Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species

Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Extract
2.2. Reagents
2.3. Cell Lines
2.4. Assay for Cell Viability
2.5. Assay for Colony Formation
2.6. Assay for DNA Laddering
2.7. Assay for Nuclear Morphology
2.8. Assay for Cell Viability by Staining with Acridine Orange/Propidium Iodide
2.9. Assay for Sub-G1 Population
2.10. Assay for Mitochondrial Membrane Potential (ΔΨ)
2.11. Assay for Protein Expression
2.12. Assay for Cell Migration
2.13. Assay for Reactive Oxygen Species Generation
2.14. Gas Chromatography-Mass Spectrometry
3. Results
3.1. Curcuma raktakanda Suppresses Viability and Reduces Long-Term Colony Formation of Glioma Cells
3.2. Acetone Extract induces Apoptosis in Glioma Cells
3.3. Acetone Extract Induces Cell Cycle Arrest and Lowers Mitochondrial Membrane Potential in Glioma Cells
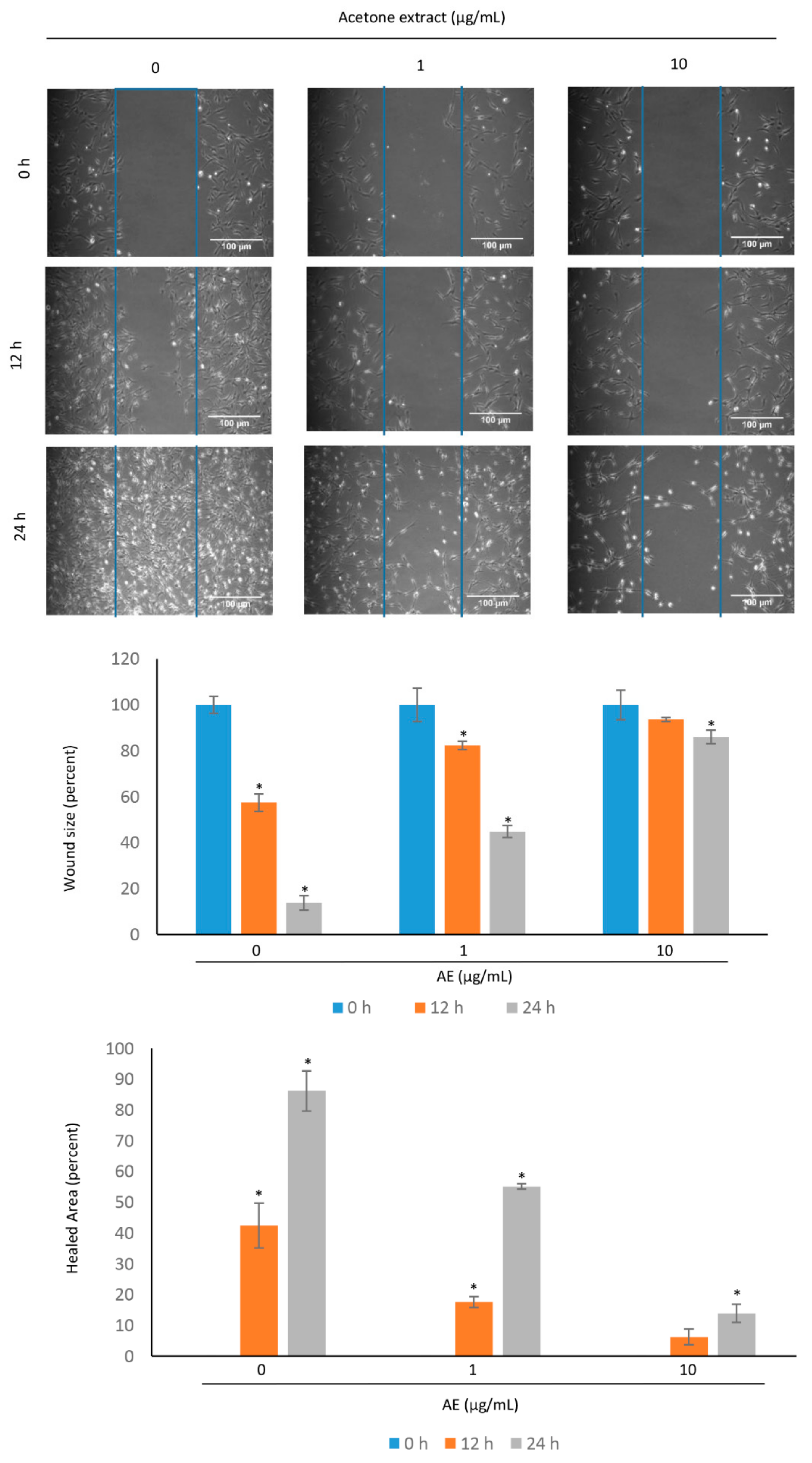
3.4. Acetone Extract Suppresses the Migration of Glioma Cells
3.5. Acetone Extract Induces Reactive Oxygen Species Generation in Glioma Cells
3.6. Curcuma raktakanda Reduces the Viability of Breast and Cervical Cancer Cells
3.7. Gas Chromatography-Mass Spectrometry Analysis of Curcuma raktakanda Rhizome Fractions
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Shirai, K.; Chakravarti, A. Towards personalized therapy for patients with glioblastoma. Expert Rev. Anticancer. Ther. 2011, 11, 1935–1944. [Google Scholar] [CrossRef]
- Greenlee, R.T.; Murray, T.; Bolden, S.; Wingo, P.A. Cancer statistics, 2000. CA Cancer J. Clin. 2000, 50, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Régina, A.; Demeule, M.; Laplante, A.; Jodoin, J.; Dagenais, C.; Berthelet, F.; Moghrabi, A.; Béliveau, R. Multidrug resistance in brain tumors: Roles of the blood–brain barrier. Cancer Metastasis Rev. 2001, 20, 13–25. [Google Scholar] [CrossRef]
- Mirimanoff, R.-O.; Gorlia, T.; Mason, W.; Van den Bent, M.J.; Kortmann, R.-D.; Fisher, B.; Reni, M.; Brandes, A.A.; Curschmann, J.; Villa, S. Radiotherapy and temozolomide for newly diagnosed glioblastoma: Recursive partitioning analysis of the EORTC 26981/22981-NCIC CE3 phase III randomized trial. J. Clin. Oncol. 2006, 24, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Evasion of apoptosis as a cellular stress response in cancer. Int. J. Cell Biol. 2010, 2010. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S. The role of natural product chemistry in drug discovery. J. Nat. Prod. 2004, 67, 2141–2153. [Google Scholar] [CrossRef] [PubMed]
- Balunas, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef]
