Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and Its Truncated Analogues

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Preparation and Secretion Harvesting
2.2. “Shotgun” Cloning of a Novel Ranatuerin-2Pb Encoded cDNA Library
2.3. Peptide Synthesis
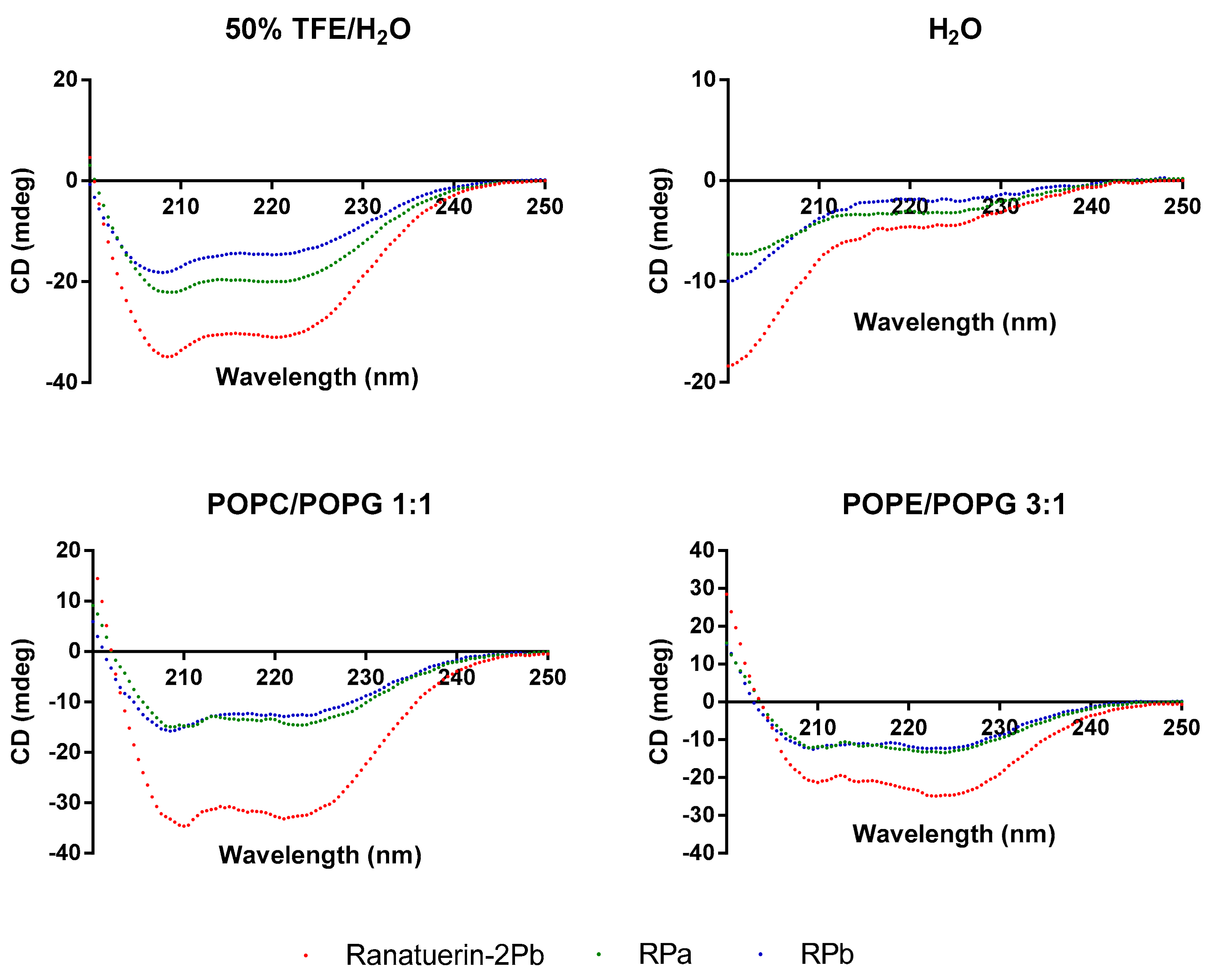
2.4. Conformation Study
2.5. Antimicrobial Activity
2.6. Antibiofilm Assays with Different Organisms
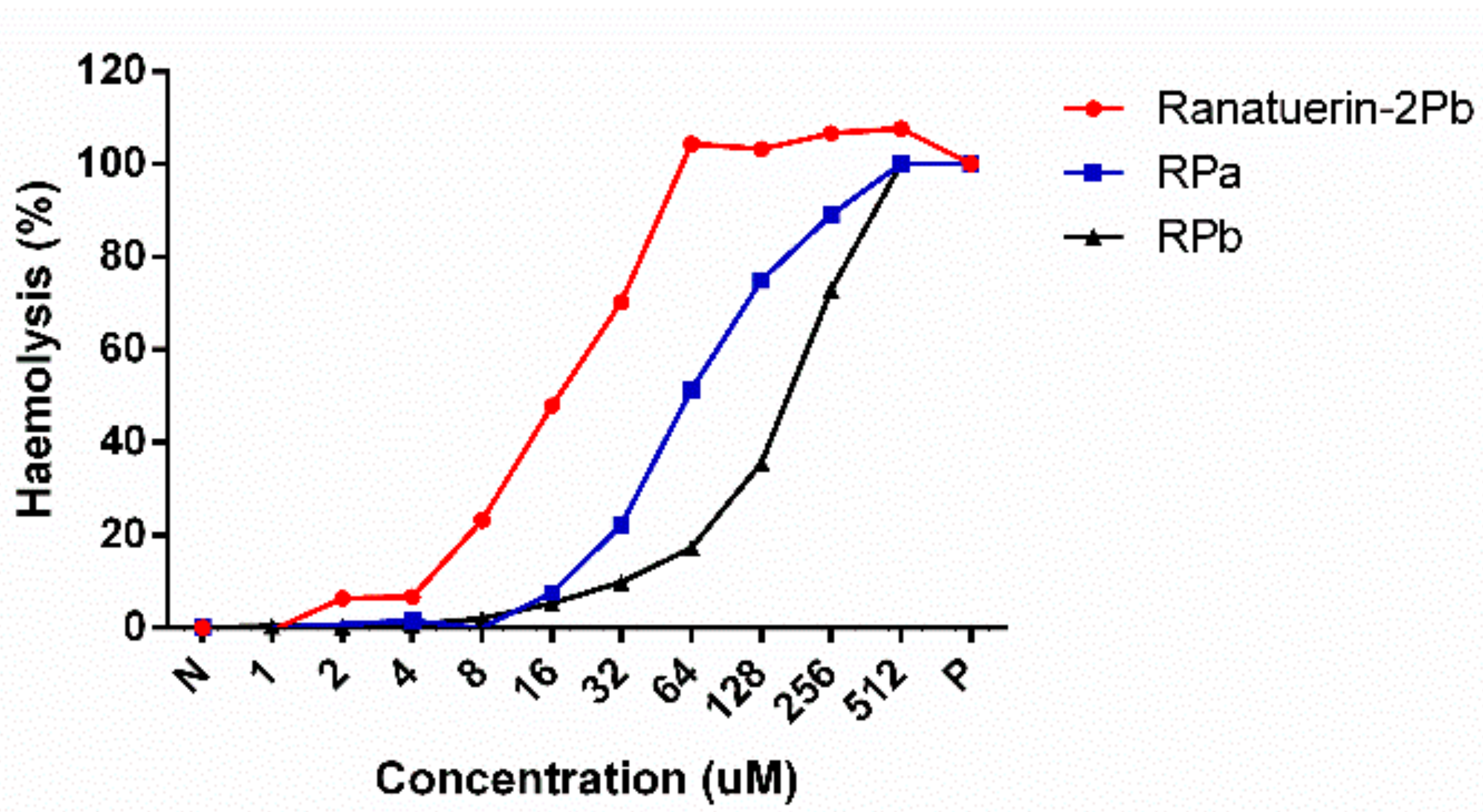
2.7. Haemolysis Assay
2.8. Membrane Permeability Assay
2.9. MTT and Lactate Dehydrogenase (LDH) Cytotoxicity Assay
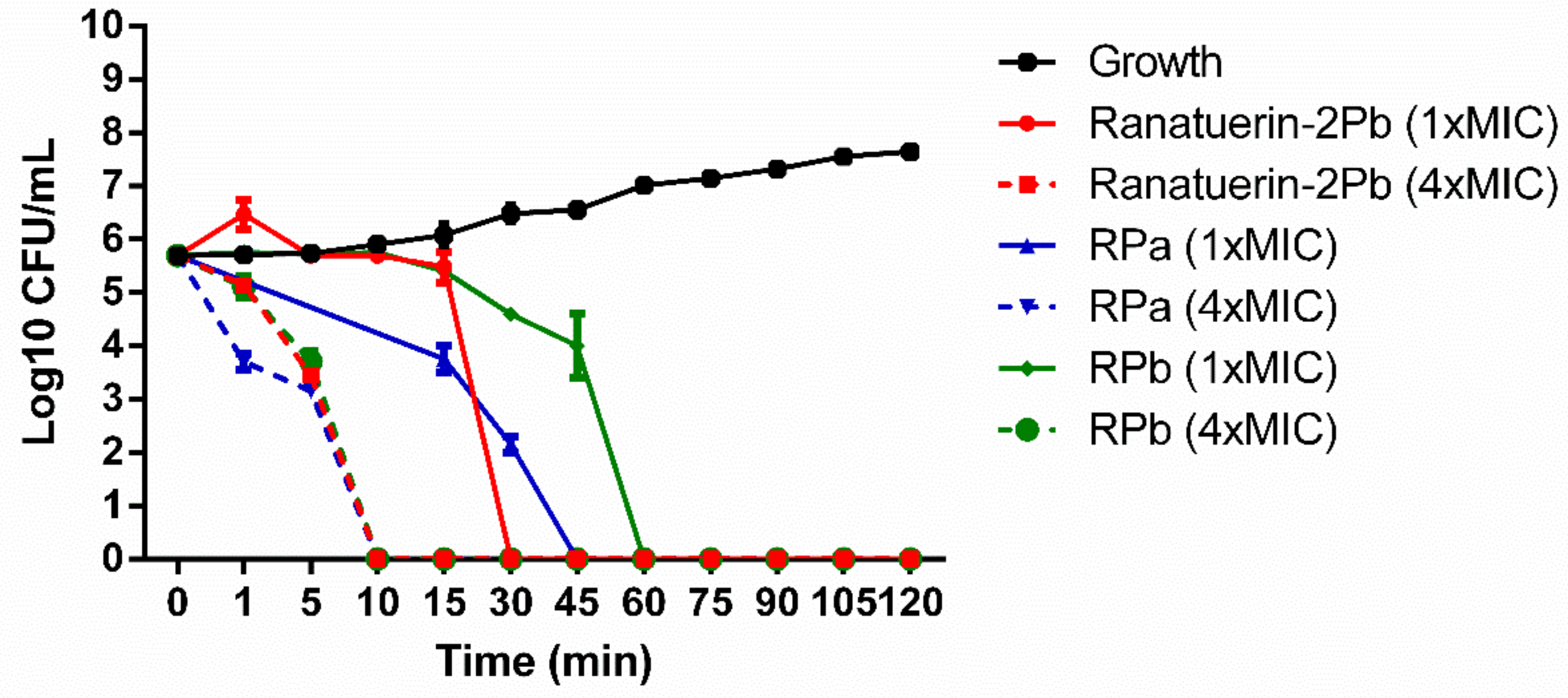
2.10. Time-Kill Assay
2.11. Assessing Efficacy of Peptides Against MRSA In Vivo
2.12. Statistical Analysis
3. Results
3.1. The Translated Open-Reading Frame Amino Acids Sequences of Ranatuerin-2 Peptides
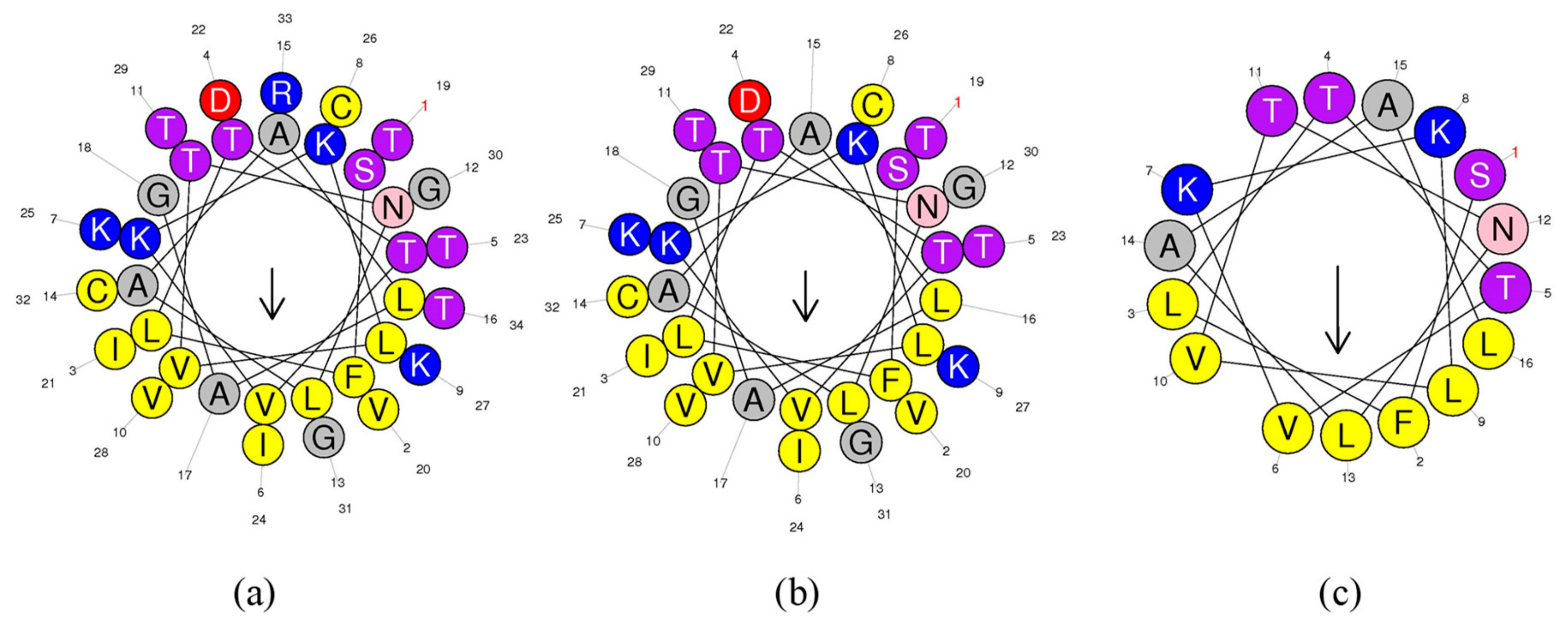
3.2. Peptide Design
3.3. Secondary Structure Analysis
3.4. Antimicrobial and Haemolytic Activity of Peptides
3.5. Time-Kill Assay against S. aureus of Peptides
3.6. Antibiofilm Assay of Peptides against S. aureus
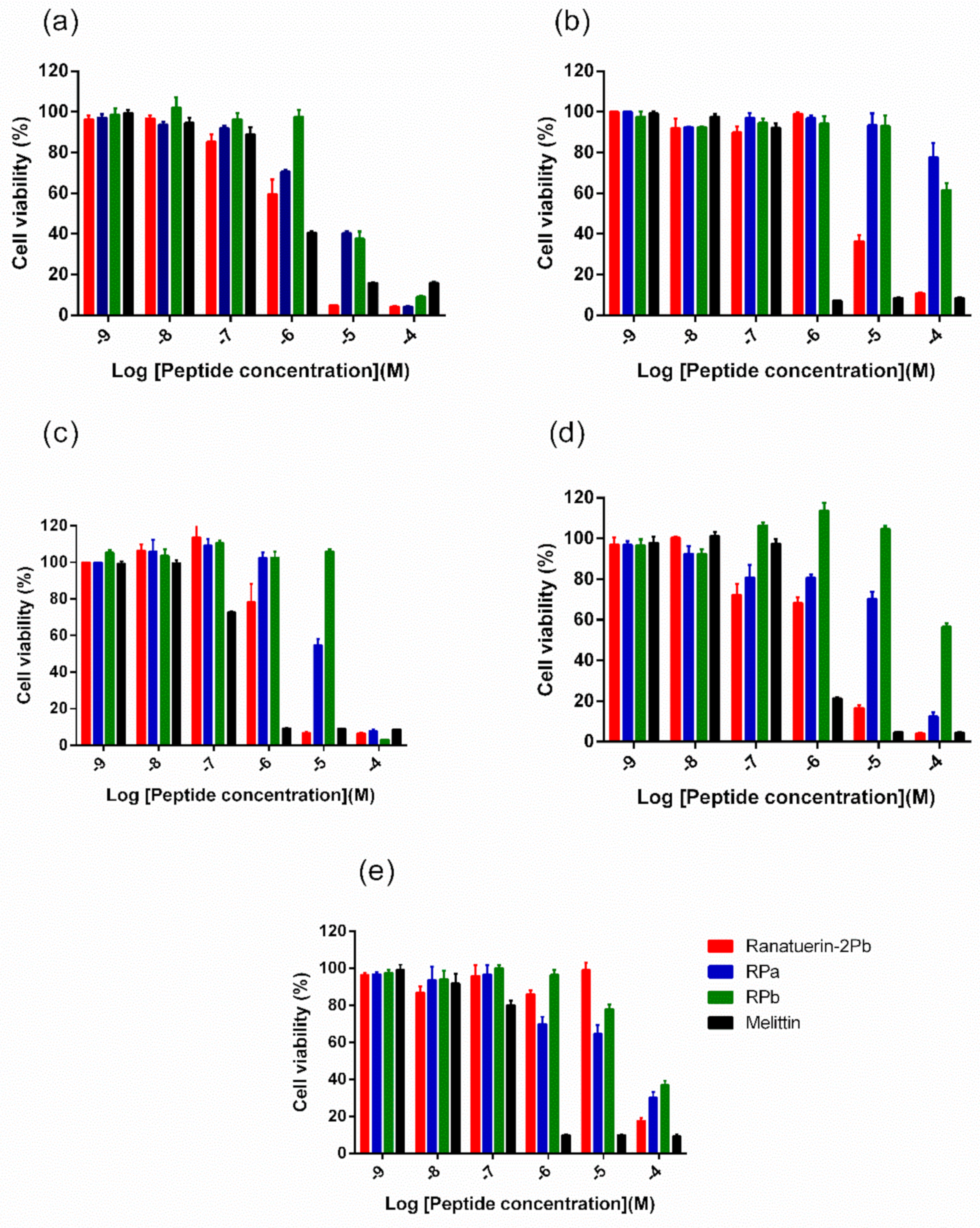
3.7. MTT Cell Viability Assay
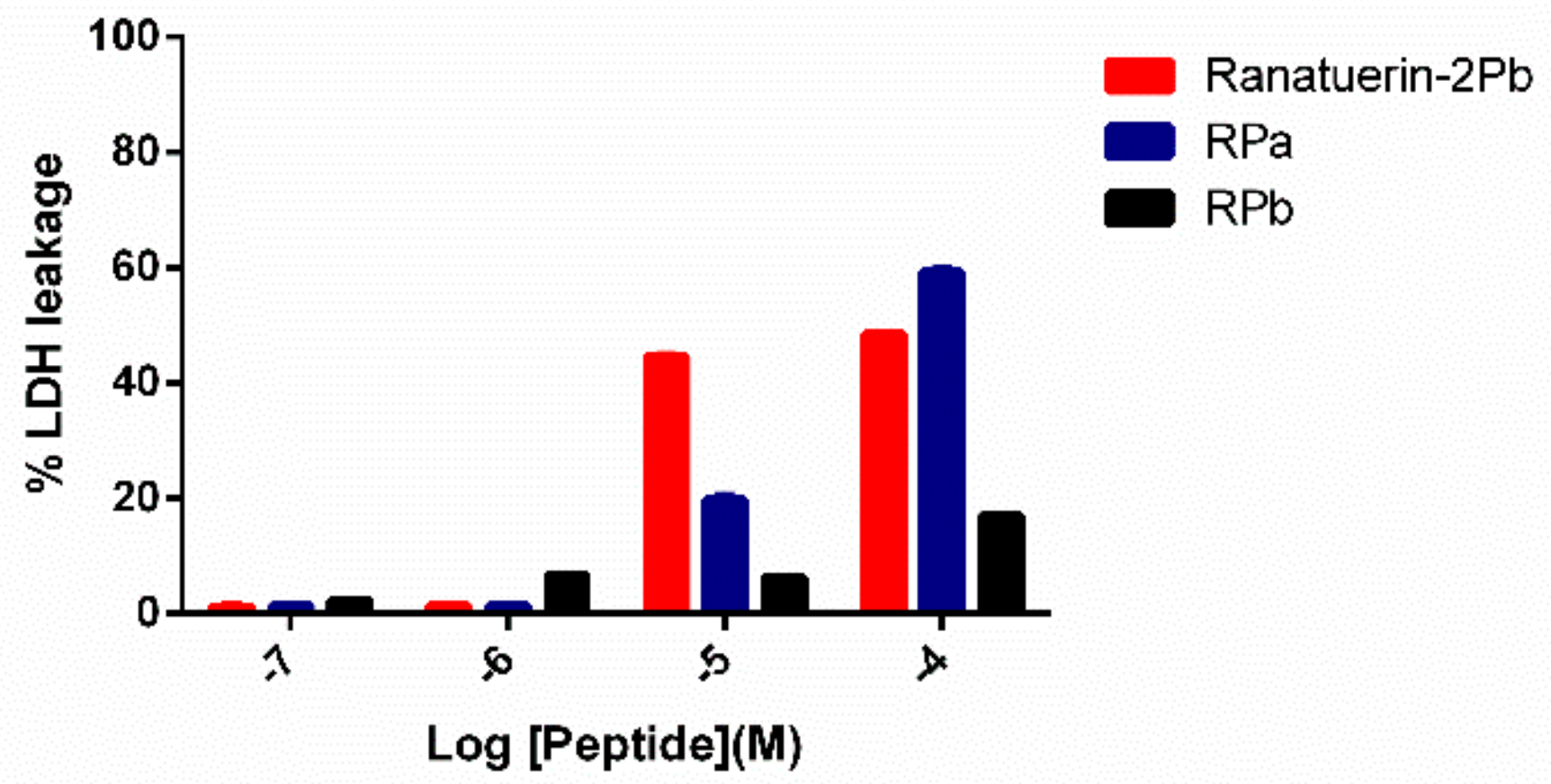
3.8. LDH Assay
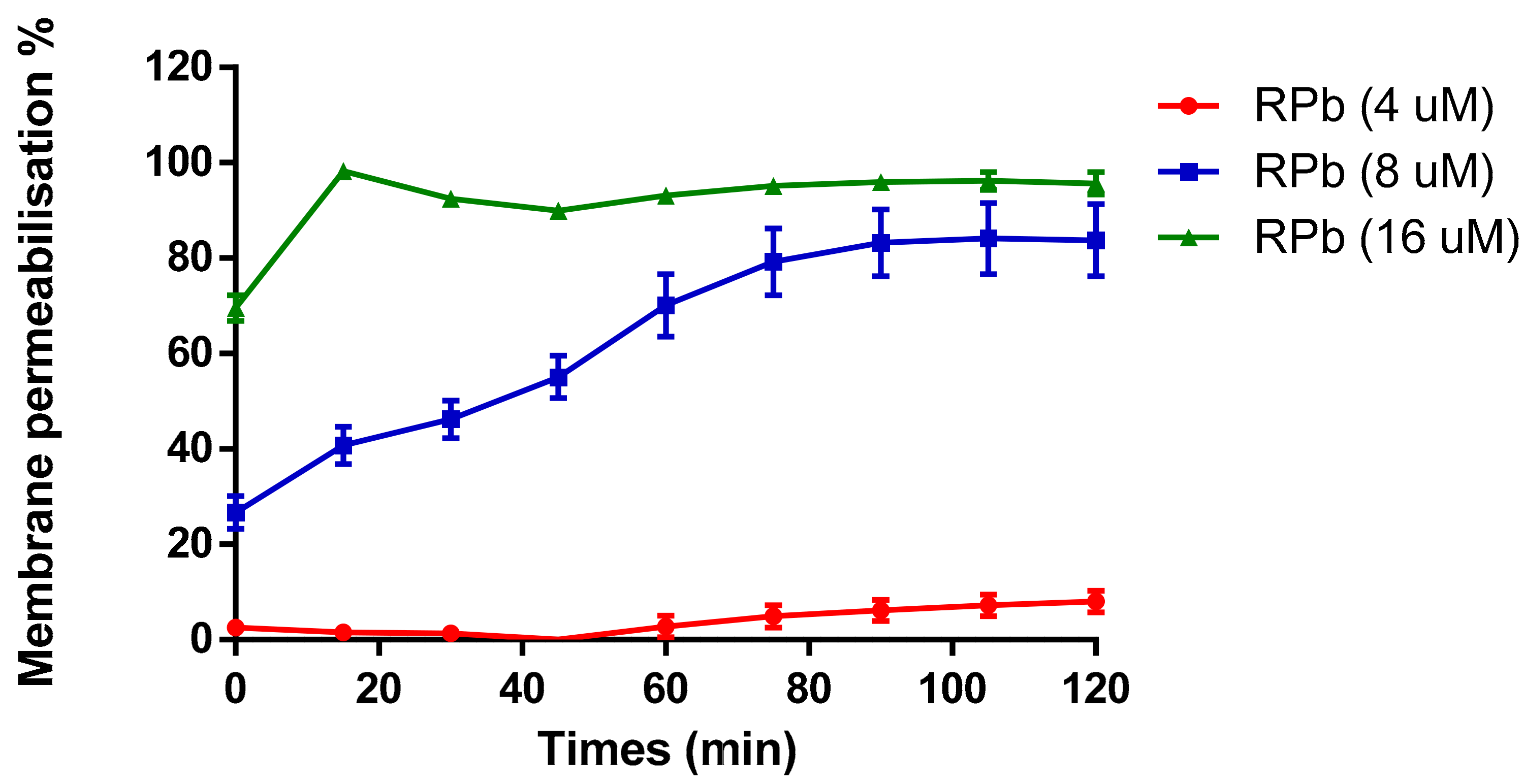
3.9. Membrane Permeability Assay
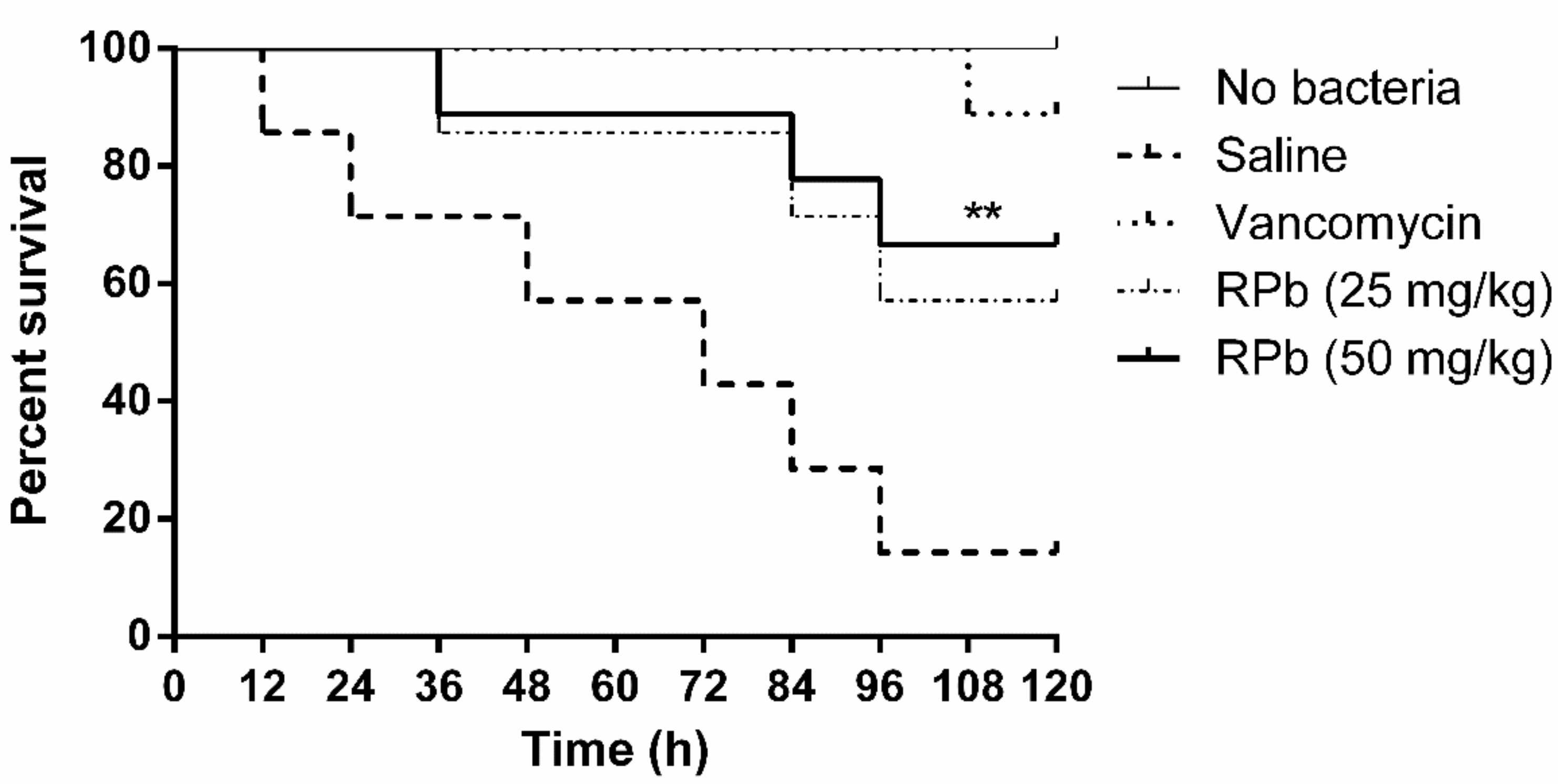
3.10. Treatment of S. Aureus-Infected Waxworms with Peptides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A review on antibiotic resistance: Alarm bells are ringing. Cureus 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. Antimicrobial Resistance Global Report on