
Comparative Transcriptomic Analysis Provides Novel Insights into the Blanched Stem of Oenanthe javanica

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Transcriptome Sequencing and Assembly
2.2. Unigene Annotation and Classification
2.3. Differentially Expressed Genes (DEG) in Water Dropwort Plant in Blanching Conditions
2.4. DEGs Functional Annotation
2.5. Gene Ontology (GO) Enrichment Analysis
2.6. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
2.6.1. Photosynthesis Related Pathways
2.6.2. Photosynthesis-Antenna Proteins Pathway
2.6.3. Carbon Fixation in Photosynthetic Organisms
2.6.4. Porphyrin and Chlorophyll Metabolism Pathway
2.6.5. Plant Hormone Signal Transduction Pathway
2.7. Transcription Factors
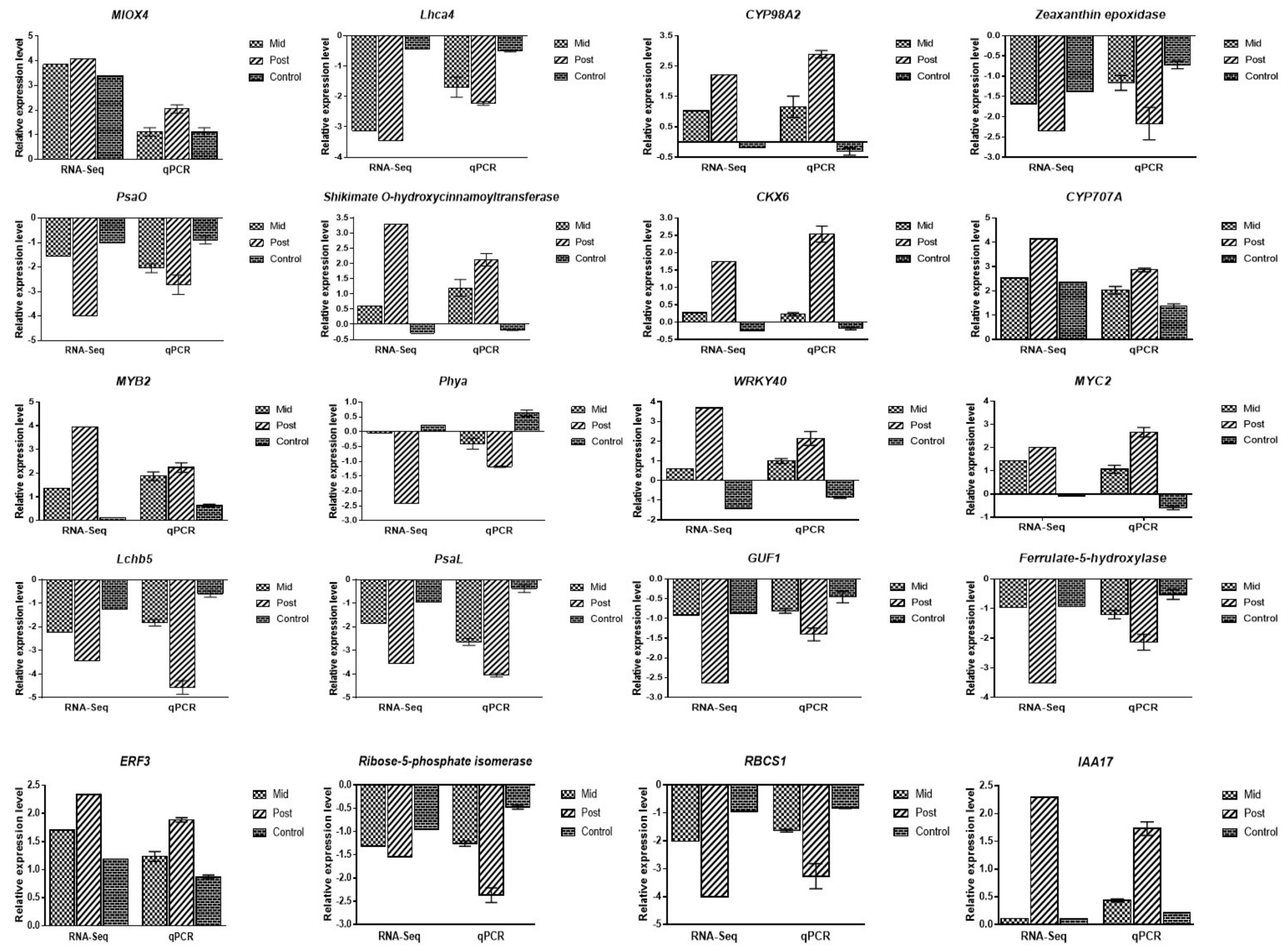
2.8. Validation of DEGs
3. Discussion
3.1. Blanching Affects the Light-Dependent and Light-Independent Reactions of Photosynthesis in Water Dropwort
3.2. Plant Hormone Signal Transduction Pathways
3.3. Transcription Factors
4. Material and Methods
4.1. Experimental Conditions
4.2. Deep Planting Technique Used for Blanch Cultivation
4.3. RNA Extraction, Library Construction, and Illumina Sequencing
4.4. Quality Control and Transcriptome Assembling
4.5. DEG Identification and Functional Annotation
4.6. Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, C.-L.; Li, X.-F. A Review ofOenanthe javanica(Blume) DC. as Traditional Medicinal Plant and Its Therapeutic Potential. Evid.-Based Complement. Altern. Med. 2019, 2019, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Wang, F.; Tan, H.-W.; Li, M.-Y.; Xu, Z.-S.; Tan, G.-F.; Xiong, A.-S. De novo transcriptome assembly, gene annotation, marker development, and miRNA potential target genes validation under abiotic stresses in Oenanthe javanica. Mol. Genet. Genom. 2014, 290, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Zhou, K.; Khan, S.; Ke, W.; Hou, H. Investigation of an Antioxidative System for Salinity Tolerance in Oenanthe javanica. Antioxidants 2020, 9, 940. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Wong, S.K.; Chan, H.T. Ulam Herbs of Oenanthe javanica and Cosmos caudatus: An overview on their Medicinal Properties. J. Nat. Remedies 2017, 16, 137–147. [Google Scholar] [CrossRef]
- Ma, C.J.; Lee, K.Y.; Jeong, E.J.; Kim, S.H.; Park, J.; Choi, Y.H.; Kim, Y.C.; Sung, S.H. Persicarin from water dropwort (Oenanthe javanica) protects primary cultured rat cortical cells from glutamate-induced neurotoxicity. Phytotherapy Res. 2009, 24, 913–918. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Huang, X.; Ji, Q.; Zhou, K.; Hou, H.; Ke, W.; Yang, J. Phenotypic, Nutritional, and Antioxidant Characterization of Blanched Oenanthe javanica for Preferable Cultivar. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Tan, Y.P.; Chin, S.J.; Gan, L.Y.; Kang, K.X.; Fong, C.H.; Chang, H.Q.; How, Y.C. Antioxidant properties of selected fresh and processed herbs and vegetables. Free Rad. Antiox. 2014, 4, 39–46. [Google Scholar] [CrossRef]
- Kong, Q.; Zhao, Y.; Guan, P.; Ke, W.; Lin, C. Chapter 5 Water dropwort. In The Aquatic Vegetable Cultivars and Resources of China; Kong, Q., Ed.; Hubei Science and Technology Press: Wuhan, China, 2005; pp. 175–215. ISBN 7-5352-3339-2. [Google Scholar]
