
Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Assessment of Body Weight and Prostate, Liver, and Heart Index
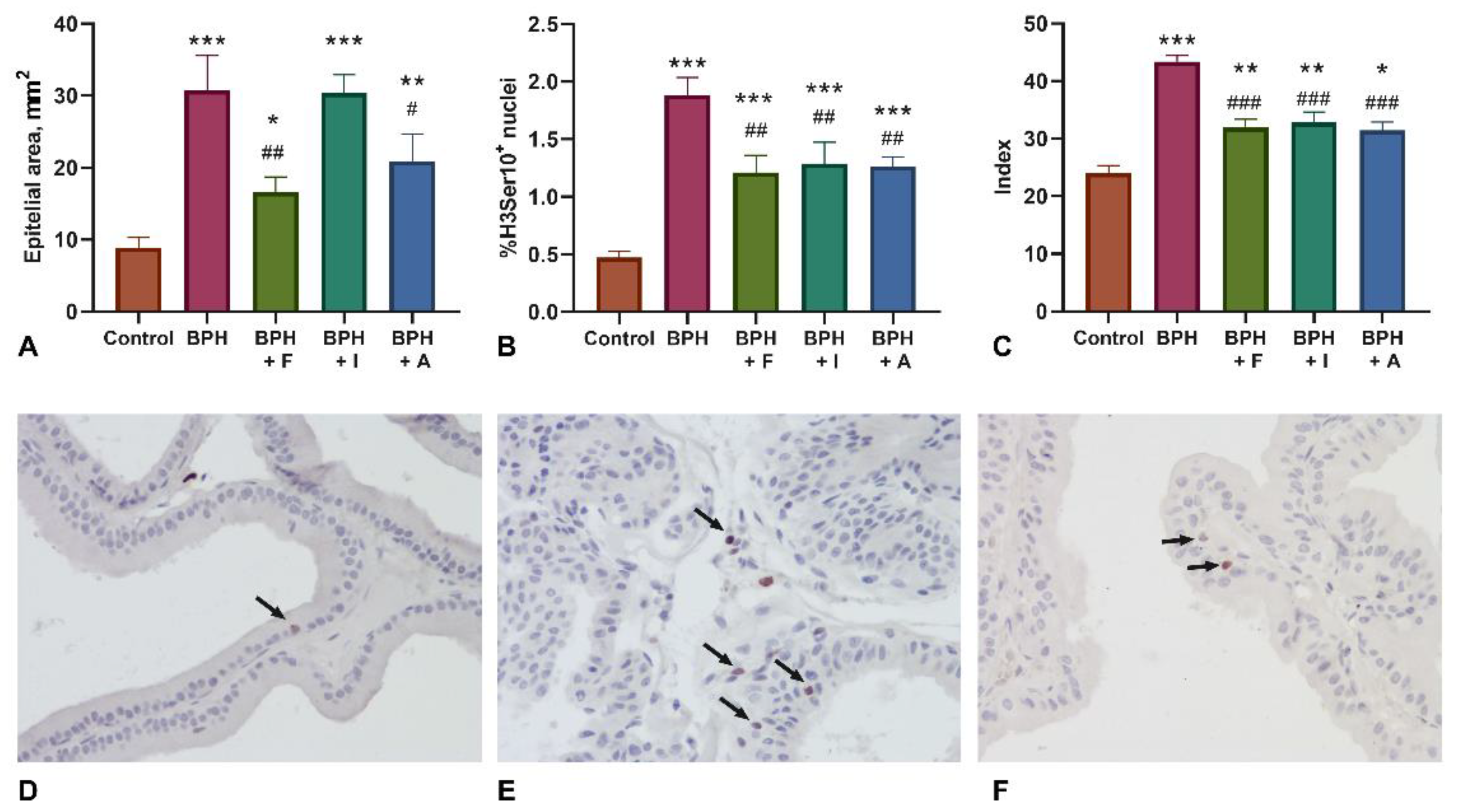
2.2. Histopathological Examination and Assessment of Proliferation Markers (H3Ser10)
2.3. Assessment of Biochemical Analysis
2.4. Assessment of Blood Count, Antioxidant Activity, Biochemistry, and ELISA Assay
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. NADES (Natural Deep Eutectic Solvent) Extract of Pueraria Lobata Roots
4.1.2. Extract of Phaffia rhodozyma
4.2. Animals
4.3. Experimental Design
4.4. Measurement of Prostate, Liver, and Heart Index
4.5. Histopathological Evaluation and Immunohistochemical Detection of H3Ser10
4.6. Blood Count
4.7. Prostate Tissue Probe Preparation
4.8. Biochemical Analysis, Antioxidant Activity Evaluation, and Testosterone Assay
4.9. Statistical Analysis
4.10. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, W.; Guo, Y.; Zhang, D.; Tian, Y.; Zhang, X. The Prevalence of Benign Prostatic Hyperplasia in Mainland China: Evidence from Epidemiological Surveys. Sci. Rep. 2015, 5, 13546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Fang, X.; Ma, Y.; Xianyu, J. Benign Prostatic Hyperplasia and the Risk of Prostate Cancer and Bladder Cancer: A Meta-Analysis of Observational Studies. Medicine 2016, 95, e3493. [Google Scholar] [CrossRef]
- Ng, M.; Baradhi, K.M. Benign Prostatic Hyperplasia. In StatPearls; StatPearls Publishing: Treasure Island, Finland, 2021. [Google Scholar]
- Speakman, M.; Kirby, R.; Doyle, S.; Ioannou, C. Burden of Male Lower Urinary Tract Symptoms (LUTS) Suggestive of Benign Prostatic Hyperplasia (BPH)—Focus on the UK: Burden of Male LUTS Suggestive of BPH. BJU Int. 2015, 115, 508–519. [Google Scholar] [CrossRef]
- Carson, C.; Rittmaster, R. The Role of Dihydrotestosterone in Benign Prostatic Hyperplasia. Urology 2003, 61, 2–7. [Google Scholar] [CrossRef]
- Randall, V.A. 9 Role of 5α-Reductase in Health and Disease. Baillière’s Clin. Endocrinol. Metab. 1994, 8, 405–431. [Google Scholar] [CrossRef]
- Aydin, A.; Arsova-Sarafinovska, Z.; Sayal, A.; Eken, A.; Erdem, O.; Erten, K.; Özgök, Y.; Dimovski, A. Oxidative Stress and Antioxidant Status in Non-Metastatic Prostate Cancer and Benign Prostatic Hyperplasia. Clin. Biochem. 2006, 39, 176–179. [Google Scholar] [CrossRef]
- Aryal, M.; Pandeya, A.; Gautam, N.; Baral, N.; Lamsal, M.; Majhi, S.; Chandra, L.; Pandit, R.; Das, B.K.L. Oxidative Stress in Benign Prostate Hyperplasia. Nepal Med. Coll. J. 2007, 9, 222–224. [Google Scholar]
