Secretome Profiling Reveals Virulence-Associated Proteins of Fusarium proliferatum during Interaction with Banana Fruit

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Condition
2.2. Protein Extraction and Sample Preparation for a Label-Free Experiment
2.3. Protein Digestion
2.4. Liquid Chromatography (LC)-Electrospray Ionization (ESI) Tandem MS (MS/MS) Analysis by Q Exactive
2.5. Sequence Database Searching and Data Analysis
2.6. Bioinformatics Analysis
2.7. Fumonisin Production Analysis
2.8. Fusarium Proliferatum Inoculations
2.9. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.10. Statistics
3. Results
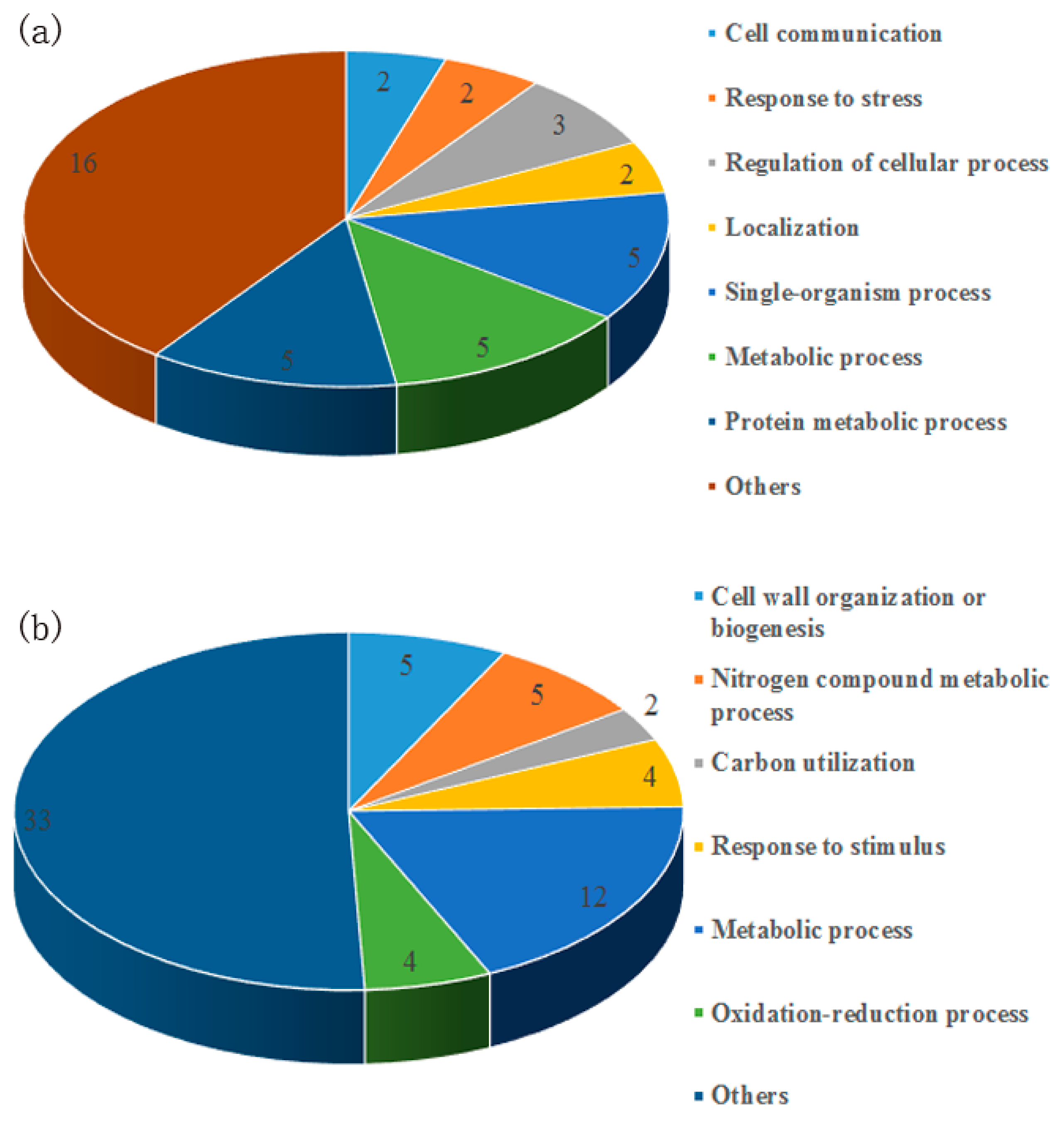
3.1. Identification of Secreted Proteins in Response to the Banana Peel
3.2. Gene Expression in Planta
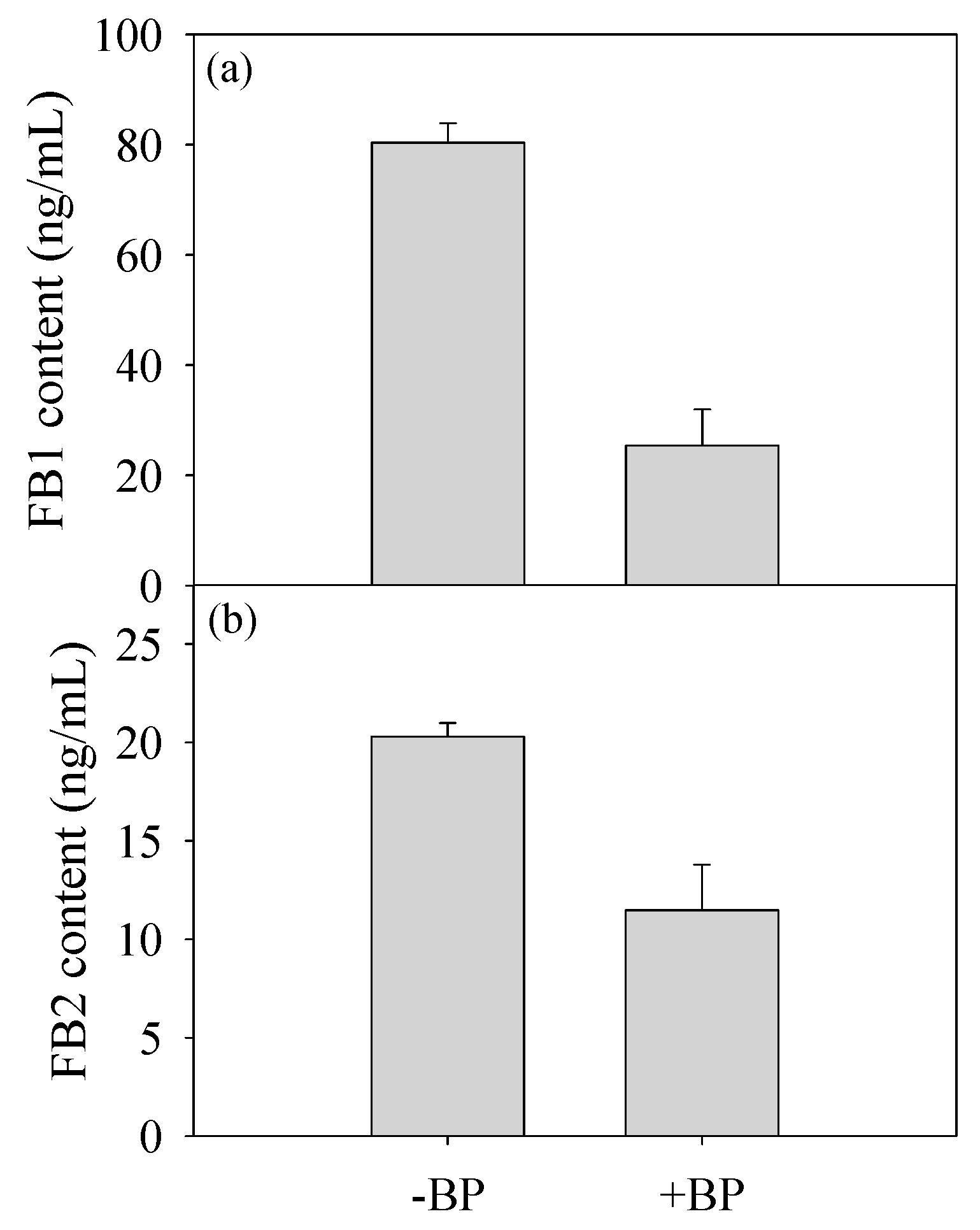
3.3. Fumonisin Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lippincott-Schwartz, J.; Roberts, T.H.; Hirschberg, K. Secretory protein trafficking and organelle dynamics in living cells. Annu. Rev. Cell Dev. Biol. 2000, 16, 557–589. [Google Scholar] [CrossRef] [PubMed]
- Mccotter, S.W.; Horianopoulos, L.C.; Kronstad, J.W. Regulation of the fungal secretome. Curr. Genet. 2016, 62, 533–545. [Google Scholar] [CrossRef]
- Christof, R.; Jacqueline, D.; Rajagopal, S.; Sean, W. Comparative secretome analysis of Fusarium graminearum and two of its non-pathogenic mutants upon deoxynivalenol induction in vitro. Proteomics 2013, 13, 1913–1921. [Google Scholar]
- Choi, J.; Park, J.; Kim, D.; Jung, K.; Kang, S.; Lee, Y.H. Fungal Secretome Database: Integrated platform for annotation of fungal secretomes. BMC Genom. 2010, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.C.; Nascimento, L.C.; Darben, L.M.; Polizel-Podanosqui, A.M.; Lopes-Caitar, V.S.; Qi, M.; Rocha, C.S.; Carazzolle, M.F.; Kuwahara, M.K.; Pereira, G.A. Prediction of the in planta P. pachyrhizi secretome and potential effector families. Mol. Plant. Pathol. 2017, 18. [Google Scholar] [CrossRef]
- Barnabas, E.L.; Ashwin, N.M.R.; Kaverinathan, K.; Trentin, A.R.; Pivato, M.; Sundar, A.R.; Malathi, P.; Viswanathan, R.; Carletti, P.; Arrigoni, G. In vitro secretomic analysis identifies putative pathogenicity-related proteins of Sporisorium scitamineum—The sugarcane smut fungus. Fungal Biol. 2017, 121, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Link, T.I.; Voegele, R.T. Secreted proteins of Uromyces fabae: Similarities and stage specificity. Mol. Plant. Pathol. 2008, 9, 59–66. [Google Scholar] [CrossRef]
- Vargas-Romero, F.; Mendoza-Hernández, G.; Suárez-Güemes, F.; Hernández-Pando, R.; Castañón-Arreola, M. Secretome profiling of highly virulent Mycobacterium bovis 04-303 strain reveals higher abundance of virulence-associated proteins. Microb. Pathog. 2016, 100, 305–311. [Google Scholar] [CrossRef]
- Yang, F.; Jensen, J.D.; Svensson, B.; Jørgensen, H.J.; Collinge, D.B.; Finnie, C. Secretomics identifies Fusarium graminearum proteins involved in the interaction with barley and wheat. Mol. Plant. Pathol. 2012, 13, 445–453. [Google Scholar] [CrossRef]