Surveillance: 2014 Summary; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two hits are better than one: Membrane-active and DNA binding-related double-action mechanism of nk-18, a novel antimicrobial peptide derived from mammalian nk-lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.-S.; Fu, C.-I.; Otto, M. Bacterial strategies of resistance to antimicrobial peptides. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria-Neto, S.; de Almeida, K.C.; Macedo, M.L.R.; Franco, O.L. Understanding bacterial resistance to antimicrobial peptides: From the surface to deep inside. Biochim. Et Biophys. Acta (Bba)-Biomembr. 2015, 1848, 3078–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleitas, O.; Agbale, C.M.; Franco, O.L. Bacterial resistance to antimicrobial peptides: An evolving phenomenon. Front Biosci. 2016, 21, 1013–1038. [Google Scholar]
- Goraya, J.; Knoop, F.C.; Conlon, J.M. Ranatuerins: Antimicrobial peptides isolated from the skin of the american bullfrog, rana catesbeiana. Biochem. Biophys. Res. Commun. 1998, 250, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Conlon, J.M.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D. Primary structures of skin antimicrobial peptides indicate a close, but not conspecific, phylogenetic relationship between the leopard frogs lithobates onca and lithobates yavapaiensis (ranidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 313–317. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Coquet, L.; Jouenne, T.; Leprince, J.; Vaudry, H.; Kolodziejek, J.; Nowotny, N.; King, J.D. Characterization of antimicrobial peptides in skin secretions from discrete populations of lithobates chiricahuensis (ranidae) from central and southern arizona. Peptides 2011, 32, 664–669. [Google Scholar] [CrossRef]
