Identification of MicroRNA Targeting Mlph and Affecting Melanosome Transport


Abstract
:1. Introduction
2. Results
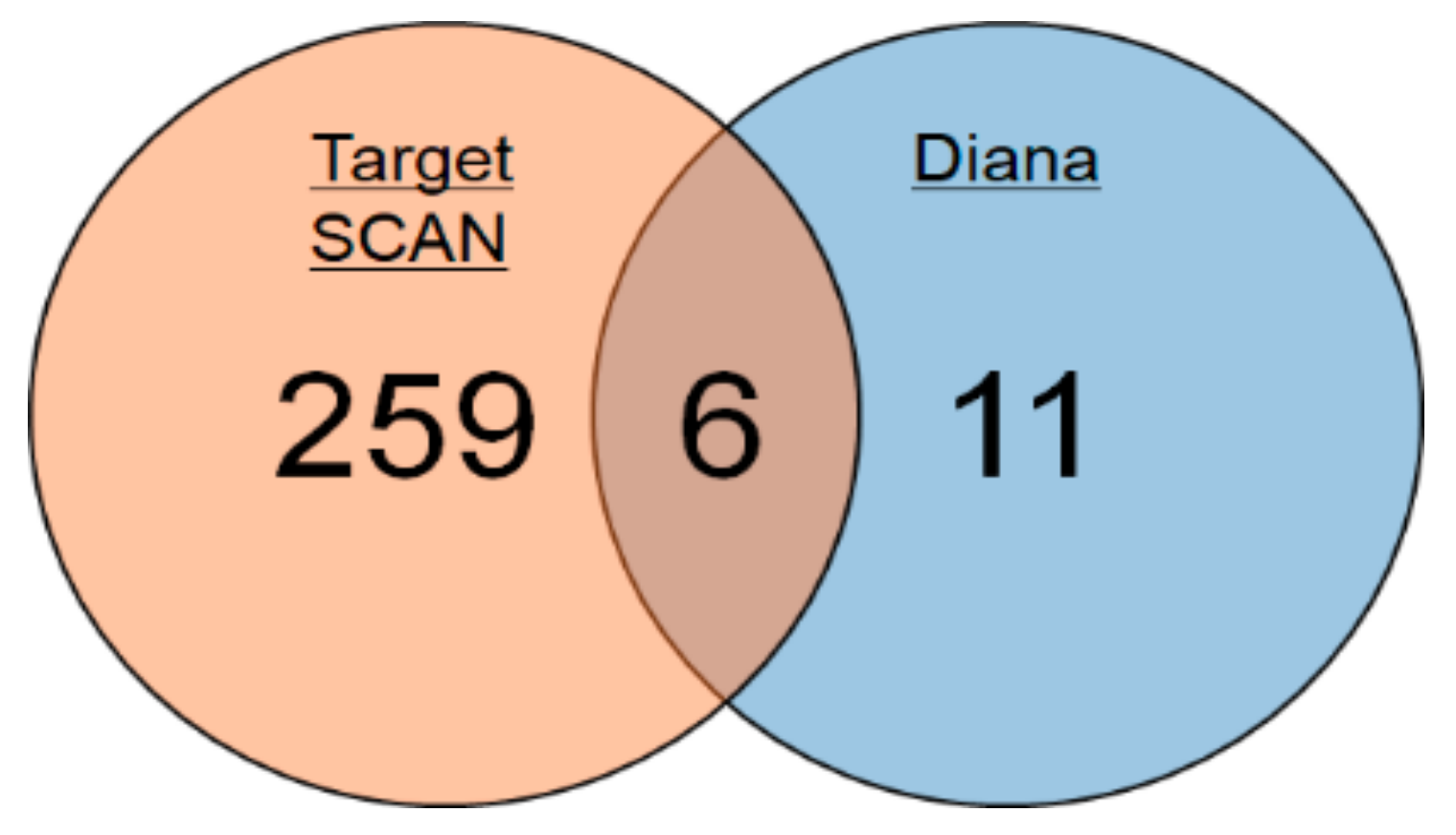
2.1. Identification of miRNAs Targeting Mlph in Mice
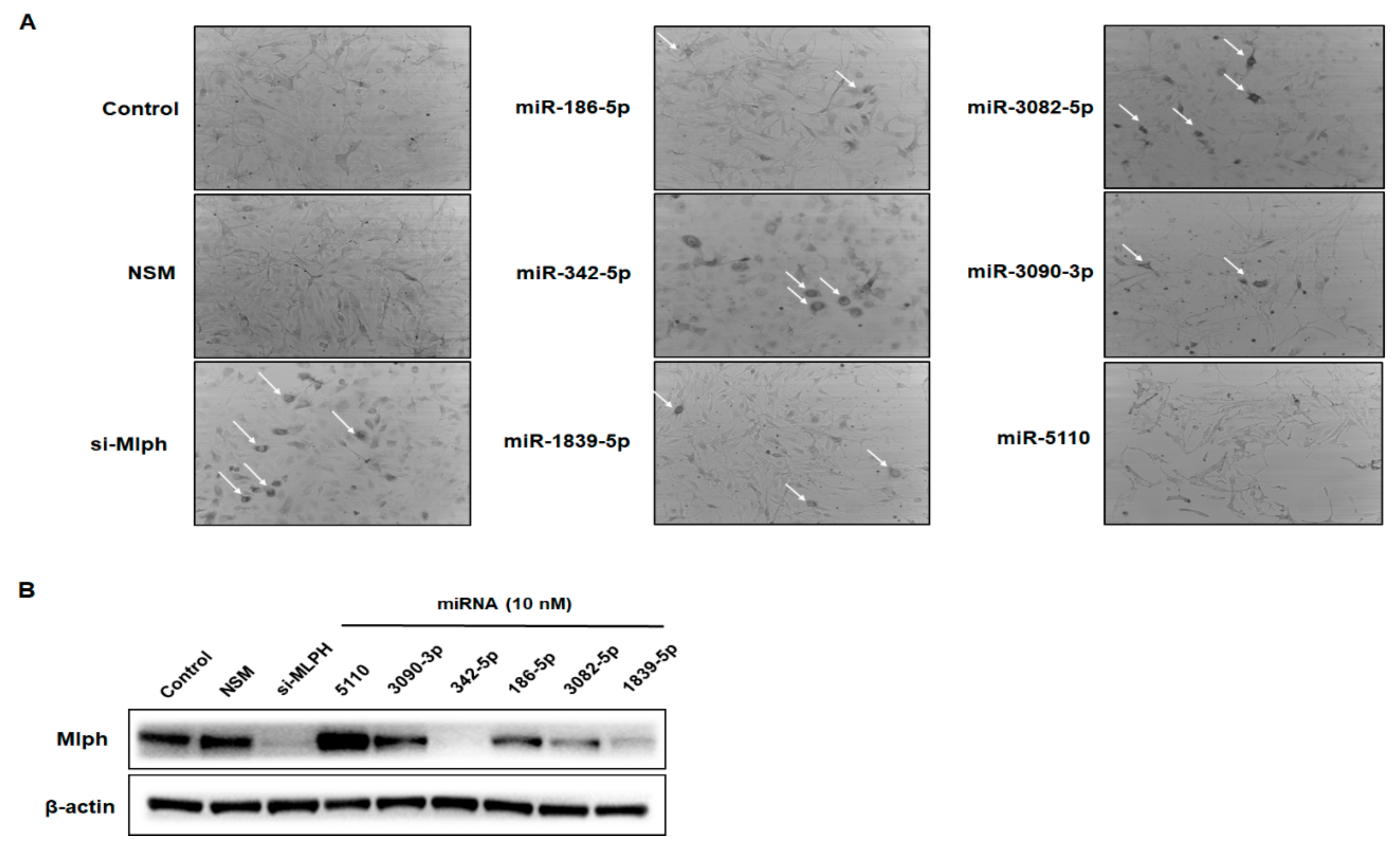
2.2. Effect of miRNAs on Mlph Expression in Melan-a Melanocytes
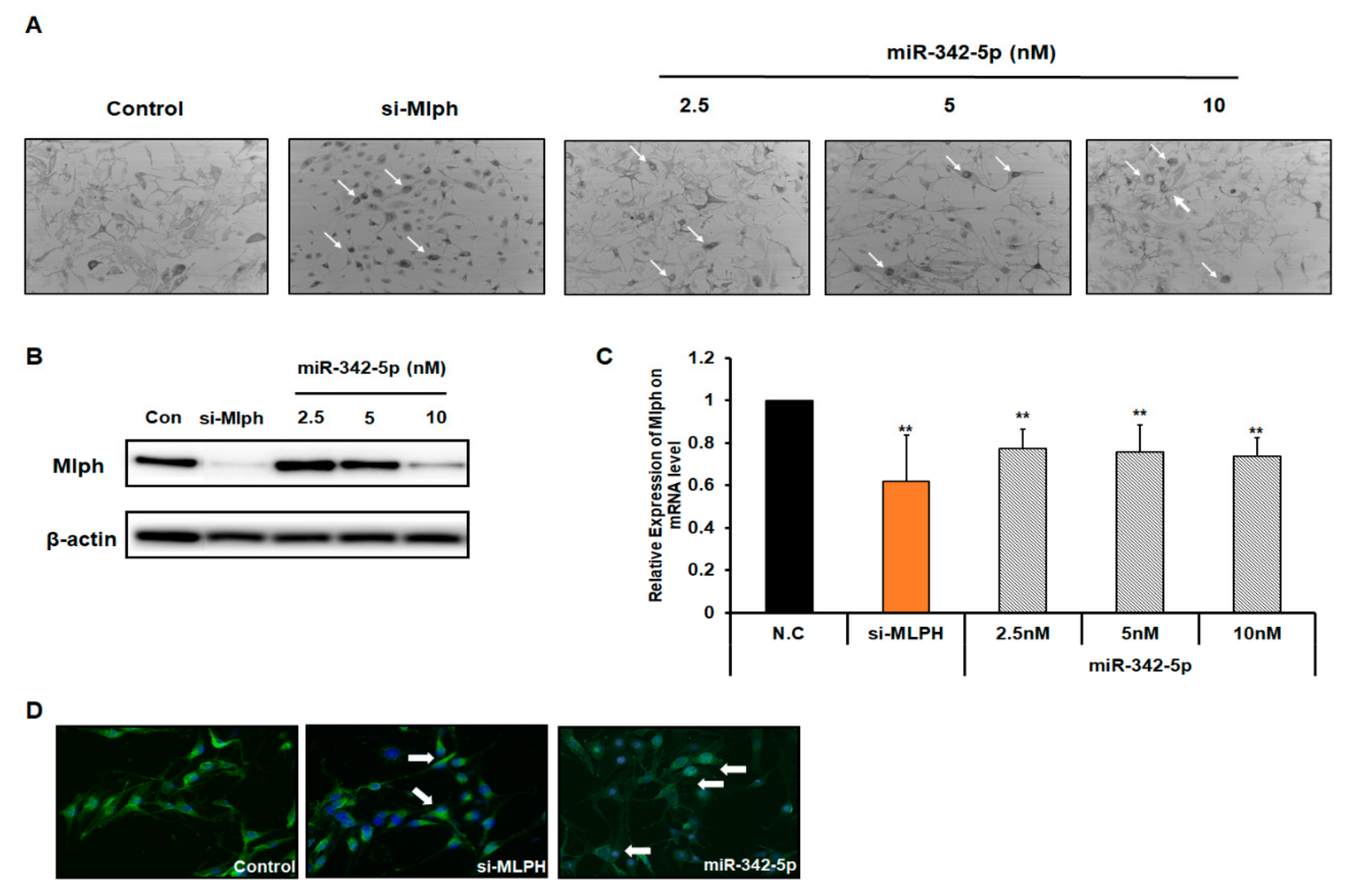
2.3. Effect of miR-342-5p on Mlph Expression in Melan-a Melanocytes
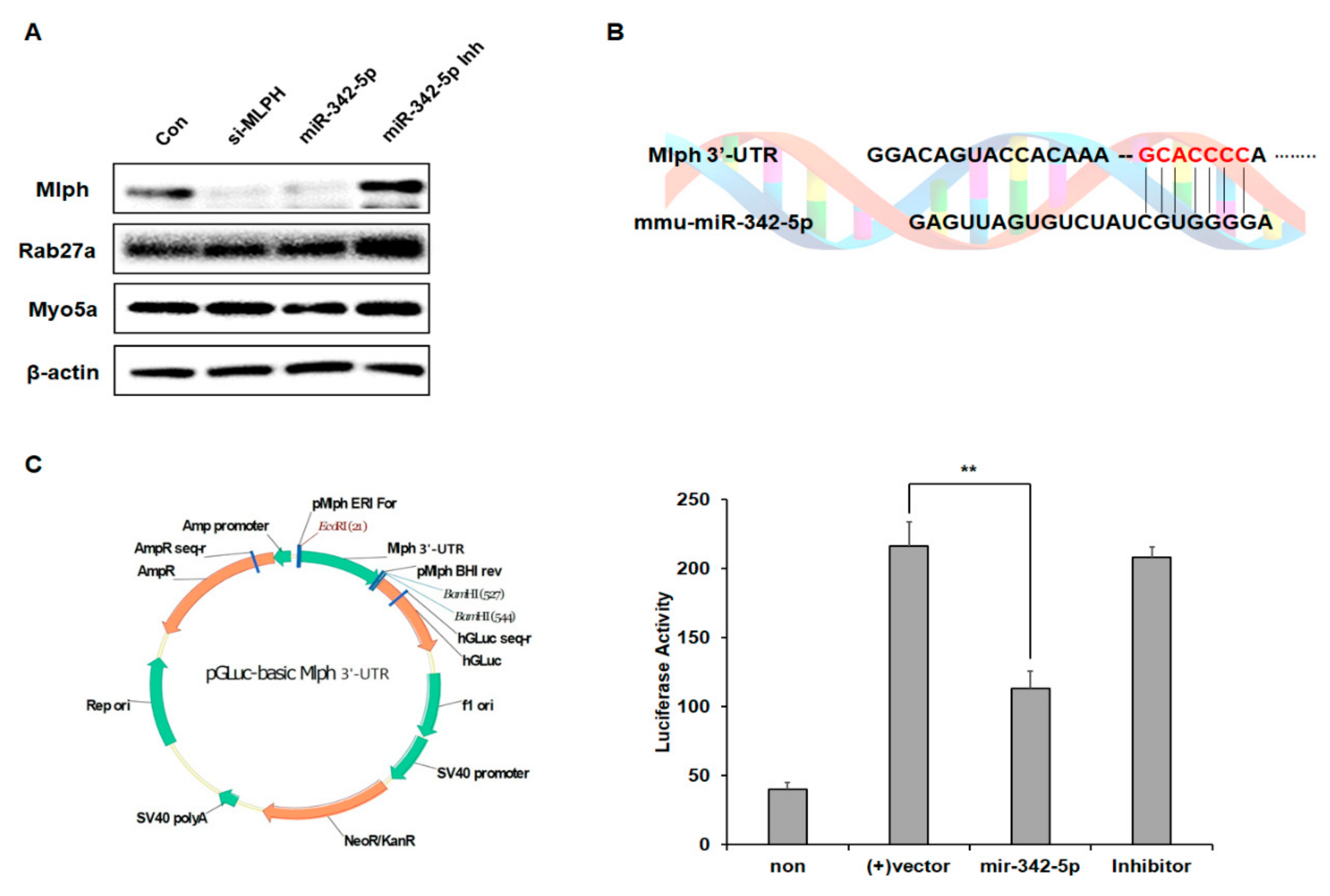
2.4. Effect of mir-342-5p on the Rab27a and MyoVa Expressions
2.5. miR-342-5p Directly Targets Mlph in Melan-a Melanocytes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Transfection
4.4. Western Blotting
4.5. Immuno-Fluorescence (IF)
4.6. Quantitative Real-Time PCR
4.7. miRNA and Small Interfering RNAs
4.8. Detection and Quantification of Melanosome Aggregation
4.9. Luciferase Assay
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gillbro, J.M.; Olsson, M.J. The melanogenesis and mechanisms of skin-lightening agents—Existing and new approaches. Int. J. Cosmet. Sci. 2011, 33, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Thody, A.J.; Higgins, E.M.; Wakamatsu, K.; Ito, S.; Burchill, S.A.; Marks, J.M. Pheomelanin as well as eumelanin is present in human epidermis. J. Invest. Dermatol. 