
Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Growth and Physiological Parameters
2.2. Carbohydrates Content
2.3. Nitrogen Containing Compounds
2.4. Sap Osmolality and Contribution of Ions and Metabolites to the Osmotic Adjustment
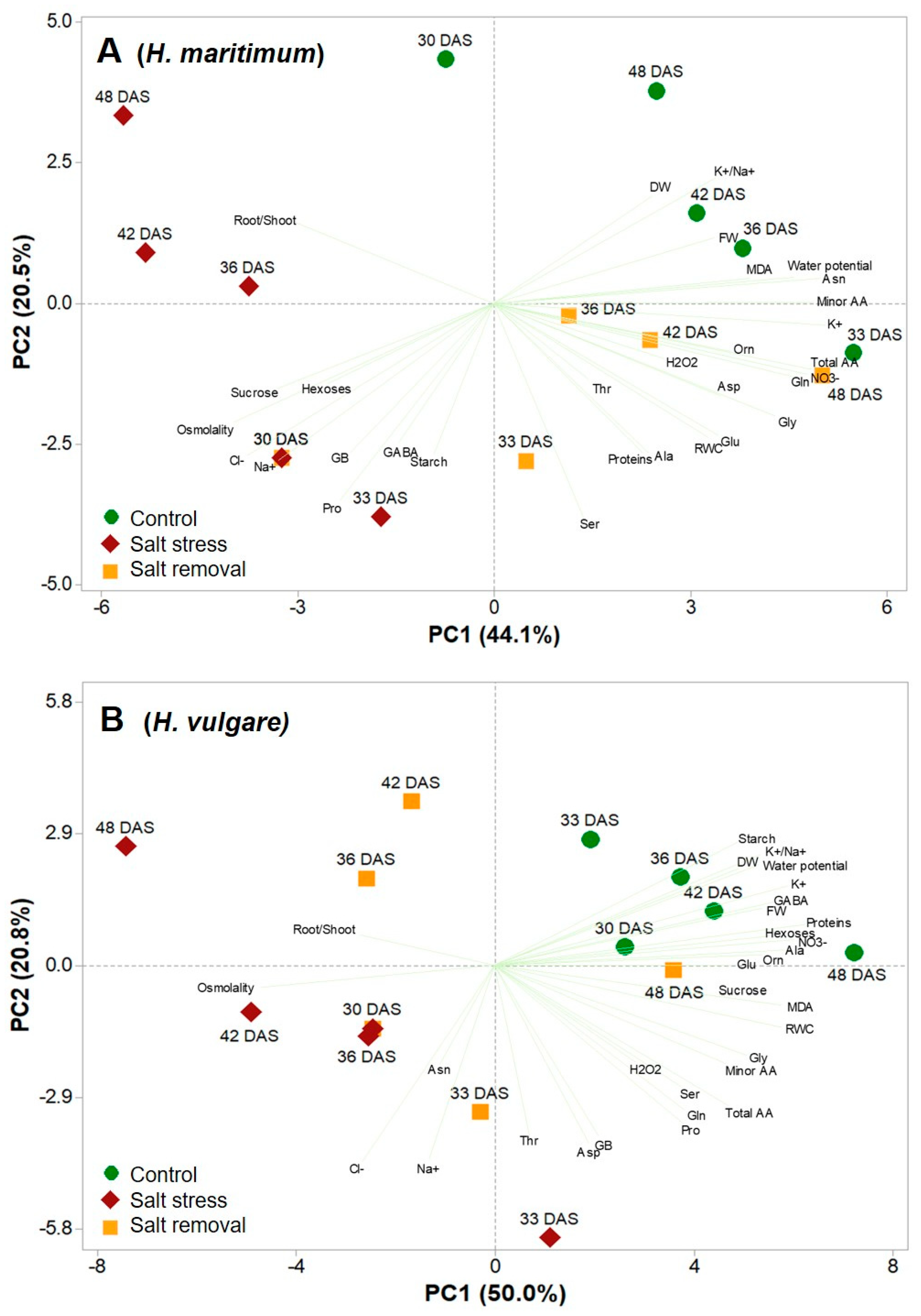
2.5. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Measurements of FW and DW, Water Relations and Osmolality
4.3. Ions, Hydrogen Peroxide, Malondialdehyde and Metabolites Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maggio, A.; De Pascale, S.; Fagnano, M.; Barbieri, G. Saline agriculture in Mediterranean environments. Ital. J. Agron. 2011, 6, e7. [Google Scholar] [CrossRef]
- Carillo, P.; Grazia, M.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity Stress and Salt Tolerance. In Abiotic Stress in Plants—Mechanisms and Adaptations; IntechOpen: Rijeka, Croatia, 2011; p. 18396. [Google Scholar]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessini, K.; Jeddi, K.; Siddique, K.H.M.; Cruz, C. Drought and salinity: A comparison of their effects on the ammoni-um-preferring species Spartina alterniflora. Physiol. Plant. 2020. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2016, 159, 290–312. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Bhowmik, P.C.; Hossain, A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. BioMed Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Hessini, K.; Ferchichi, S.; Ben Youssef, S.; Werner, K.H.; Cruz, C.; Gandour, M. How Does Salinity Duration Affect Growth and Productivity of Cultivated Barley? Agron. J. 2015, 107, 174–180. [Google Scholar] [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Munns, R. Salinity Stress: Physiological Constraints and Adaptive Mechanisms; CABI: Egham, UK, 2012. [Google Scholar]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Neumann, P.M. Inhibition of root growth by salinity stress: Toxicity or an adaptive biophysical response? In Structure and Function of Roots, Proceedings of the Fourth International Symposium on Structure and Function of Roots, Stará Lesná, Slovakia, 20–26 June 1993; Baluška, F., Čiamporová, M., Gašparíková, O., Barlow, P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 299–304. [Google Scholar]
- Cramer, G.R.; Läuchli, A.; Polito, V.S. Displacement of Ca2+ by Na+ from the Plasmalemma of Root Cells: A Primary Response to Salt Stress? Plant Physiol. 1985, 79, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Hajibagheri, M.A. Salinity tolerance in Hordeum vulgare: Ion concentrations in root cells of cultivars differing in salt tolerance**. Plant Soil 2001, 231, 1–9. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Ciarmiello, L.F.; Woodrow, P.; Piccirillo, P.; De Luca, A.; Carillo, P. Transcription Factors and Environmental Stresses in Plants. In Emerging Technologies and Management of Crop Stress Tolerance; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 1, pp. 57–78. [Google Scholar]