- Crocetto, F.; Boccellino, M.; Barone, B.; Di Zazzo, E.; Sciarra, A.; Galasso, G.; Settembre, G.; Quagliuolo, L.; Imbimbo, C.; Boffo, S. The Crosstalk between Prostate Cancer and Microbiota Inflammation: Nutraceutical Products Are Useful to Balance This Interplay? Nutrients 2020, 12, 2648. [Google Scholar] [CrossRef]
- Pagano, E.; Laudato, M.; Griffo, M.; Capasso, R. Phytotherapy of Benign Prostatic Hyperplasia. A Minireview: Phytoterapy of Benign Prostatic Hyperplasia. Phytother. Res. 2014, 28, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.E.J.; Kinkade, A.; Tung, A.C.H.; Tejani, A.M. 5α-Reductase Inhibitors for Treatment of Benign Prostatic Hyperplasia: A Systematic Review and Meta-Analysis. Can. J. Hosp. Pharm. 2017, 70, 113–119. [Google Scholar] [CrossRef]
- Salisbury, B.H.; Tadi, P. 5 Alpha Reductase Inhibitors. In StatPearls; StatPearls Publishing: Treasure Island, Finland, 2021. [Google Scholar]
- Kim, T.-H.; Lim, H.-J.; Kim, M.-S.; Lee, M.S. Dietary Supplements for Benign Prostatic Hyperplasia: An Overview of Systematic Reviews. Maturitas 2012, 73, 180–185. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Allkanjari, O.; Busetto, G.M.; Cai, T.; Larganà, G.; Magri, V.; Perletti, G.; Robustelli Della Cuna, F.S.; Russo, G.I.; Stamatiou, K.; et al. Nutraceutical Treatment and Prevention of Benign Prostatic Hyperplasia and Prostate Cancer. Arch. Ital. Urol. Androl. 2019, 91, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Crocetto, F.; Di Zazzo, E.; Buonerba, C.; Aveta, A.; Pandolfo, S.D.; Barone, B.; Trama, F.; Caputo, V.F.; Scafuri, L.; Ferro, M. Kaempferol, Myricetin and Fisetin in Prostate and Bladder Cancer: A Systematic Review of the Literature. Nutrients 2021, 13, 3750. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.-R.; Chung, K.-S.; Kim, H.-J.; An, H.-J. Chinese Skullcap (Scutellaria Baicalensis Georgi) Inhibits Inflammation and Proliferation on Benign Prostatic Hyperplasia in Rats. J. Ethnopharmacol. 2019, 235, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Campolo, M.; Cordaro, M.; Siracusa, R.; Filippone, A.; Esposito, E.; Cuzzocrea, S. Effects of Different Natural Extracts in an Experimental Model of Benign Prostatic Hyperplasia (BPH). Inflamm. Res. 2018, 67, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Roca, P. Phytoestrogens for Cancer Prevention and Treatment. Biology 2020, 9, 427. [Google Scholar] [CrossRef]
- Weber, K.; Setchell, K.; Stocco, D.; Lephart, E. Dietary Soy-Phytoestrogens Decrease Testosterone Levels and Prostate Weight without Altering LH, Prostate 5alpha-Reductase or Testicular Steroidogenic Acute Regulatory Peptide Levels in Adult Male Sprague-Dawley Rats. J. Endocrinol. 2001, 170, 591–599. [Google Scholar] [CrossRef]