1991, 97, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tyminska, A. Skin melanocytes: Biology and development. Postepy Dermatol. I. Alergol. 2013, 30, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, M.V. Signaling pathways in melanosome biogenesis and pathology. Int. J. Biochem. Cell Biol. 2010, 42, 1094–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.I.; Lee, H.Y.; Lee, J.E.; Myung, C.H.; Hwang, J.S. Inhibitory effect of 2-methyl-naphtho[1,2,3-de] quinolin-8-one on melanosome transport and skin pigmentation. Sci. Rep. 2016, 6, 29189. [Google Scholar] [CrossRef] [PubMed]
- Chia, W.J.; Tang, B.L. Emerging roles for Rab family GTPases in human cancer. Biochim. Biophys. Acta 2009, 1795, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Matesic, L.E.; Yip, R.; Reuss, A.E.; Swing, D.A.; O’Sullivan, T.N.; Fletcher, C.F.; Copeland, N.G.; Jenkins, N.A. Mutations in Mlph, encoding a member of the Rab effector family, cause the melanosome transport defects observed in leaden mice. Proc. Natl. Acad. Sci. USA 2001, 98, 10238–10243. [Google Scholar] [CrossRef]
- Griscelli, C.; Durandy, A.; Guy-Grand, D.; Daguillard, F.; Herzog, C.; Prunieras, M. A syndrome associating partial albinism and immunodeficiency. Am. J. Med. 1978, 65, 691–702. [Google Scholar] [CrossRef]
