
Serial Section-Based Three-Dimensional Reconstruction of Anaxagorea (Annonaceae) Carpel Vasculature and Implications for the Morphological Relationship between the Carpel and the Ovule


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
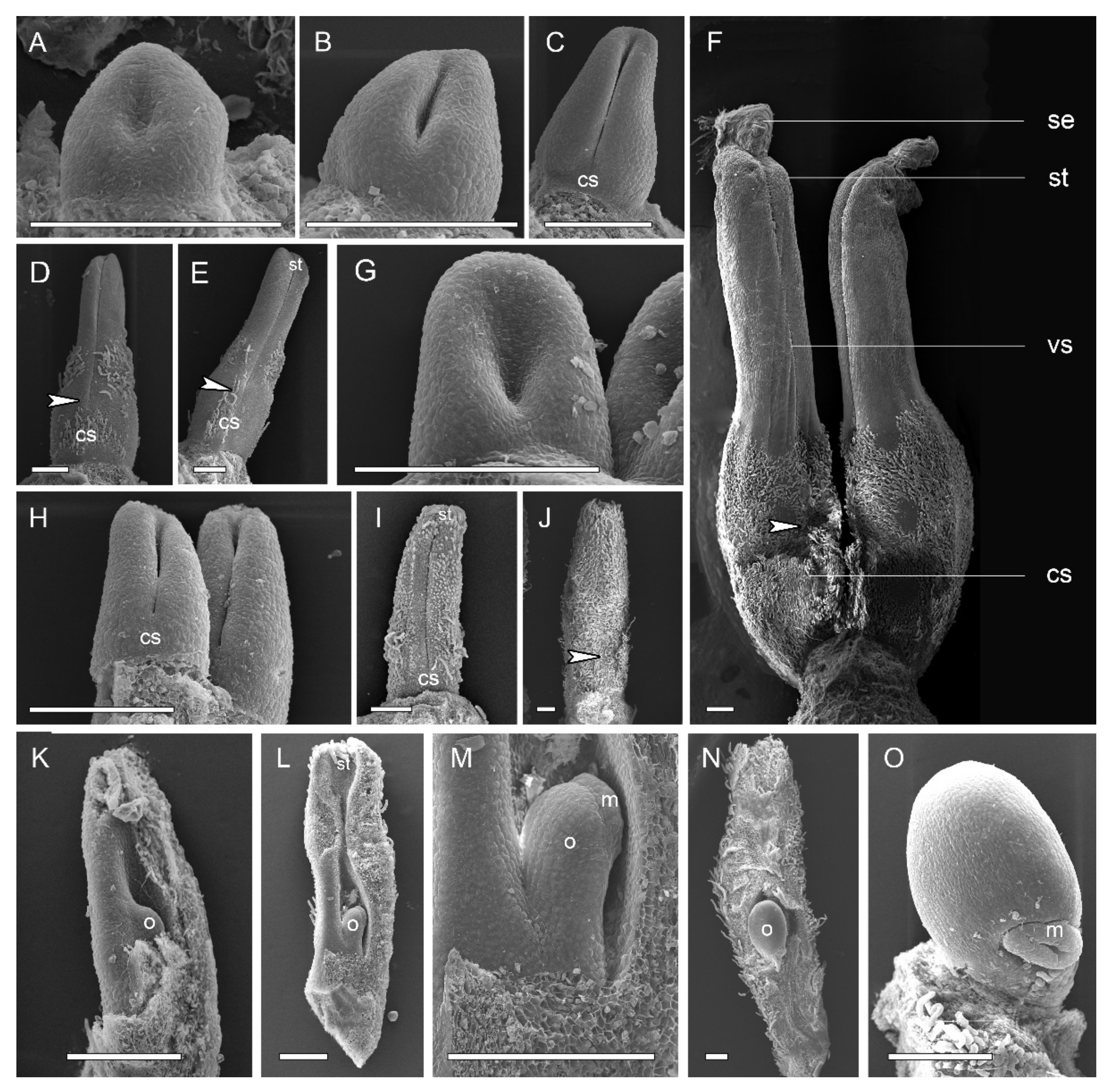
2.1. Gynoecium Structure and Carpel Organogenesis
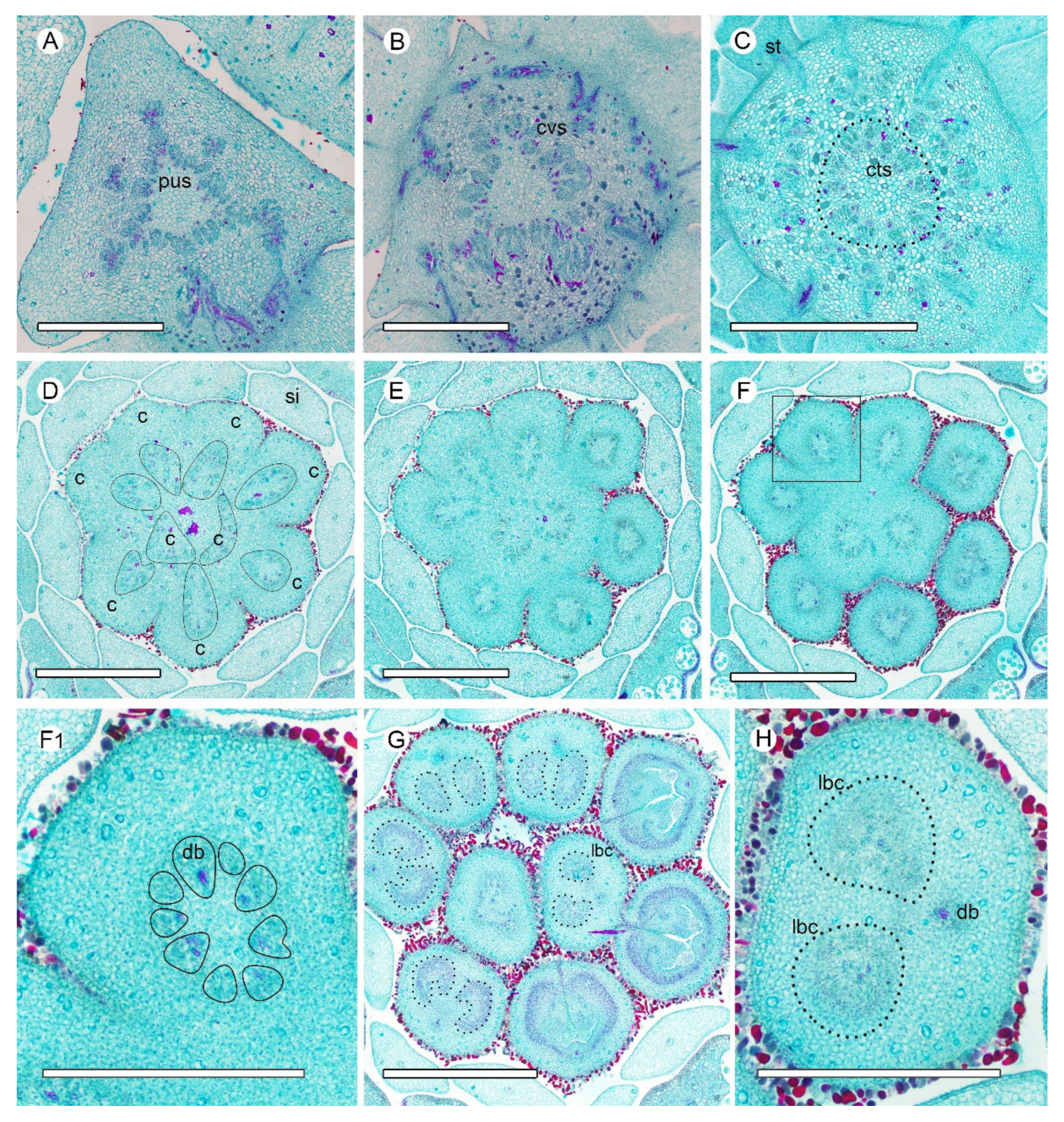
2.2. Vasculature from Receptacle to Carpel
2.3. D-Reconstruction of Carpel Vasculature
3. Discussion
3.1. Carpel Organogenesis
3.2. Carpel Vasculature
3.3. Implications for Evolution
4. Materials and Methods
4.1. Scanning Electron Microscopy and Paraffin Sectioning
4.2. Topological Analysis of Carpel Vasculature
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

Appendix C

References
- Kennedy, D.; Norman, C. What don’t we know? Science 2005, 309, 78–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuessy, T.F. A transitional-combinational theory for the origin of angiosperms. Taxon 2004, 53, 3–16. [Google Scholar] [CrossRef]
- Dunal, M.F. Monographie de la Famille des Anonacées; Treuttel & Würtz: Paris, France, 1817; pp. 10–14. [Google Scholar]
- Robinson-Beers, K.; Pruitt, R.E.; Gasser, C.S. Ovule development in wild-type Arabidopsis and two female-sterile mutants. Plant Cell 1992, 4, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Endress, P.K. Patterns of angiospermy development before carpel sealing across living angiosperms: Diversity, and morphological and systematic aspects. Bot. J. Linn. Soc. 2015, 178, 556–591. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.M. Caytoniales, Cycadales & Pteridosperms. In The Yorkshire Jurassic Flora; Harris, T.M., Millington, W., Miller, J., Eds.; British Museum (Natural History): London, UK, 1964; Volume 2, pp. 1–28. [Google Scholar]
- Doyle, J.A. Origin of angiosperms. Annu. Rev. Ecol. Syst. 1978, 9, 365–392. [Google Scholar] [CrossRef]
- Doyle, J.A. Integrating molecular phylogenetic and paleobotanical evidence on origin of the flower. Int. J. Plant Sci. 2008, 169, 816–843. [Google Scholar] [CrossRef] [Green Version]