- Fen, Y.; Jensen, J.D.; Birte, S.; Jørgensen, H.J.L.; Collinge, D.B.; Christine, F. Analysis of early events in the interaction between Fusarium graminearum and the susceptible barley (Hordeum vulgare) cultivar Scarlett. Proteomics 2010, 10, 3748–3755. [Google Scholar]
- Tian, S.; Torres, R.; Ballester, A.R.; Li, B.; Vilanova, L.; González-Candelas, L. Molecular aspects in pathogen-fruit interactions: Virulence and resistance. Postharvest Biol. Technol. 2016, 122, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteomics 2014, 13, 2513. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, H.N.; Hansen, S.H.; Herbst, F.A.; Kjeldal, H.; Stensballe, A.; Nielsen, P.H.; Dueholm, M.S. Direct identification of functional amyloid proteins by Label-free quantitative mass spectrometry. Biomolecules 2017, 7, 58. [Google Scholar] [CrossRef]
- Meissner, F.; Scheltema, R.A.; Mollenkopf, H.J.; Mann, M. Direct proteomic quantification of the secretome of activated immune cells. Science 2013, 340, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Palacios, S.A.; Susca, A.; Haidukowski, M.; Stea, G.; Cendoya, E.; Ramírez, M.L.; Chulze, S.N.; Farnochi, M.C.; Moretti, A.; Torres, A.M. Genetic variability and fumonisin production by Fusarium proliferatum isolated from durum wheat grains in Argentina. Int. J. Food Microbiol. 2015, 201, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Özer, G.; Bayraktar, H.; Oksal, E. First report of Fusarium proliferatum causing crown and root rot of Asparagus officinalis in Turkey. J. Plant. Pathol. 2011, 93, S88–S88. [Google Scholar]
- Jing, L.; Jiang, G.; Bao, Y.; Dong, X.; Feng, L.; Lin, S.; Feng, C.; Ashraf, M.; Jiang, Y. A luminescent bacterium assay of fusaric acid produced by Fusarium proliferatum from banana. Anal. Bioanal. Chem. 2012, 402, 1347–1354. [Google Scholar]
- Kamel, M.A.M.; Cortesi, P.; Saracchi, M. Etiological agents of crown rot of organic bananas in Dominican Republic. Postharvest Biol. Technol. 2016, 120, 112–120. [Google Scholar] [CrossRef]
- Li, T.; Gong, L.; Jiang, G.; Wang, Y.; Gupta, V.K.; Qu, H.; Duan, X.; Wang, J.; Jiang, Y. Carbon Sources Influence Fumonisin Production in Fusarium proliferatum. Proteomics 2017, 17, 1700070. [Google Scholar] [CrossRef]
- Li, T.; Jian, Q.; Wang, Y.; Chen, F.; Yang, C.; Gong, L.; Duan, X.; Yang, B.; Jiang, Y. Inhibitory mechanism of butylated hydroxyanisole against infection of Fusarium proliferatum based on comparative proteomic analysis. J. Proteomics 2016, 148, 1–11. [Google Scholar] [CrossRef]
- Li, T.; Wu, Q.; Yong, W.; John, A.; Qu, H.; Liang, G.; Duan, X.; Hong, Z.; Yun, Z.; Jiang, Y. Application of proteomics for the investigation of the effect of initial pH on pathogenic mechanisms of Fusarium proliferatum on banana fruit. Front. Microbiol. 2017, 8, 2327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ni, H.; Du, X.; Wang, S.; Ma, X.W.; Nã¼Rnberger, T.; Guo, H.S.; Hua, C. The Verticillium-specific protein VdSCP7 localizes to the plant nucleus and modulates immunity to fungal infections. New Phytol. 2017, 215, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; Ver Loren van Themaat, E.; van der Does, H.C.; Hacquard, S.; Stüber, K.; Will, I.; Schmalenbach, W.; et al. Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, U359–U360. [Google Scholar] [CrossRef] [PubMed]