- Malhotra, A.K.; Bhaskar, G.; Nanda, M.; Kabra, M.; Singh, M.K.; Ramam, M. Griscelli syndrome. J. Am. Acad. Dermatol. 2006, 55, 337–340. [Google Scholar] [CrossRef]
- Aslan, D.; Sari, S.; Derinoz, O.; Dalgic, B. Griscelli syndrome: description of a case with Rab27A mutation. Pediatr. Hematol. Oncol. 2006, 23, 255–261. [Google Scholar] [CrossRef]
- Menasche, G.; Ho, C.H.; Sanal, O.; Feldmann, J.; Tezcan, I.; Ersoy, F.; Houdusse, A.; Fischer, A.; de Saint Basile, G. Griscelli syndrome restricted to hypopigmentation results from a melanophilin defect (GS3) or a MYO5A F-exon deletion (GS1). J. Clin. Invest. 2003, 112, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Westbroek, W.; Klar, A.; Cullinane, A.R.; Ziegler, S.G.; Hurvitz, H.; Ganem, A.; Wilson, K.; Dorward, H.; Huizing, M.; Tamimi, H.; et al. Cellular and clinical report of new Griscelli syndrome type III cases. Pigment Cell Melanoma Res. 2012, 25, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, X.I.; Wang, A.; Meisgen, F.; Pivarcsi, A.; Sonkoly, E.; Stahle, M.; Landen, N.X. MicroRNA-31 Promotes Skin Wound Healing by Enhancing Keratinocyte Proliferation and Migration. J. Invest. Derm. 2015, 135, 1676–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Zhang, J.F.; Wang, W.M.; Cheung, F.W.; Lu, Y.F.; Ng, C.F.; Kung, H.F.; Liu, W.K. MicroRNA-218 inhibits melanogenesis