- Woodrow, P.; Pontecorvo, G.; Fantaccione, S.; Fuggi, A.; Kafantaris, I.; Parisi, D.; Carillo, P. Polymorphism of a new Ty1-copia retrotransposon in durum wheat under salt and light stresses. Theor. Appl. Genet. 2010, 121, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Pontecorvo, G.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Ttd1a promoter is involved in DNA–protein binding by salt and light stresses. Mol. Biol. Rep. 2011, 38, 3787–3794. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Woodrow, P.; Raimondi, G.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Colla, G.; Mori, M.; Giordano, M.; Giordano, M.; et al. Omeprazole Promotes Chloride Exclusion and Induces Salt Tolerance in Greenhouse Basil. Agronomy 2019, 9, 355. [Google Scholar] [CrossRef] [Green Version]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puniran-Hartley, N.; Hartley, J.; Shabala, L.; Shabala, S. Salinity-induced accumulation of organic osmolytes in barley and wheat leaves correlates with increased oxidative stress tolerance: In planta evidence for cross-tolerance. Plant Physiol. Biochem. 2014, 83, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: Accumulation of proline and glycine betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef]
- Mansour, M.M. Nitrogen Containing Compounds and Adaptation of Plants to Salinity Stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Suprasanna, P.; Nikalje, G.C.; Rai, A.N. Osmolyte Accumulation and Implications in Plant Abiotic Stress Tolerance. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Iqbal, N., Nazar, R.A., Khan, N., Eds.; Springer: New Delhi, India, 2016; pp. 1–12. [Google Scholar]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Al Hassan, M.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of salinity and drought on growth, ionic relations, compatible solutes and activation of antioxidant systems in oleander (Nerium oleander L.). PLoS ONE 2017, 12, e0185017. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Nieman, R.H.; Jung, G.A. Physiology of Plant Tolerance to Salinity. In Soil Erosion and Conservation in the Tropics; John and Wiley and Sons: Hoboken, NJ, USA, 2015; pp. 277–299. [Google Scholar]
- Stoskopf, N.C. Cereal Grain Crops; Reston Publishing Company: Reston, VA, USA, 1985. [Google Scholar]
- Nongpiur, R.C.; Singla-Pareek, S.L.; Pareek, A. Genomics Approaches for Improving Salinity Stress Tolerance in Crop Plants. Curr. Genom. 2016, 17, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vij, S.; Tyagi, A.K. Emerging trends in the functional genomics of the abiotic stress response in crop plants. Plant Biotechnol. J. 2007, 5, 361–380. [Google Scholar] [CrossRef]
