
Biodiversity of Root Endophytic Fungi from Oxyria sinensis Grown in Metal-Polluted and Unpolluted Soils in Yunnan Province, Southwestern China


Abstract
:1. Introduction
2. Results
2.1. Chemical Properties of the Soil
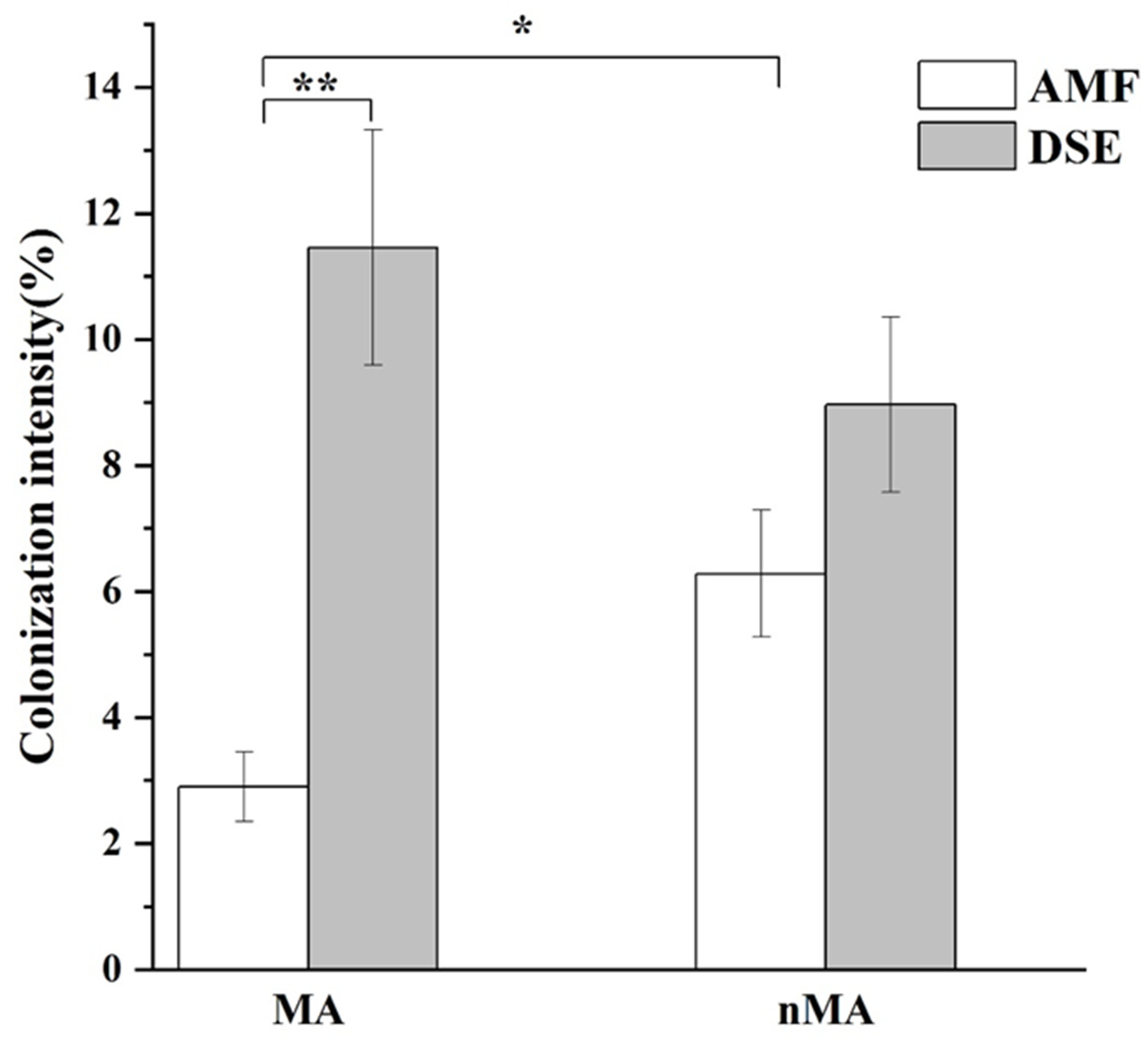
2.2. DSE and AMF Colonization
2.3. DSE Isolation and Identification
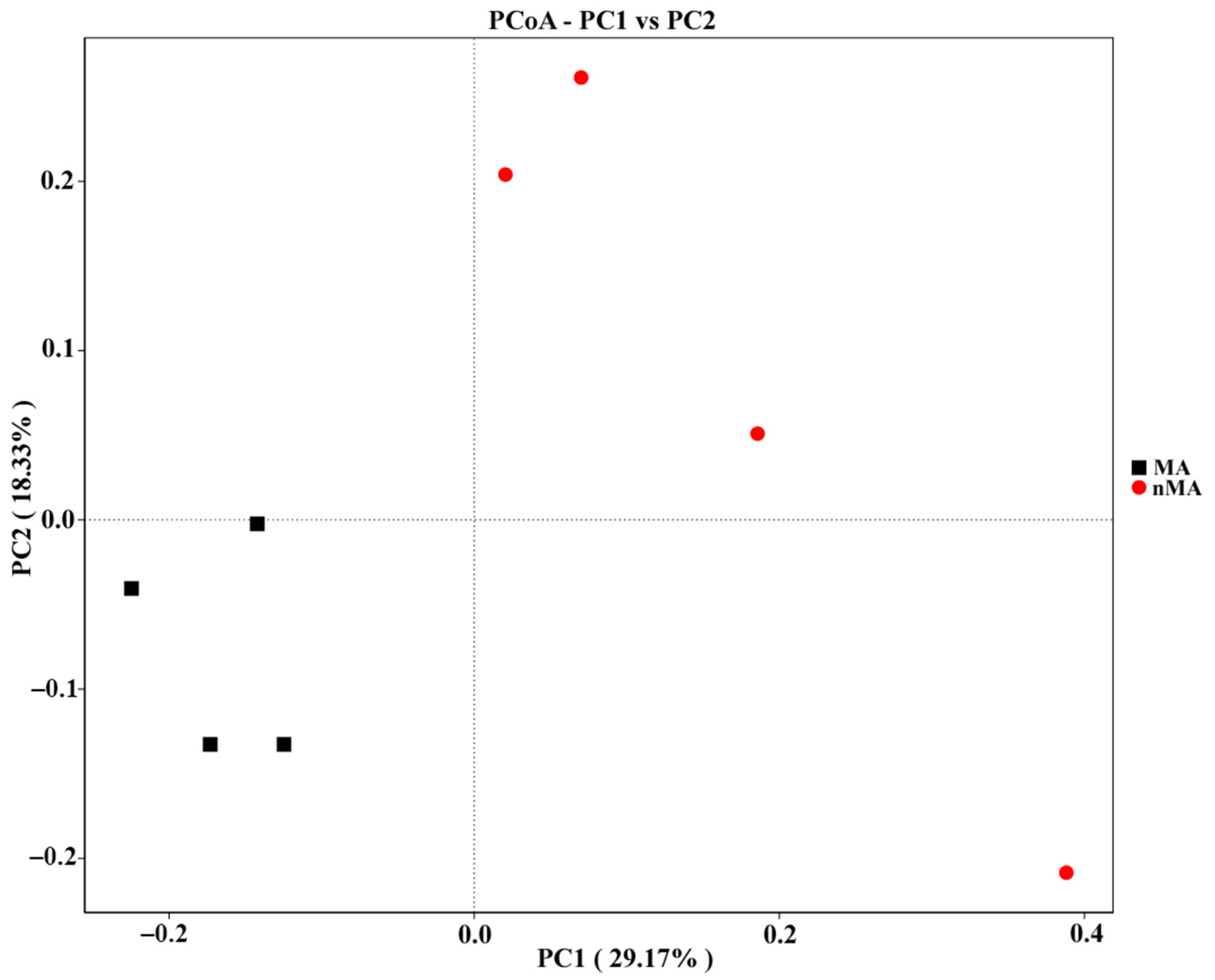
2.4. Endophytic Fungi Diversity and Community Structure in O. sinensis Roots
3. Discussion
4. Materials and Methods
4.1. Sample Sites and Collection
4.2. Soil Analysis
4.3. DSE and AMF Colonization
4.4. DSE Isolation and Identification
4.5. Root-Associated Fungal Assemblage Using Culture-Independent Methods
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buch, A.C.; Niemeyer, J.C.; Marques, E.D.; Silva-Filho, E.V. Ecological Risk Assessment of Trace Metals in Soils Affected by Mine Tailings. J. Hazard. Mater. 2020, 403, 123852. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy Metal Pollution in the Environment and Their Toxicological Effects on Humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of Heavy Metals—Concepts and Applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Liu, S.; Yang, B.; Liang, Y.; Xiao, Y.; Fang, J. Prospect of Phytoremediation Combined with Other Approaches for Remediation of Heavy Metal-Polluted Soils. Environ. Sci. Pollut. Res. 2020, 27, 16069–16085. [Google Scholar] [CrossRef] [PubMed]
- Tordoff, G.; Baker, A.; Willis, A. Current Approaches to the Revegetation and Reclamation of Metalliferous Mine Wastes. Chemosphere 2000, 41, 219–228. [Google Scholar] [CrossRef]
- Domka, A.M.; Rozpaądek, P.; Turnau, K. Are Fungal Endophytes Merely Mycorrhizal Copycats? The Role of Fungal Endophytes in the Adaptation of Plants to Metal Toxicity. Front. Microbiol. 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhu, L.L.; Li, T.; Zhao, Z.W. Mutualism between Dark Septate Endophytes (DSEs) and Their Host Plants under Metal Stress: A Case Study. All Life 2021, 14, 667–677. [Google Scholar] [CrossRef]
- Hui, F.; Liu, J.; Gao, Q.; Lou, B. Piriformosporaindica Confers Cadmium Tolerance in Nicotiana tabacum. J. Environ. Sci. 2015, 37, 184–191. [Google Scholar] [CrossRef]
- He, Y.; Fan, X.; Zhang, G.; Li, B.; Li, T.; Zu, Y.; Zhan, F. Effects of Arbuscular Mycorrhizal Fungi and Dark Septate Endophytes on Maize Performance and Root Traits under a High Cadmium Stress. S. Afr. J. Bot. 2019, 134, 415–423. [Google Scholar] [CrossRef]