- Mitsunari, K.; Miyata, Y.; Matsuo, T.; Mukae, Y.; Otsubo, A.; Harada, J.; Kondo, T.; Matsuda, T.; Ohba, K.; Sakai, H. Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia. Molecules 2021, 26, 450. [Google Scholar] [CrossRef]
- Bultosa, G. Functional Foods: Overview. In Encyclopedia of Food Grains; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–10. ISBN 978-0-12-394786-4. [Google Scholar]
- Shashank, A.; Ramakrishna, D.; Kiran, S.; Kosturkova, G.; Ravishankar, G.A. Nutraceutical Potential of Soybean: Review. Asian J. Clin. Nutr. 2015, 7, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The Potential Health Effects of Dietary Phytoestrogens: Potential Health Effects of Dietary Phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, F.H.; Li, Y. Soy Isoflavones and Cancer Prevention. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef]
- Yousef, M.I.; Kamel, K.I.; Esmail, A.M.; Baghdadi, H.H. Antioxidant Activities and Lipid Lowering Effects of Isoflavone in Male Rabbits. Food Chem. Toxicol. 2004, 42, 1497–1503. [Google Scholar] [CrossRef]
- Lewis, J.G.; Nakajin, S.; Ohno, S.; Warnock, A.; Florkowski, C.M.; Elder, P.A. Circulating Levels of Isoflavones and Markers of 5alpha-Reductase Activity Are Higher in Japanese Compared with New Zealand Males: What Is the Role of Circulating Steroids in Prostate Disease? Steroids 2005, 70, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Batista, V.S.F.; Nunes, G.L.; Viegas, G.I.; Lucas, B.N.; Bochi, V.C.; Emanuelli, T.; Barin, J.S.; de Menezes, C.R.; da Rosa, C.S. Extraction, Characterization and Microencapsulation of Isoflavones from Soybean Molasses. Cienc. Rural 2020, 50, e20190341. [Google Scholar] [CrossRef]
- Masrudin, S.S.; Mohamad, J. Preventive Effect of Pueraria Mirifica on Testosterone-Induced Prostatic Hyperplasia in Sprague Dawley Rats. Andrologia 2015, 47, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, J.; Masrudin, S.S.; Alias, Z.; Muhamad, N.A. The Effects of Pueraria Mirifica Extract, Diadzein and Genistein in Testosterone-Induced Prostate Hyperplasia in Male Sprague Dawley Rats. Mol. Biol. Rep. 2019, 46, 1855–1871. [Google Scholar] [CrossRef] [PubMed]
- Soares, N.d.C.P.; Teodoro, A.J.; Lotsch, P.F.; Granjeiro, J.M.; Borojevic, R. Anticancer Properties of Carotenoids in Prostate Cancer. A Review. Histol. Histopathol. 2015, 30, 1143–1154. [Google Scholar] [CrossRef]