- Thomas Nordahl, P.; Soren, B.; Gunnar, V.H.; Henrik, N. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Kiemer, L.; Fausboll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5. [Google Scholar] [CrossRef]
- Lum, G.; Min, X.J. FunSecKB: The Fungal secretome knowledgeBase. Database-Oxford 2011. [Google Scholar] [CrossRef]
- Meijer, H.J.G.; Mancuso, F.M.; Espadas, G.; Seidl, M.F.; Chiva, C.; Govers, F.; Sabido, E. Profiling the secretome and extracellular proteome of the potato late blight pathogen Phytophthora infestans. Mol. Cell. Proteomics 2014, 13, 2101–2113. [Google Scholar] [CrossRef]
- Shan, W.; Chen, J.Y.; Kuang, J.F.; Lu, W.J. Banana fruit NAC transcription factor MaNAC5 cooperates with MaWRKYs to enhance the expression of pathogenesis-related genes against Colletotrichum musae. Mol. Plant. Pathol. 2016, 17, 330–338. [Google Scholar] [CrossRef]
- Mueller, O.; Kahmann, R.; Aguilar, G.; Trejo-Aguilar, B.; Wu, A.; de Vries, R.P. The secretome of the maize pathogen Ustilago maydis. Fungal Genet. Biol. 2008, 45 (Suppl. S1), S63–S70. [Google Scholar] [CrossRef]
- Sang, G.K.; Wang, Y.; Lee, K.H.; Park, Z.Y.; Park, J.; Wu, J.; Kwon, S.J.; Lee, Y.H.; Agrawal, G.K.; Rakwal, R. In-depth insight into in vivo apoplastic secretome of rice-Magnaporthe oryzae interaction. J. Proteomics 2013, 78, 58–71. [Google Scholar]
- Meinhardt, L.W.; Costa, G.G.L.; Thomazella, D.P.; Teixeira, P.J.P.; Carazzolle, M.F.; Schuster, S.C.; Carlson, J.E.; Guiltinan, M.J.; Mieczkowski, P.; Farmer, A. Genome and secretome analysis of the hemibiotrophic fungal pathogen, Moniliophthora roreri, which causes frosty pod rot disease of cacao: Mechanisms of the biotrophic and necrotrophic phases. BMC Genom. 2014, 15, 164. [Google Scholar] [CrossRef] [PubMed]
- Paper, J.M.; Scott-Craig, J.S.; Adhikari, N.D.; Cuomo, C.A.; Walton, J.D. Comparative proteomics of extracellular proteins in vitro and in planta from the pathogenic fungus Fusarium graminearum. Proteomics 2007, 7, 3171–3183. [Google Scholar] [CrossRef] [PubMed]
- Phalip, V.; Delalande, F.; Carapito, C.; Goubet, F.; Hatsch, D.; Leize-Wagner, E.; Dupree, P.; Dorsselaer, A.V.; Jeltsch, J.M. Diversity of the exoproteome of Fusarium graminearum grown on plant cell wall. Curr. Genet. 2005, 48, 366–379. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sadygov, R.G.; Yates, J.R., 3rd. A model for random sampling and estimation of relative protein abundance in shotgun proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef] [PubMed]
- Rabouille, C. Pathways of unconventional protein secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef]
- Pancholi, V.; Chhatwal, G.S. Housekeeping enzymes as virulence factors for pathogens. Int J. Med. Microbiol. 2003, 293, 391–401. [Google Scholar] [CrossRef]