- Yuanying, Y.; Xinfang, H.; Weidong, K. A brief introduction of cultivation technology of water celery. Aquat. Veg. 2008, 11, 34–35. [Google Scholar] [CrossRef]
- Yuanying, Y.; Weidong, K.; Yalin, S.; Minghua, L.; Honglian, Z.; Laichun, H. Cultivar comparative trial of water dropwort (Oenanthe decumbens) under different intenerate methods. J. Chang. Veg. 2009, 16, 45–46. [Google Scholar] [CrossRef]
- Zhu, W.; Wu, Z.; Liang, J.; Yin, X.; Li, W.; Li, M. Cultivation techniques of water celery with mulching and softening. J. Chang. Veg. 2013, 18, 183–184. [Google Scholar] [CrossRef]
- Wang, N.; Qian, Z.; Luo, M.; Fan, S.; Zhang, X.; Zhang, L. Identification of Salt Stress Responding Genes Using Transcriptome Analysis in Green Alga Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2018, 19, 3359. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Chen, C.; Shi, Z.H.; Cheng, H.M.; Bing, J.; Ma, X.F.; Zheng, C.X.; Li, H.J.; Zhang, G.F. Identification of salinity-related genes in ENO2 mutant (eno2−) of Arabidopsis thaliana. J. Integr. Agric. 2018, 17, 94–110. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kanehisa, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, 277D–280D. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2018, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Zhao, J.; Pan, T.; Xi, L.; Zhang, J.; Zou, Z. Comparative Transcriptome Analysis of Gene Expression Patterns in Tomato Under Dynamic Light Conditions. Genes 2019, 10, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Yuan, J.-L.; Li, Y.-X.; Wang, C.-J.; Tang, H.-Y.; Xia, J.-H.; Yang, Q.-Y.; Wan, Z.-J. Transcriptome Analysis Reveals Candidate Genes Associated with Leaf Etiolation of a Cytoplasmic Male Sterility Line in Chinese Cabbage (Brassica Rapa L. ssp. Pekinensis). Int. J. Mol. Sci. 2018, 19, 922. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S.; Delhomme, N.; García-Gil, M.R. Global gene expression analysis in etiolated and de-etiolated seedlings in conifers. PLoS ONE 2019, 14, e0219272. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Dong, N.; Chen, Y.; Zhang, Y.; Hao, Y.; Qi, J. Comparative transcriptome analyses provide novel insights into etiolated shoot development of walnut (Juglans regia L.). Planta 2020, 252, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Xu, Z.-S.; Wang, F.; Li, M.-Y.; Ma, J.; Xiong, A.-S. Effects of abiotic stresses on the expression of Lhcb1 gene and photosynthesis of Oenanthe javanica and Apium graveolens. Biol. Plant 2014, 58, 256–264. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhriss, M.; Ben Abdullah, F. Exogenous Proline Effects on Photosynthetic Performance and Antioxidant Defense System of Young Olive Tree. J. Agric. Food Chem. 2010, 58, 4216–4222. [Google Scholar] [CrossRef]
- Jenny, A.; Mark, W.; Robin, G.W.; Caroline, A.H.; Alexander, V.R.; Peter, H.; Stefan, J. Absence of the Lhcb1 and Lhcb2 proteins of the light-harvesting complex of photosystem II—effects on photosynthesis, grana stacking and fitness. Plant J. 2003, 35, 350–361. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, S.; Li, X.; Wu, H.; Shan, X.; Wan, Y. Expression of Genes in New Sprouts of Cunninghamia lanceolata Grown Under Dark and Light Conditions. J. Plant Growth Regul. 2019, 39, 481–491. [Google Scholar] [CrossRef]