- Retallack, G.; Dilcher, D.L. A coastal hypothesis for the dispersal and rise to dominance of flowering plants. In Paleobotany, Paleoecology and Evolution; Niklas, K.J., Ed.; Praeger Publishers: New York, NY, USA, 1981; pp. 27–77. [Google Scholar]
- Frohlich, M.W.; Parker, D.S. The mostly male theory of flower evolutionary origins: From genes to fossils. Syst. Bot. 2000, 25, 155–170. [Google Scholar] [CrossRef]
- Wang, X. The Dawn Angiosperms, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 293–366. [Google Scholar]
- Shi, G.; Herrera, F.; Herendeen, P.S.; Clark, E.G.; Crane, P.R. Mesozoic cupules and the origin of the angiosperm second integument. Nature 2021, 594, 223–226. [Google Scholar] [CrossRef]
- Angenent, G.C.; Franken, J.; Busscher, M.; van Dijken, A.; van Went, J.L.; Dons, H.J.; van Tunen, A.J. A novel class of MADS box genes is involved in ovule development in Petunia. Plant Cell 1995, 7, 1569–1582. [Google Scholar] [CrossRef]
- Roe, J.L.; Nemhauser, J.L.; Zambrysk, P.C. TOUSLED Participates in apical tissue formation during gynoecium development in Arabidopsis. Plant Cell 1997, 9, 335–353. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.A. Seed ferns and the origin of angiosperms. J. Torrey Bot. Soc. 2006, 133, 169–209. [Google Scholar] [CrossRef] [Green Version]
- Frohlich, M.W.; Chase, M.W. After a dozen years of progress the origin of angiosperms is still a great mystery. Nature 2007, 450, 1184–1189. [Google Scholar] [CrossRef]
- Wynn, A.N.; Seaman, A.A.; Jones, A.L.; Franks, R.G. Novel functional roles for PERIANTHIA and SEUSS during floral organ identity specification, floral meristem termination, and gynoecial development. Front. Plant Sci. 2014, 5, 130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, W.; Wang, X. How the ovules get enclosed in magnoliaceous carpels. PLoS ONE 2017, 12, e0174955. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Z.X.; Zhao, Z. Floral ontogeny of Illicium lanceolatum (Schisandraceae) and its implications on carpel homology. Phytotaxa 2019, 416, 200–210. [Google Scholar] [CrossRef]