- Artier, J.; Da, S.Z.F.; Fm, D.S.C.; Pauletti, B.A.; Leme, A.F.; Carnielli, C.M.; Selistre-De-Araujo, H.S.; Bertolini, M.C.; Ferro, J.A.; Belasque, J.J. Comparative proteomic analysis of Xanthomonas citri ssp. citri periplasmic proteins reveals changes in cellular envelope metabolism during in vitro pathogenicity induction. Mol. Plant. Pathol. 2016, 19, 143–157. [Google Scholar] [CrossRef]
- Mulugeta, N.; Linda, D.; Melanie, K.; Anne-Kathrin, Z.; Patrick, E.; Dirk, A.; Bernhard, K.; Ralf, R.; Michael, H.; Friedrich, G.T. Secretome analysis revealed adaptive and non-adaptive responses of the Staphylococcus carnosus femB mutant. Proteomics 2015, 15, 1268–1279. [Google Scholar]
- Lu, H.; Xiong, J.; Shang, Q.; Jiang, Y.; Cao, Y. Roles of RPS41 in biofilm formation, virulence, and hydrogen peroxide sensitivity in Candida albicans. Curr. Microbiol. 2016, 72, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.; Pant, R.; Raghunath, A.; Irvine, A.G.; Pedro, H.; Hammond-Kosack, K.E. The Pathogen-Host Interactions database (PHI-base): Additions and future developments. Nucleic Acids Res. 2015, 43, D645–D655. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.S.; Flaishman, M.A.; Kolattukudy, P.E. Cloning of a gene expressed during appressorium formation by Colletotrichum gloeosporioides and a marked decrease in virulence by disruption of this gene. Plant. Cell 1995, 7, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, J.; Jia, L.J.; Yuan, T.L.; Zhang, D.; Guo, Y.; Wang, Y.; Tang, W.H. Cellular tracking and gene profiling of Fusarium graminearum during maize stalk rot disease development elucidates its strategies in confronting phosphorus limitation in the host apoplast. PLoS Pathog. 2016, 12, e1005485. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Kim, S.G.; Tsuda, K.; Gupta, R.; Park, S.Y.; Kim, S.T.; Kang, K.Y. Magnaporthe oryzae-secreted protein MSP1 induces cell death and elicits defense responses in rice. Mol. Plant. Microbe. Interact. 2016, 29, 299. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.; Schafer, W.; Salomon, S. A secreted lipase of Fusarium graminearum is a virulence factor required for infection of cereals. Plant. J. 2010, 42, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Vickers, I.; Reeves, E.P.; Kavanagh, K.A.; Doyle, S. Isolation, activity and immunological characterisation of a secreted aspartic protease, CtsD, from Aspergillus fumigatus. Protein Expr. Purif. 2007, 53, 216–224. [Google Scholar] [CrossRef]
- Li, B.Q.; Wang, W.H.; Zong, Y.Y.; Qin, G.Z.; Tian, S.P. Exploring pathogenic mechanisms of Botrytis cinerea secretome under different ambient pH based on comparative proteomic analysis. J. Proteome Res. 2012, 11, 4249–4260. [Google Scholar] [CrossRef]
- Boldo, J.T.; Junges, A.; Amaral, K.B.D.; Staats, C.C.; Vainstein, M.H.; Schrank, A. Endochitinase CHI2 of the biocontrol fungus Metarhizium anisopliae affects its virulence toward the cotton stainer bug Dysdercus peruvianus. Curr. Genet. 2009, 55, 551–560. [Google Scholar] [CrossRef]