- Wani, M.C.; Horwitz, S.B. Nature as a remarkable chemist: A personal story of the discovery and development of Taxol. Anti-Cancer Drugs 2014, 25, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Holmes, F.A.; Walters, R.S.; Theriault, R.L.; Buzdar, A.U.; Frye, D.K.; Hortobagyi, G.N.; Forman, A.D.; Newton, L.K.; Raber, M.N. Phase II trial of taxol, an active drug in the treatment of metastatic breast cancer. J. Natl. Cancer Inst. 1991, 83, 1797–1805. [Google Scholar] [CrossRef]
- McGuire, W.P.; Rowinsky, E.K.; Rosenshein, N.B.; Grumbine, F.C.; Ettinger, D.S.; Armstrong, D.K.; Donehower, R.C. Taxol: A unique antineoplastic agent with significant activity in advanced ovarian epithelial neoplasms. Ann. Intern. Med. 1989, 111, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Misra, S.K.; Saha, M.L.; Lahiri, N.; Louie, J.; Pan, D.; Stang, P.J. Orthogonal self-assembly of an organoplatinum (II) metallacycle and cucurbit [8] uril that delivers curcumin to cancer cells. Proc. Natl. Acad. Sci. USA 2018, 115, 8087–8092. [Google Scholar] [CrossRef]
- Banerjee, S.; Ji, C.; Mayfield, J.E.; Goel, A.; Xiao, J.; Dixon, J.E.; Guo, X. Ancient drug curcumin impedes 26S proteasome activity by direct inhibition of dual-specificity tyrosine-regulated kinase 2. Proc. Natl. Acad. Sci. USA 2018, 115, 8155–8160. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Sung, B.; Kim, J.H.; Prasad, S.; Li, S.; Aggarwal, B.B. Multitargeting by turmeric, the golden spice: From kitchen to clinic. Mol. Nutr. Food Res. 2013, 57, 1510–1528. [Google Scholar] [CrossRef] [PubMed]
- Tohda, C.; Nakayama, N.; Hatanaka, F.; Komatsu, K. Comparison of anti-inflammatory activities of six Curcuma rhizomes: A possible curcuminoid-independent pathway mediated by Curcuma phaeocaulis extract. Evid.-Based Complementary Altern. Med. 2006, 3, 255–260. [Google Scholar] [CrossRef]
- Latha, C.; Ammini, J. Curcuma raktakanda is a potential larvicide for mosquito control. Pharm. Biol. 2000, 38, 167–170. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sung, B.; Prasad, S.; Webb, L.J.; Aggarwal, B.B. Cancer drug discovery by repurposing: Teaching new tricks to old dogs. Trends Pharmacol. Sci. 2013, 34, 508–517. [Google Scholar] [CrossRef]
- Awasthee, N.; Rai, V.; Verma, S.S.; Francis, K.S.; Nair, M.S.; Gupta, S.C. Anti-cancer activities of Bharangin against breast cancer: Evidence for the role of NF-κB and lncRNAs. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2738–2749. [Google Scholar] [CrossRef]