- Guo, X.M.; Xiao, X.; Wang, G.X.; Gao, R.F. Vascular anatomy of kiwi fruit and its implications for the origin of carpels. Front. Plant Sci. 2013, 4, 391. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.Z.; Hilu, K.; Wang, Y.L. From leaf and branch into a flower: Magnolia tells the story. Bot. Stud. 2014, 55, 28. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.M.; Yu, Y.Y.; Bai, L.; Gao, R.F. Dianthus chinensis L.: The structural difference between vascular bundles in the placenta and ovary wall suggests their different origin. Front. Plant Sci. 2017, 8, 1986. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.A.; Le Thomas, A. Phylogenetic analysis and character evolution in Annonaceae. Adansonia 1996, 18, 279–334. [Google Scholar]
- Sauquet, H.; Doyle, J.A.; Scharaschkin, T.; Borsch, T.; Hilu, K.W.; Chatrou, L.W.; Le Thomas, A. Phylogenetic analysis of Magnoliales and Myristicaceae based on multiple data sets: Implications for character evolution. Bot. J. Linn. Soc. 2003, 142, 125–186. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.A.; Sauquet, H.; Scharaschkin, T.; Le Thomas, A. Phylogeny, molecular and fossil dating, and biogeographic history of Annonaceae and Myristicaceae (Magnoliales). Int. J. Plant Sci. 2004, 165, S55–S67. [Google Scholar] [CrossRef] [Green Version]
- Chatrou, L.W.; Pirie, M.D.; Erkens, R.H.J.; Couvreur, T.L.P.; Neubig, K.M.; Abbott, J.R.; Mols, J.B.; Maas, J.W.; Saunders, R.M.K.; Chase, M.W. A new subfamilial and tribal classification of the pantropical flowering plant family Annonaceae informed by molecular phylogenetics. Bot. J. Linn. Soc. 2012, 169, 5–40. [Google Scholar] [CrossRef] [Green Version]
- Chatrou, L.W.; Turner, I.M.; Klitgaard, B.B.; Maas, P.J.M.; Utteridge, T.M.A. A linear sequence to facilitate curation of herbarium specimens of Annonaceae. Kew Bull. 2018, 73, 39. [Google Scholar] [CrossRef]
- Deroin, T. Aspects Anatomiques et Biologiques de la Fleur des Annonacées. Ph.D. Thesis, Université de Paris-Sud, Centre d’Orsay, France, 1988. [Google Scholar]
- Endress, P.K.; Armstrong, J.E. Floral development and floral phyllotaxis in Anaxagorea (Annonaceae). Ann. Bot. 2011, 108, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Deroin, T. Définition et Signification Phylogénique des Systèmes Corticaux Floraux: L’Exemple des Annonacées; Série III; Comptes Rendus de l’Académie des Sciences: Paris, France, 1989; Volume 308, pp. 71–75. [Google Scholar]