- Chang, Q.; Liu, J.; Lin, X.; Hu, S.; Yang, Y.; Li, D.; Chen, L.; Huai, B.; Huang, L.; Voegele, R.T. A unique invertase is important for sugar absorption of an obligate biotrophic pathogen during infection. New Phytol. 2017, 215, 1548. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houterman, P.M.; Cornelissen, B.J.C.; Rep, M. Suppression of plant resistance gene-based immunity by a fungal effector. PLoS Pathog. 2008, 4, e1000061. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal Effector Proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloppholz, S.; Kuhn, H.; Requena, N. A secreted fungal effector of Glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Krijger, J.J.; Horbach, R.; Behr, M.; Schweizer, P.; Deising, H.B.; Wirsel, S.G.R. The yeast signal sequence trap identifies secreted proteins of the hemibiotrophic corn pathogen Colletotrichum graminicola. Mol. Plant. Microbe Interact. 2008, 21, 1325–1336. [Google Scholar] [CrossRef]
- Son, H.; Seo, Y.S.; Min, K.; Park, A.R.; Lee, J.; Jin, J.M.; Lin, Y.; Cao, P.J.; Hong, S.Y.; Kim, E.K.; et al. A Phenome-Based Functional Analysis of Transcription Factors in the Cereal Head Blight Fungus, Fusarium graminearum. PLoS Pathog. 2011, 7, e1002310. [Google Scholar] [CrossRef]
- Wee, J.; Hong, S.Y.; Roze, L.V.; Day, D.M.; Chanda, A.; Linz, J.E. The fungal bZIP transcription factor AtfB controls virulence-associated processes in Aspergillus parasiticus. Toxins 2017, 9, 287. [Google Scholar] [CrossRef]
- Gelderblom, W.C.A.; Jaskiewicz, K.; Marasas, W.F.O.; Thiel, P.G.; Horak, R.M.; Vleggaar, R.; Kriek, N.P.J. Fumonisins - novel mycotoxins with cancer-promoting activity produced by Fusarium-moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar]
- Picot, A.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Lannou, C.; Richard-Forget, F. Factors of the Fusarium verticillioides-maize environment modulating fumonisin production. Crit Rev. Microbiol. 2010, 36, 221–231. [Google Scholar] [CrossRef]
- Glenn, A.E.; Zitomer, N.C.; Zimeri, A.M.; Williams, L.D.; Riley, R.T.; Proctor, R.H. Transformation-mediated complementation of a FUM gene cluster deletion in Fusarium verticillioides restores both fumonisin production and pathogenicity on maize seedlings. Mol. Plant. Microbe Interact. 2008, 21, 87–97. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Munkvold, G.P.; Plattner, R.D.; Proctor, R.H. FUM1—A gene required for fumonisin biosynthesis but not for maize ear rot and ear infection by Gibberella moniliformis in field tests. Mol. Plant. Microbe Interact. 2002, 15, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Busman, M.; Muhitch, M.; Proctor, R.H. Complementary host-pathogen genetic analyses of the role of fumonisins in the Zea mays-Gibberella moniliformis interaction. Physiol. Mol. Plant. Pathol. 2007, 70, 149–160. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein IDs | Protein Description | Unique Peptides | Mr (kDa) | Signal | NLS Sequence |
|---|---|---|---|---|---|
| CZR33670.1 | APT1-adenine phosphoribosyltransferase | 1 | 23.3 | P | |