- Dulińska-Litewka, J.; Sharoni, Y.; Hałubiec, P.; Łazarczyk, A.; Szafrański, O.; McCubrey, J.A.; Gąsiorkiewicz, B.; Laidler, P.; Bohn, T. Recent Progress in Discovering the Role of Carotenoids and Their Metabolites in Prostatic Physiology and Pathology with a Focus on Prostate Cancer—A Review—Part I: Molecular Mechanisms of Carotenoid Action. Antioxidants 2021, 10, 585. [Google Scholar] [CrossRef]
- Puah, B.-P.; Jalil, J.; Attiq, A.; Kamisah, Y. New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules 2021, 26, 3888. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Shatunova, S.A.; Glukhareva, T.V.; Kovaleva, E.G. Effect of Different Sugar Sources on P. Rhodozyma Y1654 Growth and Astaxanthin Production. Agronomy Research 2020, 18, 1700–1716. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Glukhareva, T.V.; Danilova, I.G.; Kovaleva, E.G. Natural Antioxidants in Diabetes Treatment and Management: Prospects of Astaxanthin. Crit. Rev. Food Sci. Nutr. 2021, 1–24, [Online ahead of print]. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Ravishankar, G.A. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—a Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Gokare, R.A. Global Microalgal-Based Products for Industrial Applications. In Handbook of Algal Technologies and Phytochemicals; CRC Press: Boca Raton, FL, USA, 2019; pp. 267–278. ISBN 0-429-05789-X. [Google Scholar]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A Mechanistic Review on Its Biological Activities and Health Benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H. Multiple Mechanisms of Anti-Cancer Effects Exerted by Astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraone, I.; Sinisgalli, C.; Ostuni, A.; Armentano, M.F.; Carmosino, M.; Milella, L.; Russo, D.; Labanca, F.; Khan, H. Astaxanthin Anticancer Effects Are Mediated through Multiple Molecular Mechanisms: A Systematic Review. Pharmacol. Res. 2020, 155, 104689. [Google Scholar] [CrossRef]
- Anderson, M.L. A Preliminary Investigation of the Enzymatic Inhibition of 5alpha-Reduction and Growth of Prostatic Carcinoma Cell Line LNCap-FGC by Natural Astaxanthin and Saw Palmetto Lipid Extract in Vitro. J. Herb. Pharmacother. 2005, 5, 17–26. [Google Scholar] [CrossRef]
- Scolnik, M.D.; Servadio, C.; Abramovici, A. Comparative Study of Experimentally Induced Benign and Atypical Hyperplasia in the Ventral Prostate of Different Rat Strains. J. Androl. 1994, 15, 287–297. [Google Scholar] [CrossRef]
- Guo, W.; Bachman, E.; Vogel, J.; Li, M.; Peng, L.; Pencina, K.; Serra, C.; Sandor, N.L.; Jasuja, R.; Montano, M.; et al. The Effects of Short-Term and Long-Term Testosterone Supplementation on Blood Viscosity and Erythrocyte Deformability in Healthy Adult Mice. Endocrinology 2015, 156, 1623–1629. [Google Scholar] [CrossRef] [Green Version]