by directly suppressing microphthalmia-associated transcription factor expression. Rna Biol. 2014, 11, 732–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, S.; Kumazaki, M.; Mori, T.; Baba, K.; Okuda, M.; Mizuno, T.; Akao, Y. Analysis of microRNA-203 function in CREB/MITF/RAB27a pathway: Comparison between canine and human melanoma cells. Vet. Comp. Oncol. 2016, 14, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Dynoodt, P.; Speeckaert, R.; De Wever, O.; Chevolet, I.; Brochez, L.; Lambert, J.; Van Gele, M. miR-145 overexpression suppresses the migration and invasion of metastatic melanoma cells. Int. J. Oncol. 2013, 42, 1443–1451. [Google Scholar] [CrossRef] [Green Version]
- Wasmeier, C.; Hume, A.N.; Bolasco, G.; Seabra, M.C. Melanosomes at a glance. J. Cell Sci. 2008, 121, 3995–3999. [Google Scholar] [CrossRef] [Green Version]
- Mackintosh, J.A. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001, 211, 101–113. [Google Scholar] [CrossRef]
- Van Gele, M.; Geusens, B.; Schmitt, A.M.; Aguilar, L.; Lambert, J. Knockdown of myosin Va isoforms by RNAi as a tool to block melanosome transport in primary human melanocytes. J. Invest. Derm. 2008, 128, 2474–2484. [Google Scholar] [CrossRef]
- Van Den Bossche, K.; Naeyaert, J.M.; Lambert, J. The quest for the mechanism of melanin transfer. Traffic 2006, 7, 769–778. [Google Scholar] [CrossRef]