- Ultra, V.U., Jr.; Manyiwa, T. Influence of Mycorrhiza and Fly Ash on the Survival, Growth and Heavy Metal Accumulation in Three Acacia Species Grown in Cu–Ni Mine Soil. Environ. Geochem. Health 2021, 43, 1337–1353. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, S.; Wang, J.; Zhang, X.; Shen, Z.; Shi, L.; Chen, Y. The Great Potential for Phytoremediation of Abandoned Tailings Pond Using Ectomycorrhizal Pinus sylvestris. Sci. Total Environ. 2020, 719, 137475. [Google Scholar] [CrossRef]
- Gange, A.C.; Brown, V.K.; Sinclair, G.S. Vesicular-Arbuscular Mycorrhizal Fungi: A Determinant of Plant Community Structure in Early Succession. Funct. Ecol. 1993, 7, 616. [Google Scholar] [CrossRef]
- Koziol, L.; Bever, J.D. Mycorrhizal Feedbacks Generate Positive Frequency Dependence Accelerating Grassland Succession. J. Ecol. 2018, 107, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Pérez, R.; Tapia, Y.; Antilén, M.; Casanova, M.; Vidal, C.; Santander, C.; Aponte, H.; Cornejo, P. Interactive Effect of Compost Application and Inoculation with the Fungus Claroideoglomus claroideum in Oenothera picensis Plants Growing in Mine Tailings. Ecotoxicol. Environ. Saf. 2020, 208, 111495. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Kupriynanov, A.N.; Manakov, A.; Betekhtina, A.A.; Prasad, M.N.V. Mycorrhizal Plants’ Accelerated Revegetation on Coal Mine Overburden in the Dry Steppes of Kazakhstan. In Environmental Sustainability; Springer: New Delhi, India, 2014; pp. 265–280. [Google Scholar] [CrossRef]
- Durand, A.; Leglize, P.; Benizri, E. Are Endophytes Essential Partners for Plants and What Are the Prospects for Metal Phytoremediation? Plant Soil 2021, 460, 1–30. [Google Scholar] [CrossRef]
- Yanqun, Z.; Yuan, L.; Schvartz, C.; Langlade, L.; Fan, L. Accumulation of Pb, Cd, Cu and Zn in Plants and Hyperaccumulator Choice in Lanping Lead-Zinc Mine Area, China. Environ. Int. 2004, 30, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of Native Plants with Phytoremediation Potential for Highly Contaminated Mediterranean Soil Restoration: Tools for a Non-Destructive and Integrative Approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Lei, D.; Duan, C. Restoration Potential of Pioneer Plants Growing on Lead-Zinc Mine Tailings in Lanping, Southwest China. J. Environ. Sci. 2008, 20, 1202–1209. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, Y.P. Reproductive Allocation in a Dioecious Perennial Oxyria sinensis (Polygonaceae) along Altitudinal Gradients. J. Syst. Evol. 2008, 46, 830–835. [Google Scholar] [CrossRef]