- Zhou, X.; Ma, C.; Zhou, M.; Zhang, Y.; Xi, X.; Zhong, R.; Chen, T.; Shaw, C.; Wang, L. Pharmacological effects of two novel bombesin-like peptides from the skin secretions of chinese piebald odorous frog (odorrana schmackeri) and european edible frog (pelophylax kl. Esculentus) on smooth muscle. Molecules 2017, 22, 1798. [Google Scholar] [CrossRef]
- Wu, D.; Gao, Y.; Tan, Y.; Liu, Y.; Wang, L.; Zhou, M.; Xi, X.; Ma, C.; Bininda-Emonds, O.R.; Chen, T. Discovery of distinctin-like-peptide-ph (dlp-ph) from the skin secretion of phyllomedusa hypochondrialis, a prototype of a novel family of antimicrobial peptide. Front. Microbiol. 2018, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.-H.; Goto, Y.; Réfrégiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-ph: A novel peptide with antimicrobial and anticancer activities from the skin secretion of the south american orange-legged leaf frog, pithecopus (phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm activity and synergistic inhibition of staphylococcus aureus biofilms by bactericidal protein p128 in combination with antibiotics. Antimicrob Agents Chemother 2016, 60, 7280–7289. [Google Scholar]
- Chen, Q.; Cheng, P.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Bian, H.; Chen, T. Evaluating the bioactivity of a novel broad-spectrum antimicrobial peptide brevinin-1gha from the frog skin secretion of hylarana guentheri and its analogues. Toxins 2018, 10, 413. [Google Scholar] [CrossRef] [PubMed]