- Yang, S.; Liu, B.; Ji, K.; Fan, R.; Dong, C. MicroRNA-5110 regulates pigmentation by cotargeting melanophilin and WNT family member 1. FASEB J. 2018, 32, 5405–5412. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Akiyama, Y.; Yuasa, Y. Multiple-to-multiple relationships between microRNAs and target genes in gastric cancer. PLoS ONE 2013, 8, e62589. [Google Scholar] [CrossRef] [PubMed]
- Pham, X.; Song, G.; Lao, S.; Goff, L.; Zhu, H.; Valle, D.; Avramopoulos, D. The DPYSL2 gene connects mTOR and schizophrenia. Transl. Psychliatry 2016, 6, e933. [Google Scholar] [CrossRef] [PubMed]
- Dahmke, I.N.; Backes, C.; Rudzitis-Auth, J.; Laschke, M.W.; Eidinger, P.; Menger, M.D.; Meese, E.; Mahlknecht, U. Curcumin intake affects miRNA signature in murine melanoma with mmu-miR-205-5p most significantly altered. PLoS ONE 2013, 8, e81122. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wu, Y.; Gu, M.; Zhang, Y. miR-342-5p decreases ankyrin G levels in Alzheimer’s disease transgenic mouse models. Cell Rep. 2014, 6, 264–270. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| microRNA | Sequence |
|---|---|
| miR-1839-5p | 5’-AAGGUAGAUAGAACAGGUCUUG-3’ |
| miR-5110 | 5’-GGAGGAGGUAGAGGGUGGUGGAAUU-3’ |