- Li, Y.; Zu, Y.; Fang, Q.; Chen, H.; Schvartz, C. Characteristics of Heavy-Metal Tolerance and Growth in Two Ecotypes of Oxyria sinensis Hemsl. Grown on Huize Lead-Zinc Mining Area in Yunnan Province, China. Commun. Soil Sci. Plant Anal. 2013, 44, 2428–2442. [Google Scholar] [CrossRef]
- Ruotsalainen, A.L.; Markkola, A.; Kozlov, M.V. Root Fungal Colonisation in Deschampsia flexuosa: Effects of Pollution and Neighbouring Trees. Environ. Pollut. 2007, 147, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, C.; Blaudez, D.; Beguiristain, T.; Chalot, M.; Leyval, C. Co-Inoculation of Lolium perenne with Funneliformis mosseae and the Dark Septate Endophyte Cadophora sp. in a Trace Element-Polluted Soil. Mycorrhiza 2018, 28, 301–314. [Google Scholar] [CrossRef] [Green Version]
- Gucwa-Przepióra, E.; Nadgórska-Socha, A.; Fojcik, B.; Chmura, D. Enzymatic Activities and Arbuscular Mycorrhizal Colonization of Plantago lanceolata and Plantago major in a Soil Root Zone under Heavy Metal Stress. Environ. Sci. Pollut. Res. 2015, 23, 4742–4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deram, A.; Languereau-Leman, F.; Howsam, M.; Petit, D.; Van Haluwyn, C. Seasonal Patterns of Cadmium Accumulation in Arrhenatherum elatius (Poaceae): Influence of Mycorrhizal and Endophytic Fungal Colonisation. Soil Biol. Biochem. 2008, 40, 845–848. [Google Scholar] [CrossRef]
- Regvar, M.; Likar, M.; Piltaver, A.; Kugonič, N.; Smith, J.E. Fungal Community Structure under Goat Willows (Salix caprea L.) Growing at Metal Polluted Site: The Potential of Screening in a Model Phytostabilisation Study. Plant Soil 2009, 330, 345–356. [Google Scholar] [CrossRef]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Schultz, N.; Balieiro, F.D.C.; Medeiros, P.S.; Santos, L.A.; Xavier, G.R.; Zilli, J.E. Dark Septate Endophytic Fungi Help Tomato to Acquire Nutrients from Ground Plant Material. Front. Microbiol. 2017, 8, 2437. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Linares, D.R.; Grosch, R.; Restrepo, S.; Krumbein, A.; Franken, P. Effects of Dark Septate Endophytes on Tomato Plant Performance. Mycorrhiza 2010, 21, 413–422. [Google Scholar] [CrossRef]
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.-W. Evidence for a Dark Septate Endophyte (Exophiala pisciphila, H93) Enhancing Phosphorus Absorption by Maize Seedlings. Plant Soil 2020, 452, 249–266. [Google Scholar] [CrossRef]
