmacol. Res. 2010, 62, 514–522. [Google Scholar] [CrossRef]
- Mustafa, R.A.; Abdul Hamid, A.; Mohamed, S.; Bakar, F.A. Total phenolic compounds, flavonoids, and radical scavenging activity of 21 selected tropical plants. J. Food Sci. 2010, 75, C28–C35. [Google Scholar] [CrossRef]
- Pittella, F.; Dutra, R.C.; Junior, D.D.; Lopes, M.T.P.; Barbosa, N.R. Antioxidant and Cytotoxic Activities of Centella asiatica (L) Urb. Int. J. Mol. Sci. 2009, 10, 3713–3721. [Google Scholar] [CrossRef]
- Guo, J.; Jono, H.; Kugimiya, T.; Saito, S.; Maruyama, T.; Misumi, Y.; Hoshii, Y.; Su, Y.; Shono, M.; Ueda, M.; et al. Antioxidative effect of albumin on amyloid fibril formation in transthyretin-related amyloidosis. Amyloid 2011, 18 (Suppl. 1), 17–18. [Google Scholar] [CrossRef]
- Alemi, M.; Silva, S.C.; Santana, I.; Cardoso, I. Transthyretin stability is critical in assisting beta amyloid clearance- Relevance of transthyretin stabilization in Alzheimer’s disease. CNS Neurosci. Ther. 2017, 23, 605–619. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, Z.; Khan, I.; Luo, Y. Gotu Kola (Centella Asiatica) extract enhances phosphorylation of cyclic AMP response element binding protein in neuroblastoma cells expressing amyloid beta peptide. J. Alzheimers Dis. 2008, 13, 341–349. [Google Scholar] [CrossRef]
- Gray, N.E.; Harris, C.J.; Quinn, J.F.; Soumyanath, A. Centella asiatica modulates antioxidant and mitochondrial pathways and improves cognitive function in mice. J. Ethnopharmacol. 2016, 180, 78–86. [Google Scholar] [CrossRef]
- Wanakhachornkrai, O.; Pongrakhananon, V.; Chunhacha, P.; Wanasuntronwong, A.; Vattanajun, A.; Tantisira, B.; Chanvorachote, P.; Tantisira, M.H. Neuritogenic effect of standardized extract of Centella asiatica ECa233 on human neuroblastoma cells. BMC Complement. Altern. Med. 2013, 13, 204. [Google Scholar] [CrossRef]
- Chanana, P.; Kumar, A. Possible Involvement of Nitric Oxide Modulatory Mechanisms in the Neuroprotective Effect of Centella asiatica Against Sleep Deprivation Induced Anxiety Like Behaviour, Oxidative Damage and Neuroinflammation. Phytother. Res. 2016, 30, 671–680. [Google Scholar] [CrossRef]
- Kumari, S.; Deori, M.; Elancheran, R.; Kotoky, J.; Devi, R. In vitro and In vivo Antioxidant, Anti-hyperlipidemic Properties and Chemical Characterization of Centella asiatica (L.) Extract. Front. Pharmacol. 2016, 7, 400. [Google Scholar] [CrossRef]
- Ramesh, B.N.; Girish, T.K.; Raghavendra, R.H.; Naidu, K.A.; Rao, U.J.S.P.; Rao, K.S. Comparative study on anti-oxidant and anti-inflammatory activities of Caesalpinia crista and Centella asiatica leaf extracts. J. Pharm. Bioallied Sci. 2014, 6, 86–91. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time | Observed Accurate Mass (m/z) | Predicted Formula | Calculated Mass (Da) | Tentative Identity of Compound |
|---|---|---|---|---|---|
| 4 | 2.0 | 503.1527 [M − H]− | C21H28O14 | 504.1479 | Caffeic acid dihexoside |
| 12 | 15.1 | 353.0877 [M − H]− | C16H18O9 | 354.0951 | 3-O-Caffeoylquinic acid |
| 14 | 17.0 | 353.088 [M − H]− | C16H18O9 | 354.0951 | 5-O-Caffeoylquinic acid |
| 21 | 21.3 | 693.2790 [M − H2O − H]− | C34H48O16 | 712.2942 | Nominilic acid 17-glucoside |
| 22 | 21.6 | 477.0685 [M − H]− | C21H18O13 | 478.0747 | Quercetin 3-O-glucoronide |
| 23 | 22.4 | 5151195 [M − H]− | C25H24O12 | 516.1268 | 3,4-O-Dicaffeoylquinic acid |
| 24 | 22.9 | 515.1209 [M − H]− | C25H24O12 | 516.1268 | 3,5-O-Dicaffeoylquinic acid |
| 25 | 23.3 | 601.1226 [M − H]− | C28H26O15 | 602.1272 | 3,5-O-Dicaffeoyl-4-malonylquinic acid |
| 26 | 23.6 | 515.1203 [M − H]− | C25H24O12 | 516.1268 | 4,5-O-Dicaffeoylquinic acid |
| 27 | 24.0 | 601.1209 [M − H]− | C28H26O15 | 602.1272 | 3,5-O-Dicaffeoyl-4-malonylquinic acid isomer |
| 28 | 24.4 | 601.1228 [M − H]− | C28H26O15 | 602.1272 | Eriodictyol 7-(6-galloylglucoside) |
| 29 | 24.7 | 1019.5149 [M + Formate]− | C49H79O22 | 1019.5063 | Asiaticoside B |
| 30 | 24.8 | 1019.5149 [M + Formate]− | C49H79O22 | 1019.5063 | Madecassoside |
| 31 | 25.1 | 529.1351 [M − H]− | C26H26O12 | 530.1424 | 3-Caffeoyl-4-feruloylquinic acid |
| 32 | 25.2 | 873.4522 [M + Formate − H]− | C43H69O18 | 874.4562 | Centellasaponin B |
| 33 | 25.7 | 1003.5166 [M + Formate − H]− | C49H79O21 | 1004.5192 | Centellasaponin A |
| 34 | 26.1 | 957.5088 [M − H]− | C48H78O19 | 958.5137 | Asiaticoside |
| 36 | 27.8 | 1061.5180 [M − H]− | C51H82O23 | 1062.5247 | Avenacoside A |
| 39 | 29.8 | 987.5213 [M − H]− | C49H79O20 | 988.5243 | Soyasaponin I |
| 40 | 30.2 | 571.0881 [M − H2O − H]− | C30H20O12 | 572.0955 | Manniflavanone |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eze, F.N.; Leelawatwattana, L.; Prapunpoj, P. Structural Stabilization of Human Transthyretin by Centella asiatica (L.) Urban Extract: Implications for TTR Amyloidosis. Biomolecules 2019, 9, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040128
Eze FN, Leelawatwattana L, Prapunpoj P. Structural Stabilization of Human Transthyretin by Centella asiatica (L.) Urban Extract: Implications for TTR Amyloidosis. Biomolecules. 2019; 9(4):128. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040128
Chicago/Turabian StyleEze, Fredrick Nwude, Ladda Leelawatwattana, and Porntip Prapunpoj. 2019. "Structural Stabilization of Human Transthyretin by Centella asiatica (L.) Urban Extract: Implications for TTR Amyloidosis" Biomolecules 9, no. 4: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040128