- Gupta, S.C.; Prasad, S.; Sethumadhavan, D.R.; Nair, M.S.; Mo, Y.-Y.; Aggarwal, B.B. Nimbolide, a limonoid triterpene, inhibits growth of human colorectal cancer xenografts by suppressing the proinflammatory microenvironment. Clin. Cancer Res. 2013, 19, 4465–4476. [Google Scholar] [CrossRef]
- Herrmann, M.; Lorenz, H.; Voll, R.; Grünke, M.; Woith, W.; Kalden, J. A rapid and simple method for the isolation of apoptotic DNA fragments. Nucleic Acids Res. 1994, 22, 5506. [Google Scholar] [CrossRef]
- Gupta, S.C.; Nair, M.S.; Mo, Y.-Y.; Aggarwal, B.B. Azadirone, a Limonoid Tetranortriterpene, Induces Death Receptors and Sensitizes Human Cancer Cells to TRAIL through a p53-Independent Mechanism: Evidence for the Role of the ROS-ERK-CHOP-Death Receptor Pathway. J. Biol. Chem. 2013, 288, 32343–32356. [Google Scholar] [CrossRef]
- Bognar, Z.; Fekete, K.; Antus, C.; Hocsak, E.; Bognar, R.; Tapodi, A.; Boronkai, A.; Farkas, N.; Gallyas Jr, F.; Sumegi, B. Desethylamiodarone—A metabolite of amiodarone—Induces apoptosis on T24 human bladder cancer cells via multiple pathways. PLoS ONE 2017, 12, e0189470. [Google Scholar] [CrossRef]
- Gupta, S.; Singh, R.; Asters, M.; Liu, J.; Zhang, X.; Pabbidi, M.; Watabe, K.; Mo, Y. Regulation of breast tumorigenesis through acid sensors. Oncogene 2016, 35, 4102. [Google Scholar] [CrossRef]
- Gupta, S.C.; Singh, R.; Pochampally, R.; Watabe, K.; Mo, Y.-Y. Acidosis promotes invasiveness of breast cancer cells through ROS-AKT-NF-κB pathway. Oncotarget 2014, 5, 12070. [Google Scholar] [CrossRef]
- Omoregie, S.N.; Omoruyi, F.O.; Wright, V.F.; Jones, L.; Zimba, P.V. Antiproliferative activities of lesser galangal (Alpinia officinarum Hance Jam1), turmeric (Curcuma longa L.), and ginger (Zingiber officinale Rosc.) against acute monocytic leukemia. J. Med. Food 2013, 16, 647–655. [Google Scholar] [CrossRef]
- Trisciuoglio, D.; Tupone, M.G.; Desideri, M.; Martile, M.; Gabellini, C.; Buglioni, S.; Pallocca, M.; Alessandrini, G.; D’Aguanno, S.; Bufalo, D. BCL-X L overexpression promotes tumor progression-associated properties. Cell Death Dis. 2017, 8, 3216. [Google Scholar] [CrossRef]
- Gabellini, C.; Gómez-Abenza, E.; Ibáñez-Molero, S.; Tupone, M.G.; Pérez-Oliva, A.B.; de Oliveira, S.; Del Bufalo, D.; Mulero, V. Interleukin 8 mediates bcl-xL-induced enhancement of human melanoma cell dissemination and angiogenesis in a zebrafish xenograft model. Int. J. Cancer 2018, 142, 584–596. [Google Scholar] [CrossRef]