- Ruthrof, K.X.; Fontaine, J.; Hopkins, A.J.; McHenry, M.P.; O’Hara, G.; McComb, J.; Hardy, G.E.; Howieson, J. Potassium Amendment Increases Biomass and Reduces Heavy Metal Concentrations in Lablab purpureus after Phosphate Mining. Land Degrad. Dev. 2017, 29, 398–407. [Google Scholar] [CrossRef]
- Addy, H.D.; Piercey, M.M.; Currah, R. Microfungal Endophytes in Roots. Can. J. Bot. 2005, 83, 1–13. [Google Scholar] [CrossRef]
- Jumpponen, A.; Trappe, J.M. Dark Septate Endophytes: A Review of Facultative Biotrophic Root-Colonizing Fungi. New Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Zhao, Z.-W.; Li, T. Dark Septate Endophytes and Their Ecological Function in Metal Mining. Sci. Sin. Vitae 2019, 49, 921–926. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Zhao, Z.-W. Colonization Characteristics and Composition of Dark Septate Endophytes (DSE) in a Lead and Zinc Slag Heap in Southwest China. Soil Sediment Contam. Int. J. 2013, 22, 532–545. [Google Scholar] [CrossRef]
- Cao, G.-H.; He, S.; Chen, D.; Li, T.; Zhao, Z.-W. EpABC Genes in the Adaptive Responses of Exophiala pisciphila to Metal Stress: Functional Importance and Relation to Metal Tolerance. Appl. Environ. Microbiol. 2019, 85, e01844-19. [Google Scholar] [CrossRef]
- Li, T.; Liu, M.; Zhang, X.; Zhang, H.; Sha, T.; Zhao, Z. Improved Tolerance of Maize (Zea mays L.) to Heavy Metals by Colonization of a Dark Septate Endophyte (DSE) Exophiala pisciphila. Sci. Total Environ. 2011, 409, 1069–1074. [Google Scholar] [CrossRef]
- Wang, J.-L.; Jun-Ling, W.; Liu, G.-Y.; Smith, J.M.; Zhao, Z.-W. Unraveling the Role of Dark Septate Endophyte (DSE) Colonizing Maize (Zea mays) under Cadmium Stress: Physiological, Cytological and Genic Aspects. Sci. Rep. 2016, 6, 22028. [Google Scholar] [CrossRef] [PubMed]
- Xiao, E.; Ning, Z.; Xiao, T.; Sun, W.; Qiu, Y.; Zhang, Y.; Chen, J.; Gou, Z.; Chen, Y. Variation in Rhizosphere Microbiota Correlates with Edaphic Factor in an Abandoned Antimony Tailing Dump. Environ. Pollut. 2019, 253, 141–151. [Google Scholar] [CrossRef]
- Davis, E.C.; Franklin, J.B.; Shaw, A.J.O.; Vilgalys, R. Endophytic Xylaria (Xylariaceae) among Liverworts and Angiosperms: Phylogenetics, Distribution, and Symbiosis. Am. J. Bot. 2003, 90, 1661–1667. [Google Scholar] [CrossRef]
- Wężowicz, K.; Rozpądek, P.; Turnau, K. Interactions of Arbuscular Mycorrhizal and Endophytic Fungi Improve Seedling Survival and Growth in Post-Mining Waste. Mycorrhiza 2017, 27, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Muthukumar, T.; Udaiyan, K.; Shanmughavel, P. Mycorrhiza in Sedges—An Overview. Mycorrhiza 2004, 14, 65–77. [Google Scholar] [CrossRef]
