
Continuous Light Does Not Compromise Growth and Yield in Mini-Cucumber Greenhouse Production with Supplemental LED Light

Abstract
:1. Introduction
2. Materials and Methods
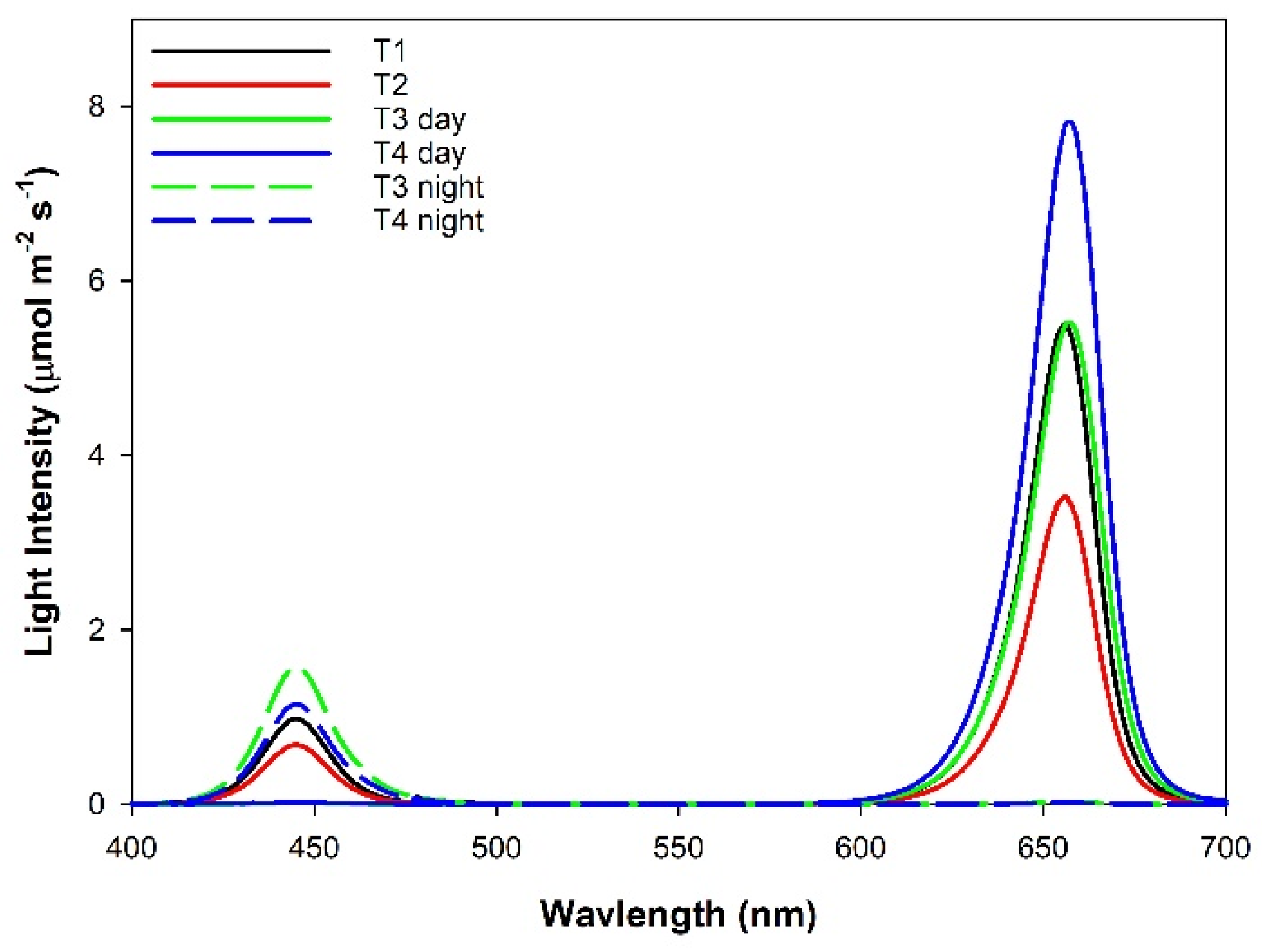
2.1. Plant Material and Experimental Design
2.2. Growth Measurements
2.3. Leaf Gas Exchange: Day and Night Measurements
2.4. Leaf Gas Exchange: Light Response Curves
2.5. Leaf Gas Exchange: CO2 Response Curves
2.6. Chlorophyll Fluorescence Imaging
2.7. Yield
2.8. Economic Analysis
2.9. Statistical Analysis
3. Results
3.1. Plant Growth
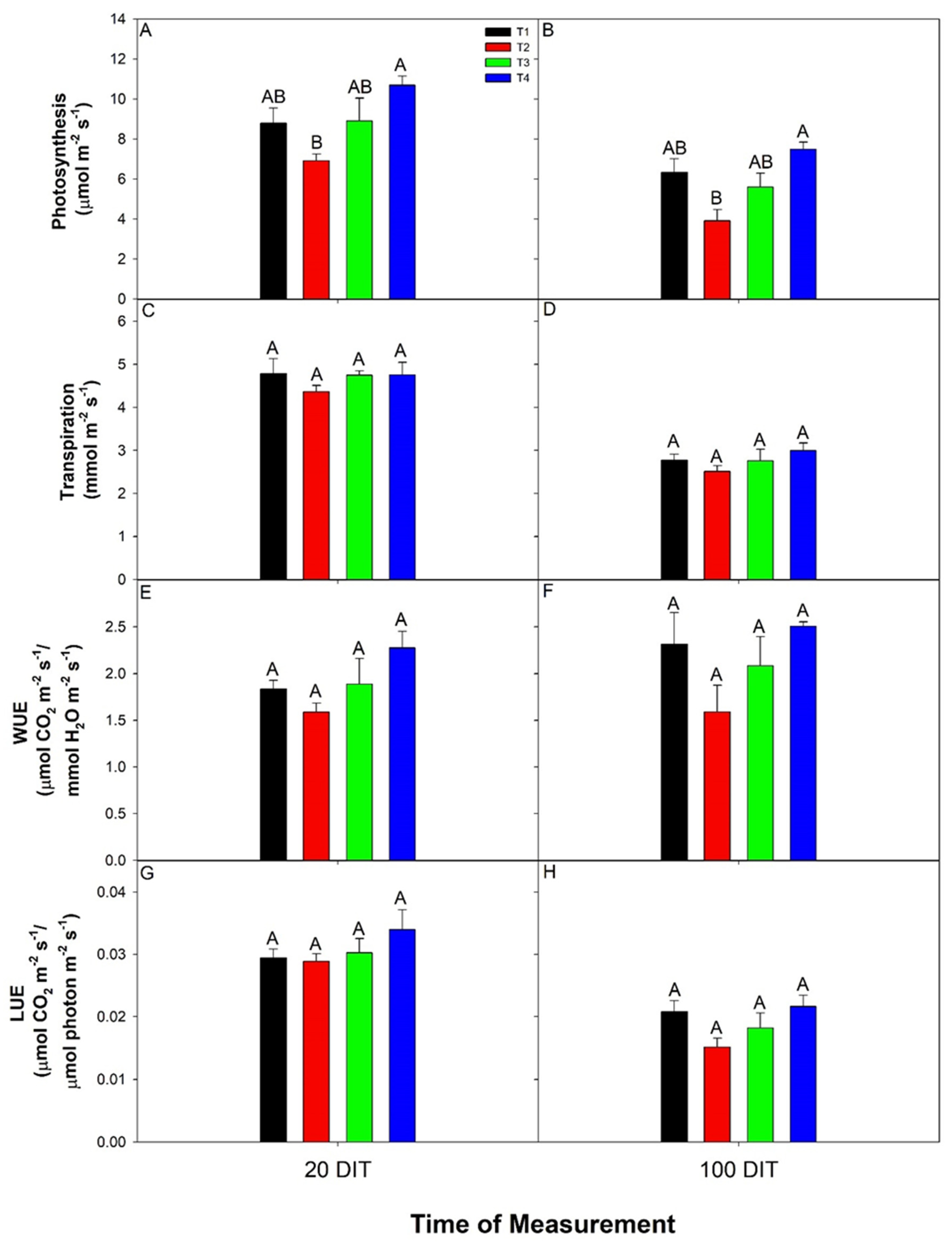
3.2. Day and Night Leaf Gas Exchanges
3.3. Leaf Light and CO2 Response Curves
3.4. Leaf Chlorophyll Fluorescence
3.5. Fruit Production
3.6. Economical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Milenaar, F.F. Plant under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.S.; Kjaer, K.H.; Rosenqvist, E.; Ottosen, C.-O. Continuous light increases growth, daily carbon gain, antioxidants, and alters carbohydrate metabolism in a cultivated and a wild tomato species. Front. Plant Sci. 2015, 6, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtake, N.; Ishikura, M.; Suzuki, H.; Yamori, W.; Goto, E. Continuous Irradiation with Alternating Red and Blue Light Enhances Plant Growth While Keeping Nutritional Quality in Lettuce. HortScience 2018, 53, 1804–1809. [Google Scholar] [CrossRef] [Green Version]
- Lanoue, J.; Zheng, J.; Little, C.; Thibodeau, A.; Grodzinski, B.; Hao, X. Alternating Red and Blue Light-Emitting Diodes Allows for Injury-Free Tomato Production with Continuous Lighting. Front. Plant Sci. 2019, 10, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez-Ramirez, A.; Heuvelink, E.; Van Ieperen, W.; Vreugdenhil, D.; Millenaar, F. Continuous light as a way to increase greenhouse tomato production: Expected challenges. Acta Hortic. 2012, 956, 51–57. [Google Scholar] [CrossRef]