- Pag, U.; Oedenkoven, M.; Papo, N.; Oren, Z.; Shai, Y.; Sahl, H.-G. In vitro activity and mode of action of diastereomeric antimicrobial peptides against bacterial clinical isolates. J. Antimicrob. Chemother. 2004, 53, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desbois, A.P.; Coote, P.J. Wax moth larva (galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother. 2011, 66, 1785–1790. [Google Scholar] [CrossRef]
- Chen, T.; Zhou, M.; Rao, P.; Walker, B.; Shaw, C. The chinese bamboo leaf odorous frog (rana (odorrana) versabilis) and north american rana frogs share the same families of skin antimicrobial peptides. Peptides 2006, 27, 1738–1744. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from ranid frogs: Taxonomic and phylogenetic markers and a potential source of new therapeutic agents. Biochim. Et Biophys. Acta (Bba)-Proteins Proteom. 2004, 1696, 1–14. [Google Scholar] [CrossRef]
- Tennessen, J.A.; Blouin, M.S. Selection for antimicrobial peptide diversity in frogs leads to gene duplication and low allelic variation. J. Mol. Evol. 2007, 65, 605–615. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, N.; Wang, Z.; Ma, Z.; Zhang, L.; Ma, Q.; Shan, A. Design of imperfectly amphipathic α-helical antimicrobial peptides with enhanced cell selectivity. Acta Biomater. 2014, 10, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Wiradharma, N.; Yang, Y.Y. Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv. Drug Deliv. Rev. 2014, 78, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.