| CZR34854.1 | uncharacterized protein FPRO_01025 | 4 | 96.7 | P | |
| CZR34878.1 | uncharacterized protein FPRO_01001 | 2 | 77.3 | P | |
| CZR35312.1 | uncharacterized protein FPRO_00565 | 2 | 24.0 | S | |
| CZR37023.1 | uncharacterized protein FPRO_02717 | 2 | 26.4 | S | LFRTIASVAFLACASNVAAEPQPYKLVKAP |
| CZR37404.1 | uncharacterized protein FPRO_02336 | 1 | 19.7 | S | |
| CZR37761.1 | uncharacterized protein FPRO_07048 | 5 | 20.8 | S | |
| CZR37893.1 | uncharacterized protein FPRO_06916 | 3 | 9.5 | S | RSILHHCGKHASWDHAKSECVCHDSGKVYTKKHH |
| CZR37952.1 | uncharacterized protein FPRO_06857 | 2 | 25.8 | T | |
| CZR38705.1 | CAP20-virulence factor | 6 | 20.3 | P | |
| CZR39121.1 | uncharacterized protein FPRO_05687 | 1 | 26. 3 | S | |
| CZR39201.1 | uncharacterized protein FPRO_05607 | 1 | 39.0 | S | |
| CZR39716.1 | H+-transporting ATPase | 2 | 85.7 | P | |
| CZR39800.1 | 60S ribosomal protein L8 | 3 | 27.6 | P | |
| CZR39939.1 | ATP-dependent RNA helicase DED1 | 1 | 71.8 | P | |
| CZR40877.1 | related to SYP1 Protein with a potential role in actin cytoskeletal organization | 3 | 96.3 | P | |
| CZR41198.1 | related to aspartic proteinase OPSB | 5 | 49.9 | S | RRRLRKRDGTIEIGIDNEQSLYFLNASLGTPP |
| CZR41331.1 | 26S proteasome regulatory subunit RPN8 | 2 | 38.3 | P | |
| CZR41485.1 | GTP-binding protein ypt1 | 4 | 22.4 | P | |
| CZR42158.1 | related to ochre suppressor tyr-tRNA | 2 | 18.1 | P | RNEVIRCLREHDRRPLDCWQEVENFKAEVKKLEKSW |
| CZR42368.1 | transcription factor BTF3a | 4 | 17.0 | P | PRRKVKRAPARSGADDKKLQLALKKLNT |
| CZR42437.1 | woronin body major protein precursor | 3 | 71.0 | P | |
| CZR42601.1 | uncharacterized protein FPRO_09904 | 2 | 15.3 | S | SKRQIVWPAYTDKQVQSGKVVKPD |
| CZR43413.1 | related to lactonohydrolase | 1 | 35.6 | P | |
| CZR43528.1 | related to 2‘-hydroxyisoflavone reductase | 2 | 32.1 | P | |
| CZR44148.1 | potassium channel beta subunit protein | 3 | 39.5 | P | |
| CZR45692.1 | related to tripeptidyl-peptidase I | 6 | 64.9 | T | |
| CZR46340.1 | related to protocatechuate 3,4-dioxygenase beta subunit | 1 | 39.8 | S | |
| CZR46385.1 | related to oxidoreductase related to nitroreductase | 3 | 22.6 | P | |
| CZR46670.1 | uncharacterized protein FPRO_12120 | 1 | 9.7 | P | |
| CZR46904.1 | cytochrome-c oxidase chain IV precursor | 3 | 27.0 | P | |
| CZR46998.1 | related to tripeptidyl-peptidase I | 6 | 64.9 | S | |
| CZR47100.1 | transcriptional repressor rco-1 | 2 | 66.1 | P | LDRTIKMWELSAPRQGNQPGPKGGKCVKT |
| CZR47653.1 | related to acetylxylan esterase precursor | 2 | 30.8 | T | |
| CZR47873.1 | uncharacterized protein FPRO_13540 | 1 | 35.5 | S | |
| CZR48663.1 | related to toxD protein | 5 | 38.3 | P | |
| CZR48742.1 | lipase precursor | 6 | 47.5 | S | |
| CZR48857.1 | related to triacylglycerol lipase V precursor | 4 | 57.3 | S | |
| CZR49188.1 | zuotin | 1 | 50.4 | P | ENRDQKRHQERKNTNARKKKKAD |
| CZR49277.1 | uncharacterized protein FPRO_08983 | 4 | 19.9 | S |
| Protein IDs | Protein Description | Unique Peptides | Mr. (kDa) | Signal | Ratio (+BP/-BP) | NLS Sequence |