- De Craene, L.P.R.; Smets, E.F. The distribution and systematic relevance of the androecial character polymery. Bot. J. Linn. Soc. 1993, 113, 285–350. [Google Scholar] [CrossRef]
- Deroin, T.; Norman, É.M. Notes on the floral anatomy of Deeringothamnus Small (Annonaceae): Cortical vascular systems in a chaotic pattern. Mod. Phytol. 2016, 9, 3–12. [Google Scholar]
- Deroin, T.; Bidault, E. Floral anatomy of Pseudartabotrys Pellegrin (Annonaceae), a monospecific genus endemic to Gabon. Adansonia 2017, 39, 111–123. [Google Scholar] [CrossRef]
- Deroin, T. Confirmation and origin of the paracarpy in Annonaceae, with comments on some methodological aspects. Candollea 1997, 52, 45–58. [Google Scholar]
- Deroin, T.; Le Thomas, A. Sur la systématique et les potentialités évolutives des Annonacées: Cas d’Ambavia gerrardii In Espèce Endémique de Madagascar; Le Thomas, A., Ed.; Série III; Comptes Rendus de l’Académie des Sciences: Paris, France, 1989; pp. 647–652. [Google Scholar]
- Deroin, T. Anatomie florale de Meiocarpidium Engler & Diels (Annonaceae-Unoneae). Bulletin du Muséum national d’histoire naturelle, Section B. Adansonia 1987, 9, 81–93. [Google Scholar]
- Endress, P.K. The morphological relationship between carpels and ovules in angiosperms: Pitfalls of morphological interpretation. Bot. J. Linn. Soc. 2019, 189, 201–227. [Google Scholar] [CrossRef] [Green Version]
- Rohweder, O. Anatomische und histogenetische Untersuchungen an Laubsprossen und Blüten der Commelinaceen. Bot. Jahrbücher für Syst. 1963, 82, 1–99. [Google Scholar]
- Chandler, J.W. Founder cell specification. Trends Plant Sci. 2011, 16, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Perales, M.; Reddy, G.V. Stem cell maintenance in shoot apical meristems. Curr. Opin. Plant Biol. 2012, 15, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Ito, M.; Kato, M. Expression pattern of INNER NO OUTER homologue in Nymphaea (water lily family, Nymphaeaceae). Dev. Genes Evol. 2003, 213, 510–513. [Google Scholar] [CrossRef]
- Yamada, T.; Sasaki, Y.; Hashimoto, K.; Nakajima, K.; Gasser, C.S. CORONA, PHABULOSA and PHAVOLUTA collaborate with BELL1 to confine WUSCHEL expression to the nucellus in Arabidopsis ovules. Development 2016, 143, 422–426. [Google Scholar] [CrossRef] [Green Version]