- Olsen, N.J.; Gu, X.; Kovacs, W.J. Bone Marrow Stromal Cells Mediate Androgenic Suppression of B Lymphocyte Development. J. Clin. Investig. 2001, 108, 1697–1704. [Google Scholar] [CrossRef]
- Shin, I.-S.; Lee, M.-Y.; Jung, D.-Y.; Seo, C.-S.; Ha, H.-K.; Shin, H.-K. Ursolic Acid Reduces Prostate Size and Dihydrotestosterone Level in a Rat Model of Benign Prostatic Hyperplasia. Food Chem. Toxicol. 2012, 50, 884–888. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, G.; Yan, Z.; Shang, X. The Effect of Xialiqi Capsule on Testosterone-Induced Benign Prostatic Hyperplasia in Rats. Evid.-Based Complementary Altern. Med. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.V.; Hermann, D.J.; Cunningham, G.R.; Wilson, T.H.; Morrill, B.B.; Hobbs, S. Marked Suppression of Dihydrotestosterone in Men with Benign Prostatic Hyperplasia by Dutasteride, a Dual 5alpha-Reductase Inhibitor. J. Clin. Endocrinol. Metab. 2004, 89, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Asare, G.A.; Ngala, R.A.; Afriyie, D.; Adjei, S.; Nyarko, A.; Anang-Quartey, Y.; Asiedu, B.; Doku, D.; Amoah, B.Y.; Bentum, K.; et al. Calcium—Magnesium Imbalance Implicated in Benign Prostatic Hyperplasia and Restoration by a Phytotherapeutic Drug—Croton Membranaceus Müll.Arg. BMC Complement. Altern. Med. 2017, 17, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tvedt, K.E.; Halgunset, J.; Kopstad, G.; Haugen, O.A. Intracellular Distribution of Calcium and Zinc in Normal, Hyperplastic, and Neoplastic Human Prostate: X-Ray Microanalysis of Freeze-Dried Cryosections. Prostate 1989, 15, 41–51. [Google Scholar] [CrossRef]
- Yaman, M.; Atici, D.; Bakırdere, S.; Akdeniz, İ. Comparison of Trace Metal Concentrations in Malign and Benign Human Prostate. J. Med. Chem. 2005, 48, 630–634. [Google Scholar] [CrossRef]
- Sun, Y.; Selvaraj, S.; Varma, A.; Derry, S.; Sahmoun, A.E.; Singh, B.B. Increase in Serum Ca2+/Mg2+ Ratio Promotes Proliferation of Prostate Cancer Cells by Activating TRPM7 Channels. J. Biol. Chem. 2013, 288, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afriyie, D.K.; Asare, G.A.; Bugyei, K.; Lin, J.; Peng, J.; Hong, Z. Mitochondria-Dependent Apoptogenic Activity of the Aqueous Root Extract of Croton Membranaceus against Human BPH-1 Cells. Genet. Mol. Res. 2015, 14, 149–162. [Google Scholar] [CrossRef]