|---|---|---|---|---|---|---|
| CZR47323.1 | uncharacterized protein FPRO_08697 | 6 | 33.5 | S | 14.6 | |
| CZR45923.1 | related to beta-glucosidase 1 precursor | 28 | 83.4 | S | 16.3 | |
| CZR44287.1 | uncharacterized protein FPRO_14048 | 3 | 23.2 | S | 8.6 | |
| CZR49124.1 | uncharacterized protein FPRO_12560 | 5 | 37.7 | S | 7.2 | PSSKRGLIYIPNSDFPSDDKVWVQKHSDLT |
| CZR35354.1 | probable 1,4-Benzoquinone reductase | 7 | 21.7 | P | 7.7 | |
| CZR43990.1 | related to glucan 1,3-beta-glucosidase | 9 | 33.5 | S | 8.4 | |
| CZR41742.1 | uncharacterized protein FPRO_11332 | 12 | 93.7 | S | 22.7 | |
| CZR46647.1 | uncharacterized protein FPRO_12097 | 5 | 16.1 | S | 5.8 | DTVGKHFIPNKQLWQSKEPNAEIQRYKGPKD |
| CZR44412.1 | related to triacylglycerol lipase V precursor | 21 | 65.1 | S | 12.1 | |
| CZR47104.1 | probable malate dehydrogenase | 14 | 34.9 | P | 3.1 | |
| CZR38616.1 | probable glyceraldehyde 3-phosphate dehydrogenase (ccg-7) | 20 | 36.1 | P | 114.7 | |
| CZR45734.1 | related to acid phosphatase Pho610 | 8 | 48.7 | S | 3.0 | |
| CZR33506.1 | uncharacterized protein FPRO_01717 | 5 | 23.8 | S | 2.2 | |
| CZR46837.1 | probable FBA1-fructose-bisphosphate aldolase | 13 | 39.6 | P | 3.7 | |
| CZR36235.1 | related to phosphatidylcholine-sterol acyltransferase precursor | 4 | 32.6 | S | 7.2 | |
| CZR47504.1 | uncharacterized protein FPRO_1317 | 11 | 20.7 | S | 2.5 | EKTWKNAHYKAGGDKAYSNRRVTCQQKQLKVP |
| CZR47726.1 | related to glu/asp-tRNA amidotransferase subunit A | 21 | 63.4 | S | 8.2 | DAPSKRRLPK |
| CZR45085.1 | probable rAsp f 9 allergen | 12 | 41.3 | S | 21.9 | WSKIALAGLFASAAAQTYSECNPMKKTCDP |
| CZR44035.1 | uncharacterized protein FPRO_13841 | 1 | 28.4 | S | 11.1 | |
| CZR35784.1 | uncharacterized protein FPRO_00093 | 1 | 20.9 | S | 5.5 | |
| CZR47211.1 | related to acetylxylan esterase precursor | 7 | 36.1 | S | 4.3 | |
| CZR46230.1 | uncharacterized protein FPRO_11677 | 8 | 19.1 | S | 12.4 | |
| CZR36609.1 | related to tyrosinase precursor | 11 | 62.9 | S | 2.7 | |
| CZR45243.1 | related to lipase (lipP) | 3 | 34.6 | P | 2.3 | |
| CZR47062.1 | uncharacterized protein FPRO_08436 | 1 | 14.8 | S | 5.1 | |
| CZR40146.1 | uncharacterized protein FPRO_05046 | 4 | 14.3 | S | 12.9 | |
| CZR42211.1 | ribosomal protein L7a | 6 | 29.8 | P | 25.2 | |
| CZR45507.1 | cytochrome P450 55A2 | 16 | 46.9 | P | 2.3 | |
| CZR37840.1 | SnodProt1 precursor | 3 | 14.6 | S | 8.9 | |
| CZR38203.1 | uncharacterized protein FPRO_06606 | 1 | 11.9 | S | 77.4 | |
| CZR42089.1 | endochitinase 2 precursor | 12 | 88.5 | P | 2.8 | |
| CZR48911.1 | subtilisin-like serine protease | 26 | 92.2 | S | 2.1 | |
| CZR40535.1 | related to amidase family protein | 16 | 70.1 | S | 7.8 | |
| CZR42124.1 | uncharacterized protein FPRO_09425 | 6 | 19.2 | S | 3.4 | |
| CZR43152.1 | related to sporulation-specific gene SPS2 | 13 | 42.3 | S | 2.4 | |
| CZR40180.1 | uncharacterized protein FPRO_05080 | 2 | 14.8 | S | 6.5 | |
| CZR42998.1 | uncharacterized protein FPRO_08086 | 4 | 16.8 | S | 8.9 | |
| CZR38140.1 | uncharacterized protein FPRO_06669 | 4 | 32.1 | S | 5.8 | |