- Khoshnan, A.; Tindell, C.; Laux, I.; Bae, D.; Bennett, B.; Nel, A.E. The NF-κB cascade is important in Bcl-xL expression and for the anti-apoptotic effects of the CD28 receptor in primary human CD4+ lymphocytes. J. Immunol. 2000, 165, 1743–1754. [Google Scholar] [CrossRef]
- Choi, S.; Chen, Z.; Tang, L.H.; Fang, Y.; Shin, S.J.; Panarelli, N.C.; Chen, Y.-T.; Li, Y.; Jiang, X.; Du, Y.-C.N. Bcl-xL promotes metastasis independent of its anti-apoptotic activity. Nat. Commun. 2016, 7, 10384. [Google Scholar] [CrossRef] [PubMed]
- Zeuner, A.; Francescangeli, F.; Contavalli, P.; Zapparelli, G.; Apuzzo, T.; Eramo, A.; Baiocchi, M.; De Angelis, M.; Biffoni, M.; Sette, G. Elimination of quiescent/slow-proliferating cancer stem cells by Bcl-X L inhibition in non-small cell lung cancer. Cell Death Differ. 2014, 21, 1877. [Google Scholar] [CrossRef]
- Rao, J.S. Molecular mechanisms of glioma invasiveness: The role of proteases. Nat. Rev. Cancer 2003, 3, 489. [Google Scholar] [CrossRef] [PubMed]
- Walia, M.; Mann, T.S.; Kumar, D.; Agnihotri, V.K.; Singh, B. Chemical composition and in vitro cytotoxic activity of essential oil of leaves of Malus domestica growing in Western Himalaya (India). Evid.-Based Complementary Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-Y.; Zhu, H.; Lin, L.-P.; Miao, Z.-H.; Geng, M.-Y.; Cai, Y.-J.; Chen, Y.; Zhao, H.-J.; Luo, H.-B.; Zhang, X.-W. MFTZ-1, an actinomycetes subspecies–derived antitumor macrolide, functions as a novel topoisomerase II poison. Mol. Cancer Ther. 2007, 6, 3059–3070. [Google Scholar] [CrossRef] [PubMed]
- Leighton, X.; Bera, A.; Eidelman, O.; Eklund, M.; Puthillathu, N.; Pollard, H.B.; Srivastava, M. High ANXA7 Potentiates Eucalyptol Toxicity in Hormone-refractory Prostate Cancer. Anticancer Res. 2018, 38, 3831–3842. [Google Scholar] [CrossRef]
- Taha, A.M.; Eldahshan, O.A. Chemical characteristics, antimicrobial, and cytotoxic activities of the essential oil of Egyptian Cinnamomum glanduliferum bark. Chem. Biodivers. 2017, 14, e1600443. [Google Scholar] [CrossRef]
- Zheng, F.; Tang, Q.; Zheng, X.-h.; Wu, J.; Huang, H.; Zhang, H.; Hann, S.S. Inactivation of Stat3 and crosstalk of miRNA155-5p and FOXO3a contribute to the induction of IGFBP1 expression by beta-elemene in human lung cancer. Exp. Mol. Med. 2018, 50, 121. [Google Scholar] [CrossRef]
- Chang, Z.; Gao, M.; Zhang, W.; Song, L.; Jia, Y.; Qin, Y. Beta-elemene treatment is associated with improved outcomes of patients with esophageal squamous cell carcinoma. Surg. Oncol. 2017, 26, 333–337. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H.; Schilder, Y.D.; Dirsch, V.M. Chronic treatment with resveratrol induces redox stress-and ataxia telangiectasia-mutated (ATM)-dependent senescence in p53-positive cancer cells. J. Biol. Chem. 2007, 282, 26759–26766. [Google Scholar] [CrossRef] [PubMed]