- Demers, D.A.; Gosselin, A. Growing greenhouse tomato and sweet pepper under supplemental lighting: Optimal photoperiod, negative effects of long photoperiod and their causes. Acta Hortic. 2002, 580, 83–88. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; Dünner-Planella, G.; Vreugdenhil, D.; Millenaar, F.F.; Van Ieperen, W. On the induction of injury in tomato under continuous light: Circadian asynchrony as the main triggering factor. Funct. Plant Biol. 2017, 44, 597. [Google Scholar] [CrossRef] [PubMed]
- Velez-Ramirez, A.I.; Carreño-Quintero, N.; Vreugdenhil, D.; Millenaar, F.F.; van Ieperen, W. Sucrose and Starch Content Negatively Correlates with PSII Maximum Quantum Efficiency in Tomato (Solanum lycopersicum) Exposed to Abnormal Light/Dark Cycles and Continuous Light. Plant Cell Physiol. 2017, 58, 1339–1349. [Google Scholar] [CrossRef] [Green Version]
- Velez-Ramirez, A.I.; Van Ieperen, W.; Vreugdenhil, D.; Van Poppel, P.M.J.A.; Heuvelink, E.; Millenaar, F.F. A single locus confers tolerance to continuous light and allows substantial yield increase in tomato. Nat. Commun. 2014, 5, 4549. [Google Scholar] [CrossRef] [Green Version]
- Velez-Ramirez, A.I.; Vreugdenhil, D.; Millenaar, F.F.; Van Ieperen, W. Phytochrome A Protects Tomato Plants from Injuries Induced by Continuous Light. Front. Plant Sci. 2019, 10, 19. [Google Scholar] [CrossRef]
- Danielson, L.L. Effect of daylength on growth and reproduction of the cucumber. Plant Physiol. 1944, 19, 638–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, N.S.; Pharr, D.M. Regulation of Photosynthetic Carbon Metabolism in Cucumber by Light Intensity and Photosynthetic Period. Plant Physiol. 1987, 85, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, G.; Gosselin, A. Influence of continuous and discontinuous supplemental lighting on the daily variation in gaseous exchange in greenhouse cucumber. Sci. Hortic. 1989, 40, 9–22. [Google Scholar] [CrossRef]
- Dorais, M. The use of supplemental lighting for vegetable crop production: Light intensity, crop response, nutrition, crop management, cultural practices. In Proceedings of the Canadian Greenhouse Conference, Niagra Falls, ON, Canada, 9 October 2003; pp. 1–8. [Google Scholar]