| CZR38273.1 | fusarubin cluster-esterase | 12 | 41.2 | S | 5.7 | |
| CZR36329.1 | uncharacterized protein FPRO_03411 | 1 | 23.4 | S | 11.0 | |
| CZR47769.1 | phosphoglycerate kinase | 31 | 44.7 | P | 2.1 | |
| CZR35108.1 | uncharacterized protein FPRO_00770 | 6 | 21.5 | S | 7.2 | |
| CZR40410.1 | uncharacterized protein FPRO_05310 | 3 | 18.4 | P | 11.1 | |
| CZR38701.1 | uncharacterized protein FPRO_06108 | 3 | 30.9 | S | 6.7 | |
| CZR44031.1 | uncharacterized protein FPRO_13838 | 2 | 79.7 | S | 3.9 | |
| CZR48068.1 | uncharacterized protein FPRO_12678 | 5 | 32.5 | S | 3.9 | |
| CZR38172.1 | related to SUC2-invertase (sucrose hydrolyzing enzyme) | 10 | 60.2 | S | 2.9 | |
| CZR49275.1 | uncharacterized protein FPRO_08985 | 4 | 28.0 | S | 6.2 | |
| CZR49368.1 | related to BNR/Asp-box repeat domain protein | 4 | 41.3 | S | 3.9 | |
| CZR34777.1 | related to extracellular matrix protein precursor | 4 | 21.9 | S | 3.5 | |
| CZR34851.1 | uncharacterized protein FPRO_01028 | 5 | 15.3 | S | 3.8 | |
| CZR36412.1 | related to myosin heavy chain | 18 | 133.6 | P | 8.3 | |
| CZR34562.1 | probable NHP6B-nonhistone chromosomal protein | 2 | 11.5 | P | 7.6 | |
| CZR44986.1 | related to endo-1,3-beta-glucanase | 4 | 33.1 | S | 3.3 | |
| CZR41328.1 | related to serine proteinase inhibitor IA-2 | 5 | 10.5 | S | 6.1 | |
| CZR41607.1 | uncharacterized protein FPRO_11196 | 5 | 22.3 | S | 2.5 | |
| CZR49589.1 | uncharacterized protein FPRO_15947 | 12 | 58.9 | S | 69.2 | |
| CZR42484.1 | related to glucan 1,3-beta-glucosidase | 13 | 94.1 | S | 2.1 | |
| CZR49007.1 | uncharacterized protein FPRO_12444 | 3 | 26.1 | S | 2.7 | |
| CZR41020.1 | uncharacterized protein FPRO_10609 | 1 | 34.3 | S | 4.4 | |
| CZR47343.1 | related to acid phosphatase precursor (pH 6-optimum acid phosphatase) | 3 | 70.1 | S | 8.5 | |
| CZR34748.1 | uncharacterized protein FPRO_01131 | 6 | 19.2 | S | 4.2 | |
| CZR45488.1 | pectinesterase precursor | 7 | 34.9 | S | 2.3 | |
| CZR39344.1 | uncharacterized protein FPRO_04241 | 2 | 27.4 | S | 3.8 | |
| CZR42912.1 | CPC2 protein | 5 | 35.0 | P | 2.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Wu, Y.; Wang, Y.; Gao, H.; Gupta, V.K.; Duan, X.; Qu, H.; Jiang, Y. Secretome Profiling Reveals Virulence-Associated Proteins of Fusarium proliferatum during Interaction with Banana Fruit. Biomolecules 2019, 9, 246. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060246
Li T, Wu Y, Wang Y, Gao H, Gupta VK, Duan X, Qu H, Jiang Y. Secretome Profiling Reveals Virulence-Associated Proteins of Fusarium proliferatum during Interaction with Banana Fruit. Biomolecules. 2019; 9(6):246. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060246
Chicago/Turabian StyleLi, Taotao, Yu Wu, Yong Wang, Haiyan Gao, Vijai Kumar Gupta, Xuewu Duan, Hongxia Qu, and Yueming Jiang. 2019. "Secretome Profiling Reveals Virulence-Associated Proteins of Fusarium proliferatum during Interaction with Banana Fruit" Biomolecules 9, no. 6: 246. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060246