- Budak, H.; Hussain, B.; Khan, Z.; Ozturk, N.Z.; Ullah, N. From Genetics to Functional Genomics: Improvement in Drought Signaling and Tolerance in Wheat. Front. Plant Sci. 2015, 6, 1012. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2017, 217, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, S.F.; Martins, D.; Agacka-Mołdoch, M.; Czubacka, A.; Araújo, S.D.S. Strategies to Alleviate Salinity Stress in Plants. In Salinity Responses and Tolerance in Plants; Springer Nature: Berlin/Heidelberg, Germany, 2018; Volume 1, pp. 307–337. [Google Scholar]
- Cuevas, J.; Daliakopoulos, I.N.; Del Moral, F.; Hueso, J.J.; Tsanis, I. A Review of Soil-Improving Cropping Systems for Soil Salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Raimondi, G.; Lucini, L.; Carillo, P.; Kyriacou, M.C.; Colla, G.; Cirillo, V.; Pannico, A.; El-Nakhel, C.; De De Pascale, S. Physiological and Metabolic Responses Triggered by Omeprazole Improve Tomato Plant Tolerance to NaCl Stress. Front. Plant Sci. 2018, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro- and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Letey, J.; Hoffman, G.; Hopmans, J.W.; Grattan, S.; Suarez, D.L.; Corwin, D.L.; Oster, J.; Wu, L.; Amrhein, C. Evaluation of soil salinity leaching requirement guidelines. Agric. Water Manag. 2011, 98, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Evolkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef]
- Yousefi, A.R.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and Seedling Growth Responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-Induced Drought Stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef] [PubMed]
- Garthwaite, A.J.; Von Bothmer, R.; Colmer, T.D. Salt tolerance in wild Hordeum species is associated with restricted entry of Na+ and Cl− into the shoots. J. Exp. Bot. 2005, 56, 2365–2378. [Google Scholar] [CrossRef]
- Gorham, J.; Jones, R.G.W.; Bristol, A. Partial characterization of the trait for enhanced K+−Na+ discrimination in the D genome of wheat. Planta 1990, 180, 590–597. [Google Scholar] [CrossRef] [PubMed]
- EAbdElGawad, H.; Ezinta, G.; Hegab, M.M.; Epandey, R.; Easard, H.; Eabuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Martínez, J.P.; Gandour, M.; Albouchi, A.; Soltani, A.; Abdelly, C. Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water-use efficiency in Spartina alterniflora. Environ. Exp. Bot. 2009, 67, 312–319. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- James, R.A.; Von Caemmerer, S.; Condon, A.G.; Zwart, A.B.; Munns, R. Genetic variation in tolerance to the osmotic stress componentof salinity stress in durum wheat. Funct. Plant Biol. 2008, 35, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose. Front. Plant Sci. 2017, 7, 2035. [Google Scholar] [CrossRef] [Green Version]