- Wolff, S.A.; Langerud, A. Fruit yield, starch content and leaf chlorosis in cucumber exposed to continuous lighting. Eur. J. Hortic. Sci. 2006, 71, 259–261. [Google Scholar]
- Shibaeva, T.G.; Markovskaya, E.F. Growth and development of cucumber Cucumis sativus L. in the prereproductive period under long photoperiods. Russ. J. Dev. Biol. 2013, 44, 78–85. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Hovi, T.; Näkkilä, J.; Tahvonen, R. Interlighting improves production of year-round cucumber. Sci. Hortic. 2004, 102, 283–294. [Google Scholar] [CrossRef]
- Kaukoranta, T.; Särkkä, L.; Jokinen, K. Energy efficiency of greenhouse cucumber production under LED and HPS lighting. Acta Hortic. 2017, 1170, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, R.I.; Torre, S.; Gislerød, H.R. Effects of leaf aging and light duration on photosynthetic characteristics in a cucumber canopy. Sci. Hortic. 2010, 125, 82–87. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Khosla, S.; Hao, X.; Grodzinski, B. Effect of elevated CO2 and spectral quality on whole plant gas exchange patterns in tomatoes. PLoS ONE 2018, 13, e0205861. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Von Caemmerer, S.; A Berry, J. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurtrie, R.E.; Wang, Y.P. Mathematical models of the photosynthetic response of tree stands to rising CO¬2 con-centrations and temperature. Plant Cell Environ. 1993, 16, 1–13. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Singsaas, E.L.; Pimentel, C.; Portis, A.R., Jr.; Long, S.P. Improved temperature response functions for models of rubisco-limited photosynthesis. Plant Cell Environ. 2001, 24, 253–259. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- IESO (Independent Electricity System Operator). 2020. Available online: http://reports.ieso.ca/public/PriceHOEPAverage/ (accessed on 20 July 2020).