- Wagg, C.; Jansa, J.; Schmid, B.; van der Heijden, M.G.A. Belowground Biodiversity Effects of Plant Symbionts Support Aboveground Productivity. Ecol. Lett. 2011, 14, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Jansa, J.; Stadler, M.; Schmid, B.; van der Heijden, M.G.A. Mycorrhizal Fungal Identity and Diversity Relaxes Plant–Plant Competition. Ecology 2011, 92, 1303–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, B.G.; Wu, L.S. Studies of Historical Data Concerning Exploitation of Silver (Lead)-Zinc Deposits in Yunnan Province and Some Opinions on Further Mineral Exploration. Miner. Depos. 2002, 21, 303–309. [Google Scholar] [CrossRef]
- Yu, F.-K.; Huang, X.-H.; Duan, C.-Q.; He, S.-Z.; Zhang, G.-S.; Liu, C.-E.; Fu, D.-G.; Shao, H.-B. Impacts of Ageratina adenophora Invasion on Soil Physical-Chemical Properties of Eucalyptus Plantation and Implications for Constructing Agro-Forest Ecosystem. Ecol. Eng. 2014, 64, 130–135. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, J.; Cai, C.; Cai, Z.; Li, X.; Wang, R. Coupling of Non-Point Source Pollution and Soil Characteristics Covered by Phyllostachys edulis Stands in Hilly Water Source Area. J. Environ. Manag. 2020, 268, 110657. [Google Scholar] [CrossRef]
- Watanabe, Y.; Itanna, F.; Izumi, Y.; Awala, S.K.; Fujioka, Y.; Tsuchiya, K.; Iijima, M. Cattle Manure and Intercropping Effects on Soil Properties and Growth and Yield of Pearl Millet and Cowpea in Namibia. J. Crop. Improv. 2019, 33, 395–409. [Google Scholar] [CrossRef]
- Jing, Y.; Li, T.; Cui, H.; Li, L.; Allen, S.; Chen, L.; Li, Y.; Zhao, Z. Shifts in the Arbuscular Mycorrhizal Fungal Community Composition of Betula alnoides along Young, Middle-Aged Plantation and Adjacent Natural Forest. iFor. Biogeosci. For. 2020, 13, 447–455. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A New Method Which Gives an Objective Measure of Colonization of Roots by Vesicular-Arbuscular Mycorrhizal Fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Olsrud, M.; Michelsen, A.; Wallander, H. Ergosterol Content in Ericaceous Hair Roots Correlates with Dark Septate Endophytes but Not with Ericoid Mycorrhizal Colonization. Soil Biol. Biochem. 2007, 39, 1218–1221. [Google Scholar] [CrossRef]
- Silvani, V.; Fracchia, S.; Fernández, L.; Pérgola, M.; Godeas, A. A Simple Method to Obtain Endophytic Microorganisms from Field-Collected Roots. Soil Biol. Biochem. 2008, 40, 1259–1263. [Google Scholar] [CrossRef]