- Rahnama, A.; Munns, R.; Poustini, K.; Watt, M. A screening method to identify genetic variation in root growth response to a salinity gradient. J. Exp. Bot. 2011, 62, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Keys, A.J. The re-assimilation of ammonia produced by photorespiration and the nitrogen economy of C3 higher plants. Photosynth. Res. 2006, 87, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Abogadallah, G.M.; Serag, M.S.; Quick, W.P. Fine and coarse regulation of reactive oxygen species in the salt tolerant mutants of barnyard grass and their wild-type parents under salt stress. Physiol. Plant. 2010, 138, 60–73. [Google Scholar] [CrossRef]
- Ma, J.; Cirillo, V.; Zhang, D.; Zhang, D.; Wang, L.; Xiao, X.; Yao, Y. Regulation of Ammonium Cellular Levels is An Important Adaptive Trait for the Euhalophytic Behavior of Salicornia europaea. Plants 2020, 9, 257. [Google Scholar] [CrossRef] [Green Version]
- Rozema, J.; Bijwaard, P.; Prast, G.; Broekman, R. Ecophysiological adaptations of coastal halophytes from foredunes and salt marshes. Vegetatio 1985, 62, 499–521. [Google Scholar] [CrossRef]
- MacNeill, G.J.; Mehrpouyan, S.A.A.; Minow, M.A.; Patterson, J.; Tetlow, I.J.; Emes, M.J. Starch as a source, starch as a sink: The bifunctional role of starch in carbon allocation. J. Exp. Bot. 2017, 68, 4433–4453. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Yousfi, S.; Rabhi, M.; Hessini, K.; Abdelly, C.; Gharsalli, M. Differences in efficient metabolite management and nutrient metabolic regulation between wild and cultivated barley grown at high salinity. Plant Biol. 2009, 12, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, D.; Jin, T.; Chang, Q.; Yin, D.; Xu, S.; Liu, B.; Liu, L. The Vacuolar Na+/H+ Antiporter Gene SsNHX1 from the Halophyte Salsola soda Confers Salt Tolerance in Transgenic Alfalfa (Medicago sativa L.). Plant Mol. Biol. Rep. 2010, 29, 278–290. [Google Scholar] [CrossRef]
- Garbarino, J.; Dupont, F.M. NaCl Induces a Na+/H+ Antiport in Tonoplast Vesicles from Barley Roots. Plant Physiol. 1988, 86, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.; Hessini, K.; Esmat, F. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef]
- Rubio, F.; Gassmann, W.; Schroeder, J.I. Sodium-Driven Potassium Uptake by the Plant Potassium Transporter HKT1 and Mutations Conferring Salt Tolerance. Science 1995, 270, 1660–1663. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Ligaba, A.; Katsuhara, M. Insights into the salt tolerance mechanism in barley (Hordeum vulgare) from comparisons of cultivars that differ in salt sensitivity. J. Plant Res. 2009, 123, 105–118. [Google Scholar] [CrossRef]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S. Chloroplast function and ion regulation in plants growing on saline soils: Lessons from halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, T.C.; Xu, L. Sensitivity of growth of roots versus leaves to water stress: Biophysical analysis and relation to water transport. J. Exp. Bot. 2000, 51, 1595–1616. [Google Scholar] [CrossRef] [PubMed]