-C.; Strandberg, E.; Grau-Campistany, A.; Wadhwani, P.; Reichert, J.; Bürck, J.; Rabanal, F.; Auger, M.l.; Paquin, J.-F.o.; Ulrich, A.S. Influence of the length and charge on the activity of α-helical amphipathic antimicrobial peptides. Biochemistry 2017, 56, 1680–1695. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238. [Google Scholar] [CrossRef] [PubMed]
- Parachin, N.S.; Franco, O.L. New edge of antibiotic development: Antimicrobial peptides and corresponding resistance. Front. Microbiol. 2014, 5, 147. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, Y.K.; Park, J.W.; Lee, B.; Lee, B.J. Solution structure of the antimicrobial peptide gaegurin 4 by 1h and 15n nuclear magnetic resonance spectroscopy. Eur. J. Biochem. 2000, 267, 2695–2704. [Google Scholar] [CrossRef]
- Won, H.-S.; Kang, S.-J.; Lee, B.-J. Action mechanism and structural requirements of the antimicrobial peptides, gaegurins. Biochim. Et Biophys. Acta (Bba)-Biomembr. 2009, 1788, 1620–1629. [Google Scholar] [CrossRef] [Green Version]
- Abraham, P.; Sundaram, A.; Asha, R.; Reshmy, V.; George, S.; Kumar, K.S. Structure-activity relationship and mode of action of a frog secreted antibacterial peptide b1ctcu5 using synthetically and modularly modified or deleted (smmd) peptides. PLoS ONE 2015, 10, e0124210. [Google Scholar] [CrossRef]