- Lim, S.-R.; Kim, D.-W.; Sung, J.; Kim, T.H.; Choi, C.-H.; Lee, S.-J. Astaxanthin Inhibits Autophagic Cell Death Induced by Bisphenol A in Human Dermal Fibroblasts. Antioxidants 2021, 10, 1273. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Woo, M.; Kusuma, I.W.; Arung, E.T.; Yang, C.H.; Kim, Y. Inhibitory Effects of Isoflavonoids on Rat Prostate Testosterone 5α-Reductase. J. Acupunct. Meridian Stud. 2012, 5, 319–322. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.-X.; Zhu, X.-Y.; Huang, T.; Yang, J.; Cheng, Y.-D.; Xu, Y.-F. Effects of Kudzu Root plus Cinnamon Granules on prostatic hyperplasia in mice. Zhonghua Nan Ke Xue 2017, 23, 353–360. [Google Scholar]
- Aboushanab, S.A.; Khedr, S.M.; Gette, I.F.; Danilova, I.G.; Kolberg, N.A.; Ravishankar, G.A.; Ambati, R.R.; Kovaleva, E.G. Isoflavones Derived from Plant Raw Materials: Bioavailability, Anti-Cancer, Anti-Aging Potentials, and Microbiome Modulation. Crit. Rev. Food Sci. Nutr. 2021, 1–27, [Online ahead of print]. [Google Scholar] [CrossRef]
- DiSilvestro, R.A.; Mattern, C.; Wood, N.; Devor, S.T. Soy Protein Intake by Active Young Adult Men Raises Plasma Antioxidant Capacity without Altering Plasma Testosterone. Nutr. Res. 2006, 26, 92–95. [Google Scholar] [CrossRef]
- Nogueira-de-Almeida, C.A.; Ferraz, I.S.; Ued, F.d.V.; Almeida, A.C.F.; Ciampo, L.A.D. Impact of Soy Consumption on Human Health: Integrative Review. Braz. J. Food Technol. 2020, 23, 1–17. [Google Scholar] [CrossRef]
- Wong, S.Y.S.; Lau, W.W.Y.; Leung, P.C.; Leung, J.C.S.; Woo, J. The Association between Isoflavone and Lower Urinary Tract Symptoms in Elderly Men. Br. J. Nutr. 2007, 98, 1237–1242. [Google Scholar] [CrossRef] [Green Version]
- Wong, W.C.W.; Wong, E.L.Y.; Li, H.; You, J.H.; Ho, S.; Woo, J.; Hui, E. Isoflavones in Treating Watchful Waiting Benign Prostate Hyperplasia: A Double-Blinded, Randomized Controlled Trial. J. Altern. Complementary Med. 2012, 18, 54–60. [Google Scholar] [CrossRef] [Green Version]
- van Die, M.D.; Bone, K.M.; Williams, S.G.; Pirotta, M.V. Soy and Soy Isoflavones in Prostate Cancer: A Systematic Review and Meta-Analysis of Randomized Controlled Trials: Soy and Soy Isoflavones in Prostate Cancer. BJU Int. 2014, 113, E119–E130. [Google Scholar] [CrossRef] [Green Version]
- Duru, K.C.; Mukhlynina, E.A.; Moroz, G.A.; Gette, I.F.; Danilova, I.G.; Kovaleva, E.G. Anti-Diabetic Effect of Isoflavone Rich Kudzu Root Extract in Experimentally Induced Diabetic Rats. J. Funct. Foods 2020, 68, 103922. [Google Scholar] [CrossRef]
- Shang, X.; Dou, Y.; Zhang, Y.; Tan, J.-N.; Liu, X.; Zhang, Z. Tailor-Made Natural Deep Eutectic Solvents for Green Extraction of Isoflavones from Chickpea (Cicer Arietinum L.) Sprouts. Ind. Crop. Prod. 2019, 140, 111724. [Google Scholar] [CrossRef]