- De Lacerda, C.F.; Cambraia, J.; Oliva, M.A.; Ruiz, H.A. Changes in growth and in solute concentrations in sorghum leaves and roots during salt stress recovery. Environ. Exp. Bot. 2005, 54, 69–76. [Google Scholar] [CrossRef]
- Al-Hakimi, A.; Hamada, A. Counteraction of Salinity Stress on Wheat Plants by Grain Soaking in Ascorbic Acid, Thiamin or Sodium Salicylate. Biol. Plant. 2001, 44, 253–261. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, E.; Fottrell, P.F. Subcellular localization of enzymes involved in the assimilation of Ammonia by soybean root nodules. Phytochemistry 1974, 13, 2647–2652. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Krishnan, S.R.; Pandian, S.; Mareeswaran, N.; Aruni, W.; Pandian, S.K.; Ramesh, M. Global analysis of threonine metabolism genes unravel key players in rice to improve the abiotic stress tolerance. Sci. Rep. 2018, 8, 9270. [Google Scholar] [CrossRef] [PubMed]
- Kalamaki, M.S.; Merkouropoulos, G.; Kanellis, A.K. Can ornithine accumulation modulate abiotic stress tolerance in Arabidopsis? Plant Signal. Behav. 2009, 4, 1099–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejri, M.; Siddique, K.H.M.; Saif, T.; Abdelly, C.; Hessini, K. Comparative effect of drought duration on growth, photosynthesis, water relations, and solute accumulation in wild and cultivated barley species. J. Plant Nutr. Soil Sci. 2016, 179, 327–335. [Google Scholar] [CrossRef]
- Rabhi, M.; Hafsi, C.; Lakhdar, A.; Barhoumi, Z.; Abdelly, C.; Smaoui, A. Evaluation of the capacity of three halophytes to desalinize their rhizosphere as grown on saline soils under nonleaching conditions. Afr. J. Ecol. 2009, 47, 463–468. [Google Scholar] [CrossRef]
- Lombardi, T.; Lupi, B. Salt-tolerance in wild Hordeum (Poaceae) species: Differences between H. maritimum With. and H. hystrix Roth. Atti. Soc. Tosc. Sci. Nat. 2006, 113, 31–35. [Google Scholar]
- Hafsi, C.; Lakhdhar, A.; Rabhi, M.; Debez, A.; Abdelly, C.; Ouerghi, Z. Interactive effects of salinity and potassium availability on growth, water status, and ionic composition of Hordeum maritimum. J. Plant Nutr. Soil Sci. 2007, 170, 469–473. [Google Scholar] [CrossRef]
- Hammami, H.; Gomes, T.; Abdelly, C.; Mahmoud, O.M.-B.; Baptista, P.; Martins, F. Impact of a natural soil salinity gradient on fungal endophytes in wild barley (Hordeum maritimum With.). World J. Microbiol. Biotechnol. 2016, 32, 184. [Google Scholar] [CrossRef] [Green Version]