- Nelson, N.; Yocum, C.F. STRUCTURE AND FUNCTION OF PHOTOSYSTEMS I AND II. Annu. Rev. Plant Biol. 2006, 57, 521–565. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Lyu, Y.; Dong, X.; Huang, L.; He, K. Photosynthetic characteristics of Koelreuteria bipinnata var. integrifoliola and it’s natural yellow mutant ‘Jinyan’’. J. Nanjing For. Univ. 2016, 40, 74–80. [Google Scholar]
- Lyu, Y.; Dong, X.; Huang, L.; Huang, L. De novo assembly of Koelreuteria transcriptome and analysis of major gene related to leaf etiolation. S. Afr. J. Bot. 2017, 113, 355–361. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.-J.; Zhang, J.-Z. Aux/IAA Gene Family in Plants: Molecular Structure, Regulation, and Function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Barbez, E.; Kubeš, M.; Rolčík, J.; Béziat, C.; Pěnčík, A.; Wang, B.; Rosquete, M.R.; Zhu, J.; Dobrev, P.I.; Lee, Y.; et al. A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 2012, 485, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.C.J.; Peeters, A.J.M.; Pierik, R. Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc. Natl. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef] [Green Version]
- Ueguchi-Tanaka, M.; Ashikari, M.; Nakajima, M.; Itoh, H.; Katoh, E.; Kobayashi, M.; Chow, T.-Y.; Hsing, Y.-I.C.; Kitano, H.; Yamaguchi, I.; et al. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature 2005, 437, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Liao, L.; Jiang, C.; Desnos, T.; Bartlett, J.; Fu, X.; Harberd, N.P. DELLAs Contribute to Plant Photomorphogenesis. Plant Physiol. 2007, 143, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Zhu, B. Evolutionary analysis of three gibberellin oxidase genesin rice, Arabidopsis, and soybean. Gene 2011, 473, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.L.; Hecht, V.; Schoor, J.K.V.; Davidson, S.E.; Ross, J.J. Light Regulation of Gibberellin Biosynthesis in Pea Is Mediated through the COP1/HY5 Pathway. Plant Cell 2009, 21, 800–813. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Sheen, J.; Müller, B. Cytokinin Signaling Networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef] [Green Version]
- Carabelli, M.; Possenti, M.; Sessa, G.; Ciolfi, A.; Sassi, M.; Morelli, G.; Ruberti, I. Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes Dev. 2007, 21, 1863–1868. [Google Scholar] [CrossRef]
- Moncaleán, P.; Rodríguez, A.; Fernández, B. Plant growth regulators as putative physiological markers of developmental stage in Prunus persica. Plant Growth Regul. 2002, 36, 27–29. [Google Scholar] [CrossRef]
- Andrés, H.; Fernández, B.; Rodríguez, R. Phytohormone contents in Corylus avellana and their relationship to age and other developmental processes. Plant Cell Tissue Organ Cult. (PCTOC) 2002, 70, 173–180. [Google Scholar] [CrossRef]
- Valdés, A.E.; Centeno, M.L.; Fernández, B. Age-related changes in the hormonal status of Pinus radiata needle fascicle meristems. Plant Sci. 2004, 167, 373–378. [Google Scholar] [CrossRef]