- Bespalov, V.G.; Alexandrov, V.A.; Semenov, A.L.; Tochilnikov, G.V.; Ermakova, E.D.; Zmitrichenko, Y.G.; Vasilyeva, I.N.; Ivantsov, A.O.; Zhilinskaya, N.T. Old Rats Are More Susceptible to Induction of Benign Prostatic Hyperplasia (BPH) at Comparative to Young Mature. CAS 2021, 14, 124–132. [Google Scholar] [CrossRef]
- de Amorim Ribeiro, I.C.; da Costa, C.A.S.; da Silva, V.A.P.; Côrrea, L.B.N.S.; Boaventura, G.T.; Chagas, M.A. Flaxseed Reduces Epithelial Proliferation but Does Not Affect Basal Cells in Induced Benign Prostatic Hyperplasia in Rats. Eur. J. Nutr. 2017, 56, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Panchenko, A.V.; Tyndyk, M.L.; Fedoros, E.I.; Maydin, M.A.; Semenov, A.L.; Gubareva, E.A.; Golubev, A.G.; Anisimov, V.N. Comparative Analysis of Experimental Data on the Effects of Different Polyphenols on Lifespan and Aging. Adv. Gerontol. 2019, 9, 396–401. [Google Scholar] [CrossRef]


{kind=link}
| Group | Mean Animal Weight (g) | Organ/Body Weight Ratio at the End of the Study (mg/100 g) | ||||||
|---|---|---|---|---|---|---|---|---|
| at the Start of the Study | at the End of the Study | Whole Prostate | Dorsolateral Lobe | Ventral Lobes | Anterior Lobes | Liver | Heart | |
| Control | 278 ± 6 | 375 ± 9 | 180 ± 18 | 90 ± 7 | 66 ± 10 | 24 ± 3 | 3833 ± 253 | 331 ± 15 |
| BPH | 280 ± 7 | 320 ± 9 ** | 508 ± 34 *** | 220 ± 21 *** | 212 ± 18 *** | 76 ± 6 *** | 3222 ± 265 | 400 ± 16 |
| BPH + finasteride | 280 ± 7 | 346 ± 6 *, # | 382 ± 14 ***, ### | 180 ± 9 ***, ## | 148 ± 8 ***, ### | 54 ± 3 ***, ## | 3348 ± 196 | 375 ± 10 |
| BPH + isoflavones | 280 ± 10 | 317 ± 13 ** | 459 ± 34 ***, # | 221 ± 15 *** | 166 ± 19 ***, ## | 75 ± 5 *** | 3435 ± 225 | 409 ± 39 |
| BPH + astaxanthin | 280 ± 5 | 339 ± 11 * | 443 ± 14 ***, ## | 190 ± 5 ***, # | 178 ± 12 ***, ## | 75 ± 4 *** | 3268 ± 229 | 387 ± 18 |
| Group | Testosterone, NG/G Protein | Ca µmol/g Protein | Mg µmol/g Protein | Ca/Mg |
|---|---|---|---|---|
| Control | 4.6 ± 0.9 | 49.0 ± 16.5 | 62.2 ± 11.8 | 0.8 ± 0.2 |
| BPH | 5.7 ± 1.7 | 62.6 ± 15.9 | 53.1 ± 15.7 | 1.3 ± 0.3 *** |
| BPH + finasteride | 5.0 ± 2.8 | 56.9 ± 8.8 | 59.1 ± 8.6 | 1.0 ± 0.1 ##, * |
| BPH + isoflavones | 4.7 ± 2.1 | 51.3 ± 2.3 | 63.4 ± 9.4 | 0.8 ± 0.1 ## |
| BPH + astaxanthin | 5.8 ± 1.9 | 52.6 ± 3.9 | 64.8 ± 5.8 | 0.8 ± 0.1 ## |
| Group | White Blood Cells, 109/L | Lymphocytes, 109/L | Monocytes, 109/L | Granulocytes, 109/L | Red Blood Cells, 1012/L | Hemoglobin, g/L | Hematocrit, % | Platelets, 109/L |
| Control | 16.2 ± 1.9 | 12.6 ± 1.5 | 0.4 ± 0.1 | 3.2 ± 0.3 | 10.1 ± 0.2 | 163 ± 3 | 50 ± 1 | 954 ± 62 |
| BPH | 9.9 ± 0.9 * | 5.9 ± 0.6 * | 0.3 ± 0.0 | 4.0 ± 0.6 | 11.3 ± 0.2 * | 182 ± 4 * | 59 ± 1 * | 1054 ± 74 |
| BPH + finasteride | 9.8 ± 0.4 * | 5.4 ± 0.4 * | 0.3 ± 0.0 | 4.3 ± 0.5 | 11.6 ± 0.2 * | 192 ± 4 * | 62 ± 1 * | 950 ± 128 |