- Abdelly, C.; Lachaal, M.; Grignon, C.; Soltani, A.; Hajji, M. Association épisodique d’halophytes strictes et de glycophytes dans un écosystème hydromorphe salé en zone semi-aride. Agronomie 1995, 15, 557–568. [Google Scholar] [CrossRef]
- Rad, S.V.; Valadabadi, S.A.R.; Pouryousef, M.; Saifzadeh, S.; Zakrin, H.R.; Mastinu, A. Quantitative and Qualitative Evaluation of Sorghum bicolor L. under Intercropping with Legumes and Different Weed Control Methods. Horticulturae 2020, 6, 78. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Shamsi, N.; Mosa, K. Effect of maternal habitat, temperature and light on germination and salt tolerance of Suaeda vermiculata, a habitat-indifferent halophyte of arid Arabian deserts. Seed Sci. Res. 2018, 28, 140–147. [Google Scholar] [CrossRef]
- Shah, S.Z.; Rasheed, A.; Gul, B.; Khan, M.A.; Nielsen, B.L.; Hameed, A. Maternal salinity improves yield, size and stress tolerance of Suaeda fruticosa seeds. J. Arid. Land 2020, 12, 283–293. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Bovill, J.; Afzal, I.; Hayes, J.E.; Roy, S.J.; Tester, M.; Collins, N.C. HVP10 encoding V-PPase is a prime candidate for the barley HvNax3 sodium exclusion gene: Evidence from fine mapping and expression analysis. Planta 2013, 237, 1111–1122. [Google Scholar] [CrossRef]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water Relations in Winter Wheat as Drought Resistance Indicators. Crop. Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Cuin, T.A.; Tian, Y.; Betts, S.A.; Chalmandrier, R.; Shabala, S. Ionic relations and osmotic adjustment in durum and bread wheat under saline conditions. Funct. Plant Biol. 2009, 36, 1110–1119. [Google Scholar] [CrossRef]
- Carillo, P.; Parisi, D.; Woodrow, P.; Pontecorvo, G.; Massaro, G.; Annunziata, M.G.; Fuggi, A.; Sulpice, R. Salt-induced accumulation of glycine betaine is inhibited by high light in durum wheat. Funct. Plant Biol. 2011, 38, 139–150. [Google Scholar] [CrossRef]
- Baptista, P.; Martins, A.; Pais, M.S.; Tavares, R.M.; Lino-Neto, T. Involvement of reactive oxygen species during early stages of ectomycorrhiza establishment between Castanea sativa and Pisolithus tinctorius. Mycorrhiza 2007, 17, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havaux, M.; Eymery, F.; Porfirova, S.; Rey, P.; Dörmann, P. Vitamin E Protects against Photoinhibition and Photooxidative Stress in Arabidopsis thaliana. Plant Cell 2005, 17, 3451–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Cacace, D.; De Pascale, S.; Rapacciuolo, M.; Fuggi, A. Organic vs. traditional potato powder. Food Chem. 2012, 133, 1264–1273. [Google Scholar] [CrossRef]
- Ciarmiello, L.F.; Piccirillo, P.; Carillo, P.; De Luca, A.; Woodrow, P. Determination of the genetic relatedness of fig (Ficus carica L.) accessions using RAPD fingerprint and their agro-morphological characterization. S. Afr. J. Bot. 2015, 97, 40–47. [Google Scholar] [CrossRef]