- Hang, W.; Yin, Z.-X.; Liu, G.; Zeng, Q.; Shen, X.-F.; Sun, Q.-H.; Li, D.-D.; Jian, Y.-P.; Zhang, Y.-H.; Wang, Y.-S. Piperlongumine and p53-reactivator APR-246 selectively induce cell death in HNSCC by targeting GSTP1. Oncogene 2018, 37, 3384. [Google Scholar] [CrossRef]
- Gupta, S.C.; Hevia, D.; Patchva, S.; Park, B.; Koh, W.; Aggarwal, B.B. Upsides and downsides of reactive oxygen species for cancer: The roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid. Redox Signal. 2012, 16, 1295–1322. [Google Scholar] [CrossRef]
- Boguski, M.S.; Mandl, K.D.; Sukhatme, V.P. Repurposing with a difference. Science 2009, 324, 1394–1395. [Google Scholar] [CrossRef] [PubMed]
- Naguib, A.; Mathew, G.; Reczek, C.R.; Watrud, K.; Ambrico, A.; Herzka, T.; Salas, I.C.; Lee, M.F.; El-Amine, N.; Zheng, W. Mitochondrial complex I inhibitors expose a vulnerability for selective killing of Pten-Null cells. Cell Rep. 2018, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Récher, C.; Beyne-Rauzy, O.; Demur, C.; Chicanne, G.; Dos Santos, C.; Mansat-De Mas, V.; Benzaquen, D.; Laurent, G.; Huguet, F.; Payrastre, B. Antileukemic activity of rapamycin in acute myeloid leukemia. Blood 2005, 105, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, B.L. Chemically modified non-antimicrobial tetracyclines are multifunctional drugs against advanced cancers. Pharmacol. Res. 2011, 63, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Shabani, S.; Awad, A.; Kaushal, M.; Doan, N. Molecular Markers of Therapy-Resistant Glioblastoma and Potential Strategy to Combat Resistance. Int. J. Mol. Sci. 2018, 19, 1765. [Google Scholar] [CrossRef]
- Yang, S.; Qiang, L.; Sample, A.; Shah, P.; He, Y.-Y. NF-κB signaling activation induced by chloroquine requires autophagosome, p62 protein, and c-Jun N-terminal Kinase (JNK) signaling and promotes tumor cell resistance. J. Biol. Chem. 2017, 292, 3379–3388. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Peak No. | Retention Time (min) | Area % | Compound Name | Molecular Weight | Molecular Formula |
|---|---|---|---|---|---|---|
| 1 | 1 | 18.985 | 0.84 | 13,17-Dimethylhentriacontane | 464.907 | C33H68 |
| 2 | 2 | 19.331 | 0.71 | Hexacosane | 366.718 | C26H54 |
| 3 | 3 | 20.129 | 0.75 | Eicosane | 282.5475 | C20H42 |
| 4 | 4 | 20.541 | 2.41 | Tetracontane | 563.096 | C40H82 |
| 5 | 5 | 21.219 | 1.17 | Tetracontane | 563.096 | C40H82 |