- Bao, K.; Yuan, W.; Ma, C.; Wang, L.; Hong, M.; Xi, X.; Zhou, M.; Chen, T. Modification targeting the “rana box” motif of a novel nigrocin peptide from hylarana latouchii enhances and broadens its potency against multiple bacteria. Front. Microbiol. 2018, 9, 2846. [Google Scholar] [CrossRef]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Et Biophys. Acta (Bba)-Biomembr. 2006, 1758, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, R.; Bhunia, A.; Bhattacharjya, S. Micelle-bound structures and dynamics of the hinge deleted analog of melittin and its diastereomer: Implications in cell selective lysis by d-amino acid containing antimicrobial peptides. Biochim. Et Biophys. Acta (Bba)-Biomembr. 2010, 1798, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Maturana, P.; Martinez, M.; Noguera, M.E.; Santos, N.; Disalvo, E.A.; Semorile, L.; Maffia, P.C.; Hollmann, A. Lipid selectivity in novel antimicrobial peptides: Implication on antimicrobial and hemolytic activity. Colloids Surf. B Biointerfaces 2017, 153, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane active antimicrobial peptides: Translating mechanistic insights to design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A. Controlling Biofilm and Persister Cells by Targeting Cell Membranes. Ph.D. Thesis, Syracuse University, Syracuse, NY, USA, 2015. [Google Scholar]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635. [Google Scholar] [CrossRef]
- Sok, M.; Šentjurc, M.; Schara, M.; Stare, J.; Rott, T. Cell membrane fluidity and prognosis of lung cancer. Ann. Thorac. Surg. 2002, 73, 1567–1571. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Antimicrobial peptides (amps) with dual mechanisms: Membrane disruption and apoptosis. J. Microbiol. Biotechnol. 2015, 25, 759–764. [Google Scholar] [CrossRef]