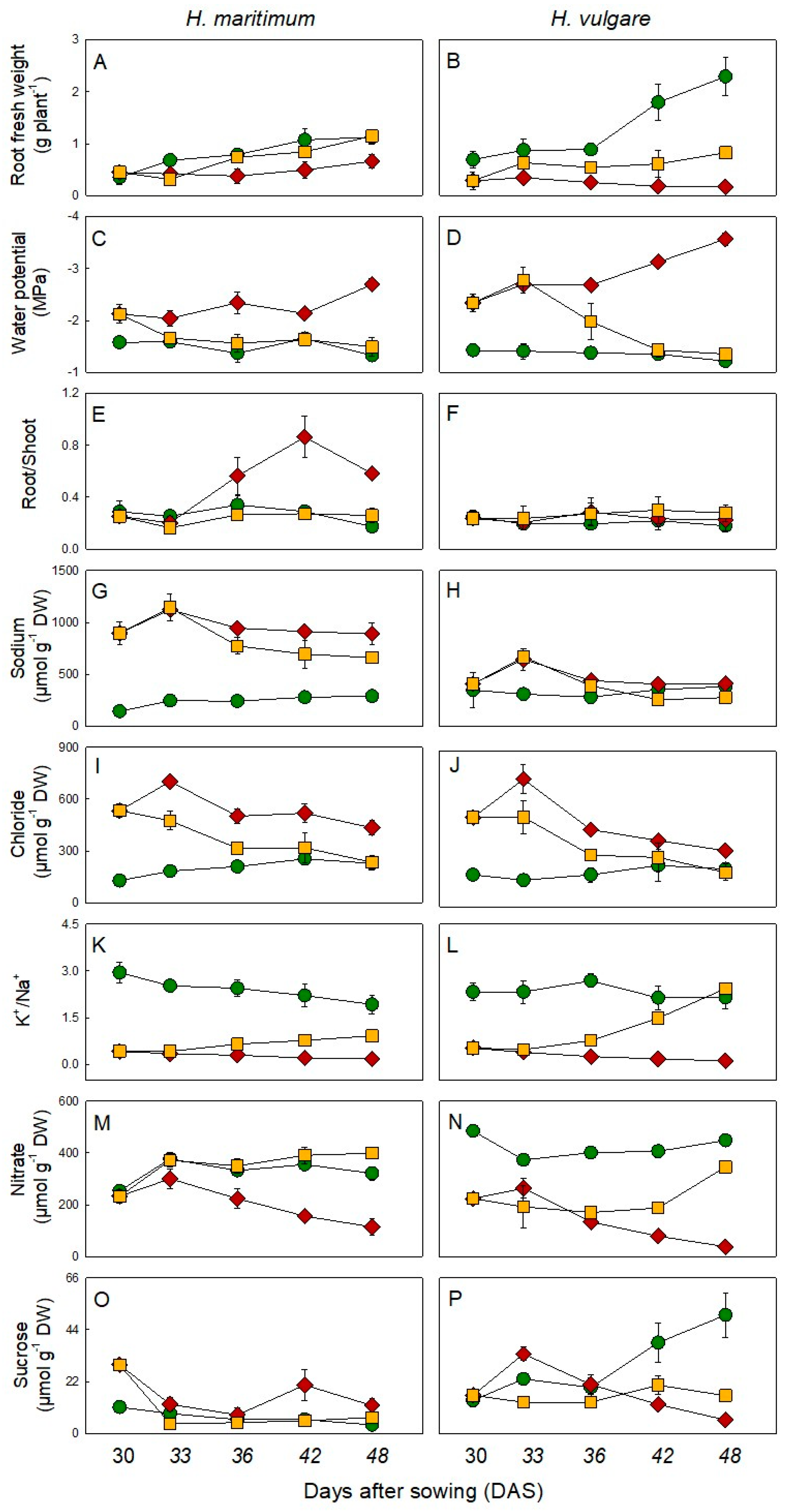
 ), salt stress (200mM NaCl,
), salt stress (200mM NaCl,  ) and salt removal treatments (
) and salt removal treatments (  ). Salt was gradually added to salt treated plants starting from 15 days after sowing (DAS). Salt removal treatment started from 30 DAS. Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Values are mean ± s.d. (n = 3).
), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salt treated plants starting from 15 days after sowing (DAS). Salt removal treatment started from 30 DAS. Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Values are mean ± s.d. (n = 3).
). Salt was gradually added to salt treated plants starting from 15 days after sowing (DAS). Salt removal treatment started from 30 DAS. Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Values are mean ± s.d. (n = 3).
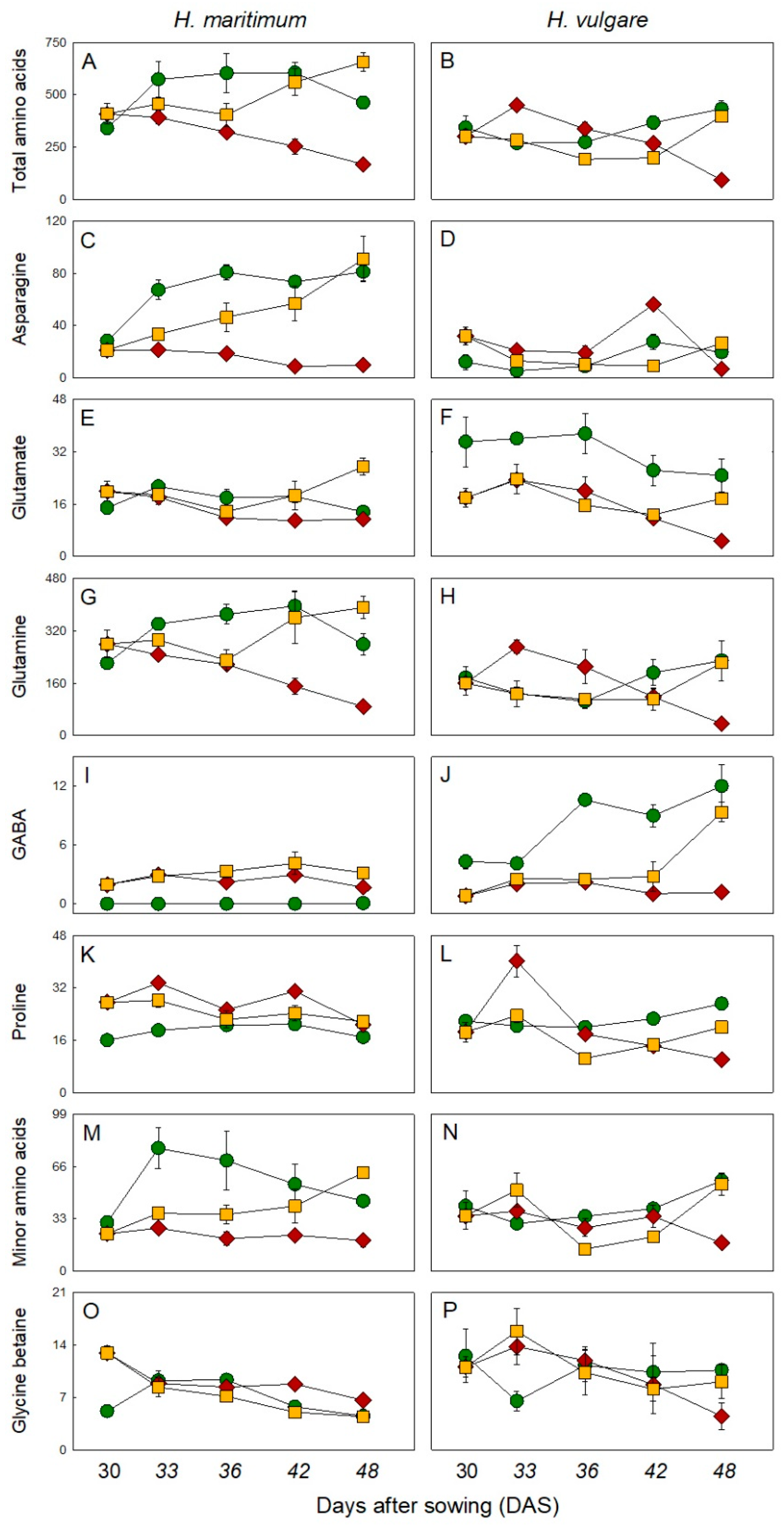
), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salt treated plants starting from 15 days after sowing (DAS). Salt removal treatment started from 30 DAS. Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Values are mean ± s.d. (n = 3). ), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salinity treatments starting from 15 days after sowing (DAS). Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Metabolites were expressed as µmol g–1 DW. Values are mean ± s.d. (n = 3).
), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salinity treatments starting from 15 days after sowing (DAS). Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Metabolites were expressed as µmol g–1 DW. Values are mean ± s.d. (n = 3).
), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salinity treatments starting from 15 days after sowing (DAS). Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Metabolites were expressed as µmol g–1 DW. Values are mean ± s.d. (n = 3).
), salt stress (200mM NaCl, ) and salt removal treatments ( ). Salt was gradually added to salinity treatments starting from 15 days after sowing (DAS). Harvests were conducted at 30, 33, 36, 42 and 48 DAS. Metabolites were expressed as µmol g–1 DW. Values are mean ± s.d. (n = 3).


{kind=link}
{kind=link}
{kind=link}
| A. Hordeum maritimum | 30 Days | 36 Days | 48 Days | |||||
| C | S/R | C | S | R | C | S | R | |
| Osmolarity (mOsmol/kg) | 330 | 617 | 307 | 553 | 460 | 315 | 645 | 453 |
| Chloride | 5.4 | 10.2 | 7.1 | 10.4 | 6.3 | 8.2 | 8.7 | 4.8 |
| Nitrate | 10.7 | 4.5 | 11.3 | 4.7 | 7.0 | 11.5 | 2.3 | 8.2 |
| Potassium | 17.8 | 7.2 | 20.1 | 5.8 | 10.1 | 20.1 | 3.1 | 12.4 |
| Sodium | 6.0 | 17.1 | 8.2 | 19.7 | 15.5 | 10.4 | 17.7 | 13.5 |
| Measured ions | 39.9 | 39.0 | 46.6 | 40.6 | 38.9 | 50.3 | 31.8 | 38.9 |
| Hexoses | 4.3 | 3.0 | 1.4 | 1.1 | 0.5 | 1.2 | 1.1 | 1.4 |
| Sucrose | 0.5 | 0.6 | 0.2 | 0.2 | 0.1 | 0.1 | 0.2 | 0.1 |