| 6 | 6 | 21.646 | 0.80 | Pentacosane | 352.691 | C25H52 |
| 7 | 7 | 22.129 | 0.12 | Pentacosane | 352.691 | C25H52 |
| 8 | 8 | 22.494 | 0.26 | Dotriacontane | 450.88 | C32H66 |
| 9 | 9 | 22.860 | 2.15 | Tetracontane | 563.079 | C40H82 |
| 10 | 10 | 27.890 | 0.08 | Acetic acid, 1-[2-(2,2,6-trimethyl-bicyclo[4.1.0]hept-1-yl)-ethyl]-vinyl ester | 250.376 | C16H26 |
| 11 | 11 | 39.280 | 1.02 | Celidoniol, deoxy- | 424.798 | C29H60 |
| 12 | 12 | 40.494 | 0.20 | Nonacosane | 408.799 | C29H60 |
| 13 | 13 | 40.727 | 0.20 | Pentatriacontane | 492.961 | C35H72 |
| 14 | 14 | 41.535 | 0.54 | Heneicosyl pentafluoropropionate | 458.5890 | C24H43F5O2 |
| 15 | 15 | 42.352 | 0.40 | 1,30-Triacontanediol | 454.812 | C30H62 |
| 16 | 16 | 43.000 | 4.14 | (2e)-1-methyl-3-(2,6,6-trimethyl-2-cyclohexen-1-yl)-2-propenyl acetate | 236.355 | C15H24O2 |
| 17 | 17 | 43.100 | 6.16 | Dotriacontane | 450.88 | C32H66 |
| 18 | 18 | 43.286 | 38.53 | Tetracontane | 563.096 | C40H82 |
| 19 | 19 | 44.396 | 0.33 | Tetratriacontyl heptafluorobutyrate | 690.9421 | C38H69F7O2 |
| 20 | 20 | 44.620 | 0.21 | Pentatriacontane | 492.961 | C35H72 |
| 21 | 21 | 44.890 | 0.19 | 1-Hentetracontanol | 593.122 | C41H84O |
| 22 | 22 | 45.290 | 0.28 | Tetrapentacontane, 1,54-dibromo- | 917.266 | C54H108Br2 |
| 23 | 23 | 46.220 | 1.67 | Hexatriacontane | 506.988 | C36H74 |
| 24 | 24 | 46.746 | 0.52 | Hexatriacontane | 506.988 | C36H74 |
| 25 | 25 | 48.345 | 9.10 | (-)-(1R,2R,4aS,8aS)-1-(2-Hydroxyethyl)-2,5,5,8a-tetramethyldecahydro-2-naphthalenol | 308.499 | C20H36 |
| 26 | 26 | 48.501 | 27.22 | Tetracontane | 563.096 | C40H82 |
| S. No. | Peak No. | Retention Time (min) | Area % | Compound Name | Molecular Weight | Molecular Formula |
|---|---|---|---|---|---|---|
| 1 | 1 | 8.277 | 4.4 | Eucalyptol | 154.249 | C10H18O |
| 2 | 2 | 11.516 | 5.17 | (+)-2-Bornanone | 152.2334 | C10H16O |
| 3 | 3 | 16.746 | 0.88 | .delta.-Elemene | 204.357 | C15H24 |
| 4 | 4 | 18.206 | 7.44 | 2,4-diisopropenyl-1-methyl-1-vinylcyclohexane | 204.351 | C15H24 |
| 5 | 5 | 19.222 | 5.14 | .gamma.-Elemene | 204.3511 | C15H24 |
| 6 | 6 | 20.739 | 2.45 | beta.-Selinene | 204.357 | C15H24 |
| 7 | 7 | 20.858 | 8.2 | Benzofuran, 6-ethenyl-4,5,6,7-tetrahydro-3,6-dimethyl-5-isopropenyl-, trans- | 216.3187 | C15H20O |
| 8 | 8 | 23.398 | 16.84 | beta.-Elemenone | 218.34 | C15H22O |
| 9 | 9 | 24.044 | 2.18 | 1H-Cycloprop[e]azulen-7-ol, decahydro-1,1,7-trimethyl-4-methylene-, [1ar-(1a.alpha.,4a.alpha.,7.beta.,7a.beta.,7b.alpha.)]- | 220.3505 | C15H24O |
| 10 | 10 | 25.539 | 3.91 | 3,7-Dimethyl-10-(1-methylethylidene)-3,7-cyclodecadien-1-one | 218.3346 | C15H22O |