- Materán, M.E.; Fernández, M.; Valenzuela, S.; Saez, K.; Seemann, P.; Sanchezolate, M.; Rios, D. Abscisic acid and 3-indolacetic acid levels during the reinvigoration process of Pinus radiata D. Don adult material. Plant Growth Regul. 2009, 59, 171–177. [Google Scholar] [CrossRef]
- Dubois, M.; Broeck, L.V.D.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Chao, Q.; Rothenberg, M.; Solano, R.; Roman, G.; Terzaghi, W.; Ecker, J.R. Activation of the Ethylene Gas Response Pathway in Arabidopsis by the Nuclear Protein ETHYLENE-INSENSITIVE3 and Related Proteins. Cell 1997, 89, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Solano, R.; Stepanova, A.; Chao, Q.; Ecker, J.R. Nuclear events in ethylene signaling: A transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhu, B.; Xu, W.; Zhu, H.; Chen, A.; Xie, Y.; Shao, Y.; Luo, Y. LeERF1 positively modulated ethylene triple response on etiolated seedling, plant development and fruit ripening and softening in tomato. Plant Cell Rep. 2007, 26, 1999–2008. [Google Scholar] [CrossRef]
- Lu, N.; Dai, L.; Luo, Z.; Wang, S.; Wen, Y.; Duan, H.; Hou, R.; Sun, Y.; Li, Y. Characterization of the Transcriptome and Gene Expression of Tetraploid Black Locust Cuttings in Response to Etiolation. Genes 2017, 8, 345. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-F.; He, J.-X. BZR1 Interacts with HY5 to Mediate Brassinosteroid- and Light-Regulated Cotyledon Opening in Arabidopsis in Darkness. Mol. Plant 2015, 9, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Kozuka, T.; Kobayashi, J.; Horiguchi, G.; Demura, T.; Sakakibara, H.; Tsukaya, H.; Nagatani, A. Involvement of Auxin and Brassinosteroid in the Regulation of Petiole Elongation under the Shade. Plant Physiol. 2010, 153, 1608–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; You, J.; Chan, Z. Identification and characterization of TIFY family genes in Brachypodium distachyon. J. Plant Res. 2015, 128, 995–1005. [Google Scholar] [CrossRef]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, M.O.; Moore, M.; König, K.; Pecher, P.; Alsharafa, K.; Lee, J.; Dietz, K.-J. Fast Retrograde Signaling in Response to High Light Involves Metabolite Export, MITOGEN-ACTIVATED PROTEIN KINASE6, and AP2/ERF Transcription Factors in Arabidopsis. Plant Cell 2014, 26, 1151–1165. [Google Scholar] [CrossRef] [Green Version]
- Huysmans, M.; Buono, R.A.; Skorzinski, N.; Radio, M.C.; De Winter, F.; Parizot, B.; Mertens, J.; Karimi, M.; Fendrych, M.; Nowack, M.K. NAC Transcription Factors ANAC087 and ANAC046 Control Distinct Aspects of Programmed Cell Death in the Arabidopsis Columella and Lateral Root Cap. Plant Cell 2018, 30, 2197–2213. [Google Scholar] [CrossRef] [Green Version]
- Kam, J.; Gresshoff, P.M.; Shorter, R.; Xue, G.-P. The Q-type C2H2 zinc finger subfamily of transcription factors in Triticum aestivum is predominantly expressed in roots and enriched with members containing an EAR repressor motif and responsive to drought stress. Plant Mol. Biol. 2008, 67, 305–322. [Google Scholar] [CrossRef] [PubMed]