| Sum of Soluble Sugars | 4.7 | 3.6 | 1.6 | 1.3 | 0.6 | 1.4 | 1.3 | 0.6 |
| Total Amino Acids | 14.4 | 7.8 | 20.5 | 6.7 | 8.1 | 16.6 | 3.4 | 13.4 |
| Asn | 1.2 | 0.4 | 2.7 | 0.4 | 0.9 | 2.9 | 0.2 | 1.9 |
| GABA | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.0 | 0.1 |
| Gln | 9.3 | 5.3 | 12.6 | 4.5 | 4.6 | 10.0 | 1.8 | 8.0 |
| Glu | 0.6 | 0.4 | 0.6 | 0.2 | 0.3 | 0.5 | 0.2 | 0.6 |
| Pro | 0.7 | 0.5 | 0.7 | 0.5 | 0.4 | 0.6 | 0.4 | 0.4 |
| Minor AA | 1.3 | 0.5 | 2.4 | 0.4 | 0.7 | 1.6 | 0.4 | 1.3 |
| Glycine betaine | 0.2 | 0.2 | 0.3 | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 |
| Measured Organic Osmolytes | 19.3 | 11.6 | 22.4 | 8.2 | 8.9 | 18.1 | 4.8 | 15.1 |
| Other Metabolites | 40.7 | 49.4 | 31.0 | 51.2 | 52.2 | 31.6 | 63.4 | 46.0 |
| B. Hordeum vulgare | 30 Days | 36 Days | 48 Days | |||||
| C | S/R | C | S | R | C | S | R | |
| Osmolarity (mOsmol/kg) | 354 | 370 | 338 | 393 | 304 | 295 | 475 | 292 |
| Chloride | 4.0 | 15.0 | 4.7 | 15.8 | 16.7 | 4.9 | 13.6 | 6.0 |
| Nitrate | 10.5 | 6.5 | 10.3 | 4.7 | 9.5 | 10.0 | 1.6 | 10.4 |
| Potassium | 17.6 | 6.2 | 19.3 | 3.9 | 16.6 | 18.4 | 1.9 | 20.3 |
| Sodium | 7.5 | 11.8 | 7.2 | 15.6 | 21.7 | 8.6 | 17.3 | 8.3 |
| Measured ions | 39.5 | 39.5 | 41.4 | 40.0 | 64.5 | 41.8 | 34.5 | 45.0 |
| Hexoses | 2.3 | 1.3 | 4.9 | 5.4 | 5.8 | 7.6 | 1.8 | 6.1 |
| Sucrose | 0.3 | 0.5 | 0.5 | 0.7 | 0.7 | 1.1 | 0.2 | 0.5 |
| Sum of Soluble Sugars | 2.7 | 1.8 | 5.4 | 6.1 | 6.6 | 8.7 | 2.00 | 6.6 |
| Total Amino Acids | 7.4 | 8.8 | 7.0 | 11.9 | 10.8 | 9.7 | 3.9 | 12.0 |
| Asn | 0.3 | 0.9 | 0.2 | 0.7 | 0.6 | 0.4 | 0.3 | 0.8 |
| GABA | 0.1 | 0.0 | 0.3 | 0.1 | 0.1 | 0.3 | 0.0 | 0.3 |
| Gln | 3.8 | 4.6 | 2.7 | 7.4 | 6.2 | 5.1 | 1.5 | 6.7 |
| Glu | 0.8 | 0.5 | 1.0 | 0.7 | 0.9 | 0.6 | 0.2 | 0.5 |
| Pro | 0.5 | 0.6 | 0.6 | 0.7 | 0.7 | 0.7 | 0.5 | 0.7 |
| Minor AA | 0.9 | 1.0 | 0.9 | 1.0 | 0.8 | 1.3 | 0.7 | 1.6 |
| Glycine betaine | 0.3 | 0.3 | 0.3 | 0.4 | 0.6 | 0.2 | 0.2 | 0.3 |
| Measured Organic Osmolytes | 10.3 | 10.8 | 12.7 | 18.5 | 17.9 | 18.6 | 6.2 | 18.9 |
| Other Metabolites | 50.1 | 49.6 | 45.9 | 41.5 | 17.6 | 39.6 | 59.4 | 36.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants 2021, 10, 307. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020307
Dell’Aversana E, Hessini K, Ferchichi S, Fusco GM, Woodrow P, Ciarmiello LF, Abdelly C, Carillo P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants. 2021; 10(2):307. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020307
Chicago/Turabian StyleDell’Aversana, Emilia, Kamel Hessini, Selma Ferchichi, Giovanna Marta Fusco, Pasqualina Woodrow, Loredana F. Ciarmiello, Chedly Abdelly, and Petronia Carillo. 2021. "Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes" Plants 10, no. 2: 307. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020307