- Xu, R.; Li, T.; Cui, H.; Wang, J.; Yu, X.; Ding, Y.; Wang, C.; Yang, Z.; Zhao, Z. Diversity and Characterization of Cd-Tolerant Dark Septate Endophytes (DSEs) Associated with the roots of Nepal Alder (Alnus nepalensis) in a Metal Mine Tailing of Southwest China. Appl. Soil Ecol. 2015, 93, 11–18. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi. Mycologia 1972, 64, 930. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990. [Google Scholar]
- Huang, Y.; Kuang, Z.; Wang, W.; Cao, L. Exploring Potential Bacterial and Fungal Biocontrol Agents Transmitted from Seeds to Sprouts of Wheat. Biol. Control 2016, 98, 27–33. [Google Scholar] [CrossRef]
- Kutner, M.H.; Nachtsheim, C.J.; Neter, J.; Li, W. Diagnostics and Remedial Measures Applied Linear Statistical Models, 5th ed.; McGraw-Hill/Irwin: New York, NY, USA, 2004; p. 108. ISBN 0-07-238688-6. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Physiochemical Properties (mg kg−1) | Total Metal Concentration (mg kg−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Alkaline Hydrolysis N | Organic Matter | Available P | Available K | pH | Pb | Zn | Cd | |
| MA | 60.4 ± 13.10 | 16.9 ± 3.47 b | 19.7 ± 0.84 a | 184.9 ± 25.03 | 6.2 ± 0.06 | 191.9 ± 37.67 b | 719.5 ± 210.00 b | 1.9 ± 0.51 b |
| nMA | 42.5 ± 2.80 | 5.3 ± 0.72 a | 26.1 ± 1.06 b | 232.0 ± 8.18 | 6.2 ± 0.03 | 31.9 ± 3.87 a | 192.5 ± 25.40 a | 0.0 ± 0.00 a |
| Sites | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|
| MA | 235.1 ± 18.23 | 231.7 ± 16.21 | 2.7 ± 0.31 | 0.7 ± 0.08 |
| nMA | 196.3 ± 32.88 | 191.4 ± 31.70 | 3.0 ± 0.22 | 0.8 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Ding, Y.; Li, X.; Xiao, Y.; Zhao, Z.; Li, T. Biodiversity of Root Endophytic Fungi from Oxyria sinensis Grown in Metal-Polluted and Unpolluted Soils in Yunnan Province, Southwestern China. Plants 2021, 10, 2731. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122731
Zhu M, Ding Y, Li X, Xiao Y, Zhao Z, Li T. Biodiversity of Root Endophytic Fungi from Oxyria sinensis Grown in Metal-Polluted and Unpolluted Soils in Yunnan Province, Southwestern China. Plants. 2021; 10(12):2731. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122731
Chicago/Turabian StyleZhu, Meiyan, Yanhua Ding, Xuejiao Li, Yuqing Xiao, Zhiwei Zhao, and Tao Li. 2021. "Biodiversity of Root Endophytic Fungi from Oxyria sinensis Grown in Metal-Polluted and Unpolluted Soils in Yunnan Province, Southwestern China" Plants 10, no. 12: 2731. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122731