- Vyas, S.; Zaganjor, E.; Haigis, M.C. Mitochondria and cancer. Cell 2016, 166, 555–566. [Google Scholar] [CrossRef]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zai, Y.; Xi, X.; Ma, C.; Wang, L.; Zhou, M.; Shaw, C.; Chen, T. A novel membrane-disruptive antimicrobial peptide from frog skin secretion against cystic fibrosis isolates and evaluation of anti-mrsa effect using galleria mellonella model. Biochim. Et Biophys. Acta (Bba)-Gen. Subj. 2019, 1863, 849–856. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | H | µH | Net Charge (z) |
|---|---|---|---|---|
| Ranatuerin-2Pb | SFLTTVKKLVTNLAALAGTVIDTIKCKVTGGCRT-OH | 0.486 | 0.368 | 4 |
| RPa | SFLTTVKKLVTNLAALAGTVIDTIKCKVTGGC-OH | 0.540 | 0.358 | 3 |
| RPb | SFLTTVKKLVTNLAAL-NH2 | 0.613 | 0.511 | 3 |
| Solution | Peptide | Helix | Antiparallel | Parallel | Turn | Others |
|---|---|---|---|---|---|---|
| 50% TFE/ H2O | Ranatuerin-2Pb | 50 | 7.8 | 1.2 | 10.9 | 29.8 |
| RPa | 33.6 | 18.1 | 4.3 | 11.4 | 32.5 | |
| RPb | 49.6 | 7.6 | 0 | 9.8 | 33 | |
| H2O | Ranatuerin-2Pb | 3.5 | 26.6 | 0 | 18.3 | 51.5 |
| RPa | 2.4 | 30.6 | 0 | 17.3 | 49.7 | |
| RPb | 0.3 | 25.4 | 0 | 19.4 | 55 | |
| POPC/POPG 1:1 | Ranatuerin-2Pb | 59.2 | 1.2 | 2.7 | 10.8 | 26.1 |
| RPa | 24.9 | 13.7 | 8.6 | 13.7 | 39.1 | |
| RPb | 51.8 | 3.6 | 0 | 12.2 | 32.4 | |
| POPE/POPG 3:1 | Ranatuerin-2Pb | 40 | 3.8 | 8.7 | 11.8 | 35.7 |
| RPa | 23.6 | 24.4 | 6.9 | 12.1 | 33 | |
| RPb | 51.6 | 9.2 | 0 | 12 | 27.2 |
| MICs/MBCs (µM) | ||||
|---|---|---|---|---|
| Ampicillin | Ranatuerin-2Pb | RPa | RPb | |