| miR-3082-5p | 5’-GACAGAGUGUGUGUGUCUGUGU-3’ |
| miR-342-5p | 5’-AGGGGUGCUAUCUGUGAUUGAG-3’ |
| miR-3090-3p | 5’-UCCCAGGUGACACCCUGACUCA-3’ |
| miR-186-5p | 5’-CAAAGAAUUCUCCUUUUGGGCU-3’ |
| miR-1839-5p Inh. | 5’-CAAGACCUGUUCUAUCUACCUU-3’ |
| miR-5110 Inh. | 5’-AAUUCCACCACCCUCUACCUCCUCC-3’ |
| miR-3082-5p Inh. | 5’-ACACAGACACACACACUCUGUC-3’ |
| miR-342-5p Inh. | 5’-CUCAAUCACAGAUAGCACCCCU-3’ |
| miR-3090-3p Inh. | 5’-UGAGUCAGGGUGUCACCUGGGA-3’ |
| miR-186-5p Inh. | 5’-AGCCCAAAAGGAGAAUUCUUUG-3’ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.A.; Hwang, S.J.; Hong, S.C.; Myung, C.H.; Lee, J.E.; Park, J.I.; Hwang, J.S. Identification of MicroRNA Targeting Mlph and Affecting Melanosome Transport. Biomolecules 2019, 9, 265. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9070265
Lee JA, Hwang SJ, Hong SC, Myung CH, Lee JE, Park JI, Hwang JS. Identification of MicroRNA Targeting Mlph and Affecting Melanosome Transport. Biomolecules. 2019; 9(7):265. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9070265
Chicago/Turabian StyleLee, Jeong Ah, Seok Joon Hwang, Sung Chan Hong, Cheol Hwan Myung, Ji Eun Lee, Jong Il Park, and Jae Sung Hwang. 2019. "Identification of MicroRNA Targeting Mlph and Affecting Melanosome Transport" Biomolecules 9, no. 7: 265. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9070265