- Esau, K. Vascular Differentiation in Plants; Holt, Rinehart and Winston: New York, NY, USA, 1965. [Google Scholar]
- Evert, R.F.; Eichhorn, S.E. Raven Biology of Plants, 8th ed.; W.H. Freeman Press: New York, NY, USA, 2013; pp. 579–613. [Google Scholar]
- Alvarez, J.; Smyth, D.R. CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development 1999, 126, 2377–2386. [Google Scholar] [CrossRef]
- Theißen, G.; Melzer, R.; Rümpler, F. MADS-domain transcription factors and the floral quartet model of flower development: Linking plant development and evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, N.; Huang, J.; Tatsumi, Y.; Abe, M.; Sugano, S.S.; Kojima, M.; Takebayashi, Y.; Kiba, T.; Yokoyama, R.; Nishitani, K.; et al. Chromatin-mediated feed-forward auxin biosynthesis in floral meristem determinacy. Nat. Commun. 2018, 9, 5290. [Google Scholar] [CrossRef]
- Gambaryan, P.P.; Kuznetsov, A.N. Gnetum and Nymphaeaceans as Models for a Scenario of the Origin of Morphotype of Flowering Plants. Biol. Bull. Rev. 2021, 11, 237–253. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Du, W.; Chen, Y.; Wang, S.; Wang, X.-F. Serial Section-Based Three-Dimensional Reconstruction of Anaxagorea (Annonaceae) Carpel Vasculature and Implications for the Morphological Relationship between the Carpel and the Ovule. Plants 2021, 10, 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10102221
Li Y, Du W, Chen Y, Wang S, Wang X-F. Serial Section-Based Three-Dimensional Reconstruction of Anaxagorea (Annonaceae) Carpel Vasculature and Implications for the Morphological Relationship between the Carpel and the Ovule. Plants. 2021; 10(10):2221. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10102221
Chicago/Turabian StyleLi, Ya, Wei Du, Ye Chen, Shuai Wang, and Xiao-Fan Wang. 2021. "Serial Section-Based Three-Dimensional Reconstruction of Anaxagorea (Annonaceae) Carpel Vasculature and Implications for the Morphological Relationship between the Carpel and the Ovule" Plants 10, no. 10: 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10102221