| 11 | 11 | 26.899 | 8.33 | 5,8-Dihydroxy-4a-methyl-4,4a,4b,5,6,7,8,8a,9,10-decahydro-2(3H)-phenanthrenone | 250.333 | C15H22 |
| 12 | 12 | 27.652 | 10.49 | Cycloprop[e]indene-1a,2(1H)-dicarboxaldehyde, 3a,4,5,6,6a,6b-hexahydro-5,5,6b-trimethyl-, (1a.alpha.,3a.beta.,6a.beta.,6b.alpha | 232.323 | C15H20O2 |
| 13 | 13 | 28.643 | 2.17 | Elemene | 204.3511 | C15H24 |
| 14 | 14 | 28.958 | 6.54 | Acetic acid, 6-(1-hydroxymethyl-vinyl)-4,8a-dimethyl-3-oxo-1,2,3,5,6,7,8,8a-octahydronaphthalen-2-yl ester | 292.375 | C17H24O4 |
| 15 | 15 | 29.872 | 0.72 | Bufa-20,22-dienolide, 14,15-epoxy-3,5,16-trihydroxy-, (3.beta.,5.beta.,15.beta.,16.beta.)- | 500.588 | C28H36O8 |
| 16 | 16 | 30.033 | 1.07 | Cyclohexane, 1-ethenyl-1-methyl-2-(1-methylethenyl)-4-(1-methylethylidene)- | 204.3511 | C15H24 |
| 17 | 17 | 31.13 | 1.41 | n-Hexadecanoic acid | 256.4241 | C16H32O2 |
| 18 | 18 | 31.617 | 0.51 | Hydroxydehydrostevic acid | 318.457 | C20H30O3 |
| 19 | 19 | 35.362 | 1.67 | 4,8,13-Cyclotetradecatriene-1,3-diol, 1,5,9-trimethyl-12-(1-methylethyl)- | 306.49 | C20H34O2 |
| 20 | 18 | 41.197 | 8.47 | Alloaromadendrene oxide-(1) | 220.3505 | C15H24O |
| 21 | 19 | 44.861 | 2.01 | Longifolenaldehyde | 220.356 | C15H24O |
| S. No. | Peak No. | Retention Time (min) | Area % | Compound Name | Molecular Weight | Molecular Formula |
|---|---|---|---|---|---|---|
| 1 | 1. | 11.553 | 0.74 | Bicyclo[2.2.1]heptan-2-one, 1,7,7-trimethyl- | 152.2334 | C10H16O |
| 2 | 2. | 12.262 | 0.28 | Bicyclo[2.2.1]heptan-2-ol, 1,7,7-trimethyl-, (1S-endo)- | 196.2860 | C12H20O2 |
| 3 | 3. | 18.197 | 1.14 | 2,4-Diisopropenyl-1-methyl-1-vinylcyclohexane | 204.351 | C15H24 |
| 4 | 4. | 18.435 | 0.52 | Hexadecane | 226.41 | C16H34 |
| 5 | 5. | 19.219 | 0.68 | .gamma.-Elemene | 204.357 | C15H24 |
| 6 | 6. | 20.743 | 0.33 | .beta.-Selinene | 204.357 | C15H24 |
| 7 | 7. | 20.832 | 0.34 | 5-Isopropenyl-3,6-dimethyl-6-vinyl-4,5,6,7-tetrahydro-1-benzofuran | 216.3187 | C15H20O |
| 8 | 8. | 20.951 | 1.19 | Heptadecane | 240.4677 | C17H36 |
| 9 | 9. | 23.346 | 3.81 | beta.-Elemenone | 218.34 | C15H22O |
| 10 | 10. | 24.041 | 0.61 | Isospathulenol | 220.356 | C15H24O |
| 11 | 11. | 25.531 | 0.64 | 3,7-Dimethyl-10-(1-methylethylidene)-3,7-cyclodecadien-1-one | 218.3346 | C15H22O |
| 12 | 12. | 27.585 | 0.86 | Cycloprop[e]indene-1a,2(1H)-dicarboxaldehyde, 3a,4,5,6,6a,6b-hexahydro-5,5,6b-trimethyl-, (1a.alpha.,3a.beta.,6a.beta.,6b.alpha | 232.323 | C15H20O2 |
| 13 | 13. | 27.745 | 1.68 | Norethindrone | 298.426 | C20H26O2 |