- He, G.-H.; Xu, J.-Y.; Wang, Y.-X.; Liu, J.-M.; Li, P.-S.; Chen, M.; Ma, Y.-Z.; Xu, Z.-S. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.-T.; Zhao, M.-J.; Wang, C.-T.; Gao, Y.; Wang, Y.-X.; Liu, Y.-W.; Chen, M.; Chen, J.; Zhou, Y.-B.; Xu, Z.-S.; et al. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18, 1–18. [Google Scholar] [CrossRef]
- Wang, C.-T.; Ru, J.-N.; Liu, Y.-W.; Li, M.; Zhao, D.; Yang, J.-F.; Fu, J.-D.; Xu, Z.-S. Maize WRKY Transcription Factor ZmWRKY106 Confers Drought and Heat Tolerance in Transgenic Plants. Int. J. Mol. Sci. 2018, 19, 3046. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.-S.; Chen, M.; Li, L.-C.; Ma, Y.-Z. Functions of the ERF transcription factor family in plants. Botany 2008, 86, 969–977. [Google Scholar] [CrossRef]
- Yang, Z.; Tian, L.; Latoszek-Green, M.; Brown, D.; Wu, K. Arabidopsis ERF4 is a transcriptional repressor capable of modulating ethylene and abscisic acid responses. Plant Mol. Biol. 2005, 58, 585–596. [Google Scholar] [CrossRef]
- Brown, R.L.; Kazan, K.; McGrath, K.; Maclean, D.J.; Manners, J.M. A Role for the GCC-Box in Jasmonate-Mediated Activation of the PDF1.2 Gene of Arabidopsis. Plant Physiol. 2003, 132, 1020–1032. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.; Briggs, W.R. Blue Light Sensing in Higher Plants. J. Biol. Chem. 2001, 276, 11457–11460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 Interacts with CIB1 to Regulate Transcription and Floral Initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterlund, M.T.; Hardtke, C.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, B.; Sun, M.; Li, Y.; Kawabata, S. UV-A Light Induces Anthocyanin Biosynthesis in a Manner Distinct from Synergistic Blue + UV-B Light and UV-A/Blue Light Responses in Different Parts of the Hypocotyls in Turnip Seedlings. Plant Cell Physiol. 2012, 53, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raineri, J.; Ribichich, K.F.; Chan, R.L. The sunflower transcription factor HaWRKY76 confers drought and flood tolerance to Arabidopsis thaliana plants without yield penalty. Plant Cell Rep. 2015, 34, 2065–2080. [Google Scholar] [CrossRef]
- Tamang, B.G.; Fukao, T. Plant Adaptation to Multiple Stresses during Submergence and Following Desubmergence. Int. J. Mol. Sci. 2015, 16, 30164–30180. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Huang, X.; Li, G.; Ji, Q.; Zhou, K.; Zhu, G.; Ke, W.; Hou, H.; Zhu, H.; Yang, J. Comparative Transcriptomic Analysis Provides Novel Insights into the Blanched Stem of Oenanthe javanica. Plants 2021, 10, 2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112484
Kumar S, Huang X, Li G, Ji Q, Zhou K, Zhu G, Ke W, Hou H, Zhu H, Yang J. Comparative Transcriptomic Analysis Provides Novel Insights into the Blanched Stem of Oenanthe javanica. Plants. 2021; 10(11):2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112484
Chicago/Turabian StyleKumar, Sunjeet, Xinfang Huang, Gaojie Li, Qun Ji, Kai Zhou, Guopeng Zhu, Weidong Ke, Hongwei Hou, Honglian Zhu, and Jingjing Yang. 2021. "Comparative Transcriptomic Analysis Provides Novel Insights into the Blanched Stem of Oenanthe javanica" Plants 10, no. 11: 2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112484