| Staphylococcus aureus | 0.3/0.3 | 8/8 | 16/32 | 8/8 |
| Escherichia coli | 36.6/36.6 | 8/8 | 32/64 | 16/16 |
| Candida albicans | 146/>512 | 8/16 | >256/>256 | 16/16 |
| MRSA | >512/>512 | 16/32 | >256/>256 | 16/32 |
| Enterococcus faecalis | 12.8/12.8 | >256/>256 | >256/>256 | 32/128 |
| Pseudomonas aeruginosa | >512/>512 | >256/>256 | >256/>256 | 64/256 |
| HC50 | >512 | 16.11 | 63.90 | 178 |
| TI (Overall) | 19.42 | 0.449 | 0.353 | 8.83 |
| TI (Gram-positive bacteria and yeast) | 37.20 | 0.503 | 1.258 | 11.125 |
| MBIC/MBEC (µM) | |||
|---|---|---|---|
| Ranatuerin-2Pb | RPa | RPb | |
| S. aureus | 8/32 | 16/128 | 8/32 |
| E. coli | 16/64 | 32/128 | 32/128 |
| C. albicans | 8/32 | >256/>256 | 16/32 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Shi, D.; Zhong, R.; Ye, Z.; Ma, C.; Zhou, M.; Xi, X.; Wang, L.; Chen, T.; Kwok, H.F. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and Its Truncated Analogues. Biomolecules 2019, 9, 249. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060249
Zhou X, Shi D, Zhong R, Ye Z, Ma C, Zhou M, Xi X, Wang L, Chen T, Kwok HF. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and Its Truncated Analogues. Biomolecules. 2019; 9(6):249. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060249
Chicago/Turabian StyleZhou, Xiaowei, Daning Shi, Ruimin Zhong, Zhuming Ye, Chengbang Ma, Mei Zhou, Xinping Xi, Lei Wang, Tianbao Chen, and Hang Fai Kwok. 2019. "Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and Its Truncated Analogues" Biomolecules 9, no. 6: 249. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060249