| BPH + isoflavones | 14.2 ± 1.1 # | 9.4 ± 0.8 # | 0.5 ± 0.1 | 4.5 ± 0.4 * | 11.9 ± 0.2 * | 197 ± 2 * | 63 ± 1 * | 951 ± 143 |
| BPH + astaxanthin | 12.2 ± 1.2 | 7.7 ± 1 * | 0.4 ± 0 | 4.3 ± 0.2 * | 11.5 ± 0.1 * | 190 ± 3 * | 61 ± 1 * | 1000 ± 31 |
| Group | Glucose (mmol/L) | Triglycerides (mmol/L) | Ca (mmol/L) | Mg (mmol/L) | Ca/Mg | SOD, Relative Activity | Testosterone, ng/mL |
|---|---|---|---|---|---|---|---|
| Control | 9.2 ± 1.4 | 2.9 ± 0.8 | 2.8 ± 0.3 | 0.9 ± 0.1 | 3.2 ± 0.2 | 100 ± 8% | 3.4 ± 1.9 |
| BPH | 7.2 ± 1.4 ** | 2.4 ± 1.2 | 3.4 ± 0.6 * | 1.5 ± 0.8 * | 2.7 ± 0.7 | 67 ± 7% * | 13.5 ± 4.2 *** |
| BPH + finasteride | 7.0 ± 0.3 ** | 2.2 ± 1.5 | 2.8 ± 0.3 # | 0.9 ± 0.1 # | 3.0 ± 0.2 | 98 ± 6% # | 18.2 ± 3.0 ***, # |
| BPH + isoflavones | 6.8 ± 0.9 ** | 1.7 ± 1.4 | 2.9 ± 0.2 # | 1.0 ± 0.1 # | 2.9 ± 0.1 | 88 ± 5% | 14.2 ± 2.3 *** |
| BPH + astaxanthin | 6.5 ± 1.2 ** | 1.8 ± 1.3 | 2.8 ± 0.3 # | 1.0 ± 0.1 # | 2.9 ± 0.2 | 111 ± 13% # | 17.9 ± 3.3 ***, # |
| Name | Chemical Structure | Chemical Name |
|---|---|---|
| Kudzu root NADES extract | ||
| Daidzein |  | 7-Hydroxy-3-(4-hydroxyphenyl)-4H-chromen-4-one |
| Genistein |  | 5,7-Dihydroxy-3-(4-hydroxyphenyl)-4H-chromen-4-one |
| Puerarin |  | 8-(β-D-Glucopyranosyl)-7-hydroxy-3-(4-hydroxyphenyl)-4H-chromen-4-one |
| Formononetin |  | 7-Hydroxy-3-(4-methoxyphenyl)-4H-chromen-4-one |
| Biochanin A |  | 5,7-Dihydroxy-3-(4-methoxyphenyl)-4H-chromen-4-one |
| Phaffia rhodozyma extract | ||
| Astaxanthin |  | (3S,3′S)-3,3′-Dihydroxy-β,β-carotene-4,4′-dione |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, A.L.; Gubareva, E.A.; Ermakova, E.D.; Dorofeeva, A.A.; Tumanyan, I.A.; Radetskaya, E.A.; Yurova, M.N.; Aboushanab, S.A.; Kanwugu, O.N.; Fedoros, E.I.; et al. Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance. Plants 2021, 10, 2735. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122735
Semenov AL, Gubareva EA, Ermakova ED, Dorofeeva AA, Tumanyan IA, Radetskaya EA, Yurova MN, Aboushanab SA, Kanwugu ON, Fedoros EI, et al. Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance. Plants. 2021; 10(12):2735. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122735
Chicago/Turabian StyleSemenov, Alexander L., Ekaterina A. Gubareva, Elena D. Ermakova, Anastasia A. Dorofeeva, Irina A. Tumanyan, Ekaterina A. Radetskaya, Maria N. Yurova, Saied A. Aboushanab, Osman N. Kanwugu, Elena I. Fedoros, and et al. 2021. "Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance" Plants 10, no. 12: 2735. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122735