| 14 | 14. | 27.864 | 1.25 | (Albicanol) Decahydro-2-methylene-5,5,8a-trimethyl-1-naphthalenemethanol | 222.366 | C15H26 |
| 15 | 15. | 28.63 | 0.88 | 9-t-Butyltricyclo[4.2.1.1(2,5)]decane-9,10-diol | 224.339 | C14H24 |
| 16 | 16. | 30.031 | 0.7 | gamma.-Elemene | 204.357 | C15H24 |
| 17 | 17. | 30.387 | 0.81 | Longifolenaldehyde | 220.3505 | C15H24O |
| 18 | 18. | 31.097 | 1.46 | l-(+)-Ascorbic acid 2,6-dihexadecanoate | 652.954 | C38H68O8 |
| 19 | 19. | 33.24 | 8.32 | 2-Hydroxy-4-isopropyl-7-methoxytropone | 194.23 | C11H14O3 |
| 20 | 20. | 34.684 | 8.34 | Pregn-4,16-diene-3,20-dione dimethoxime | 314.469 | C21H30O2 |
| 21 | 21. | 35.367 | 1.24 | 4,8,13-Cyclotetradecatriene-1,3-diol, 1,5,9-trimethyl-12-(1-methylethyl)- | 306.49 | C20H34O2 |
| 22 | 22. | 36.495 | 1.9 | 2H-Cyclohepta[b]furan-2-one, 6-[1-(acetyloxy)-3-oxobutyl]-3,3a,4,7,8,8a-hexahydro-7-methyl-3-methylene- | 306.146 | C17H22O5 |
| 23 | 23 | 38.001 | 2.03 | 4,7-Methanofuro[3,2-c]oxacycloundecin-6(4H)-one, 7,8,9,12-tetrahydro-3,11-dimethyl- | 246.306 | C15H18O3 |
| 24 | 24 | 39.499 | 0.9 | Behenic alcohol | 326.6000 | C22H46O |
| 25 | 25 | 41.176 | 9.48 | 1,1,6-trimethyl-3-methylene-2-(3,6,9,13-tetramethyl-6-ethenye-10,14-dimethylene-pentadec-4-enyl)cyclohexane | 220.350 | C15H24 |
| 26 | 26 | 42.513 | 16 | 2-Phosphabicyclo[3.1.0]hex-3-ene, 6,6-dimethyl-2,3,4-triphenyl-, (endo)- | 136.194 | C8H12N |
| 27 | 27 | 43.429 | 8.67 | Cobalt, hexamethylbenzene-pentamethylcyclopentadienyl- | 329.39 | C20H30Co |
| 28 | 28 | 44.587 | 12.6 | Desmethylnomifensine | 224.307 | C15H16N2 |
| 29 | 29 | 44.871 | 8.26 | Longifolenaldehyde | 220.3505 | C15H24O |
| 30 | 30 | 46.344 | 4.34 | Furan, 2,5-bis(3,4-dimethoxyphenyl)tetrahydro-3,4-dimethyl-, [2S-(2.alpha.,3.beta.,4.alpha.,5.beta.)]- | 372.461 | C22H28O5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, S.; Verma, S.S.; Rai, V.; Awasthee, N.; Arya, J.S.; Maiti, K.K.; Gupta, S.C. Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species. Biomolecules 2019, 9, 159. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040159
Mishra S, Verma SS, Rai V, Awasthee N, Arya JS, Maiti KK, Gupta SC. Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species. Biomolecules. 2019; 9(4):159. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040159
Chicago/Turabian StyleMishra, Shruti, Sumit Singh Verma, Vipin Rai, Nikee Awasthee, Jayadev S. Arya, Kaustabh K. Maiti, and Subash C. Gupta. 2019. "Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species" Biomolecules 9, no. 4: 159. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9040159