- Hao, X.; Papadopoulos, A.P. Effects of supplemental lighting and cover materials on growth, photosynthesis, biomass partitioning, early yield and quality of greenhouse cucumber. Sci. Hortic. 1999, 80, 1–18. [Google Scholar] [CrossRef]
- Pattison, P.M.; Tsao, J.Y.; Brainard, G.C.; Bugbee, B. LEDs for photons, physiology and food. Nature 2018, 563, 493–500. [Google Scholar] [CrossRef]
- Nelson, J.A.; Bugbee, B. Economic Analysis of Greenhouse Lighting: Light Emitting Diodes vs. High Intensity Discharge Fixtures. PLoS ONE 2014, 9, e99010. [Google Scholar] [CrossRef] [Green Version]
- Hao, X.; Zheng, J.; Celeste, L.; Guo, X.; Kholsa, S. Liquid desiccant dehumidification system for improving microclimate and plant growth in greenhouse cucumber production. Acta Hortic. 2017, 1170, 861–866. [Google Scholar] [CrossRef]
- Haque, M.S.; De Sousa, A.; Soares, C.; Kjaer, K.H.; Fidalgo, F.; Rosenqvist, E.; Ottosen, C.-O. Temperature Variation under Continuous Light Restores Tomato Leaf Photosynthesis and Maintains the Diurnal Pattern in Stomatal Conductance. Front. Plant Sci. 2017, 8, 1603. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Korczynski, P.C.; Logan, J.; Faust, J.E. Mapping Monthly Distribution of Daily Light Integrals across the Contiguous United States. HortTechnology 2002, 12, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Lleqellyn, D.; Zheng, Y. Response of growth, yield and quality of pea shoots to supplemental LED lighting during winter greenhouse production. Can. J. Plant Sci. 2018, 98, 732–740. [Google Scholar] [CrossRef]
- Moe, R.; Grimstad, S.O.; Gislerød, H.A. The use of artificial light in year round production of greenhouse crops in Norway. Acta Hortic. 2006, 711, 35–42. [Google Scholar] [CrossRef]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-oxidative stress markers as a measure of abiotic stress-induced leaf se-nescence: Advantages and limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef] [Green Version]
- Millar, A.J. Input signals to the plant circadian clock. J. Exp. Bot. 2004, 55, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant Circadian Clocks Increase Photosynthesis, Growth, Survival, and Competitive Advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [Green Version]
- McClung, C.R. Plant Circadian Rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.M.; Guthrie, J.D.; Newell, J.M. Some effects of artificial climates on the growth and chemical composition of plants. Am. J. Bot. 1930, 17, 416–482. [Google Scholar] [CrossRef]
- Demers, D.-A.; Dorais, M.; Wien, C.H.; Gosselin, A. Effects of supplemental light duration on greenhouse tomato (Lycopersicum esculentum Mill.) plants and fruit yields. Sci. Hortic. 1998, 74, 295–306. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Su, Y.-S.; Lagarias, J.C. Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef] [Green Version]
- Bae, G.; Choi, G. Decoding of Light Signals by Plant Phytochromes and Their Interacting Proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef] [Green Version]
- Weller, J.L.; Schreuder, M.E.; Smith, H.; Koornneef, M.; Kendrick, R.E. Physiological interactions of phytochromes A, B1 and B2 in the control of development in tomato. Plant J. 2000, 24, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Steyn, W.J.; Wand, S.J.E.; Holcroft, D.M.; Jacobs, G. Anthocyanins in vegetative tissues: A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Ma, S.; Sun, L.; Sui, X.; Li, Y.; Chang, Y.; Fan, J.; Zhang, Z. Phloem loading in cucumber: Combined symplastic and apoplastic strategies. Plant J. 2019, 98, 391–404. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of Light Quality and Intensity on Diurnal Patterns and Rates of Photo-Assimilate Translocation and Transpiration in Tomato Leaves. Front. Plant Sci. 2018, 9, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [Green Version]
- Grodzinski, B.; Jiao, J.; Leonardos, E.D. Estimating Photosynthesis and Concurrent Export Rates in C3 and C4 Species at Ambient and Elevated CO21,2. Plant Physiol. 1998, 117, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Internode Length (cm) | Length 5th Leaf (cm) | Width 5th Leaf (cm) | Length 10th Leaf (cm) | Width 10th Leaf (cm) | Length 15th Leaf (cm) | Width 15th Leaf (cm) |
|---|---|---|---|---|---|---|---|---|
| 19 DIT | ||||||||
| T1 | 200 ± 3A | 9.3 ± 0.2A | 18.7 ± 0.3A | 19.2 ± 0.6A | 24.3 ± 0.6A | 26.2 ± 0.7A | 27.2 ± 0.7A | 30.3 ± 0.6A |
| T2 | 205 ± 5A | 9.4 ± 0.3A | 18.7 ± 0.3A | 19.0 ± 0.3A | 24.2 ± 0.7A | 25.7 ± 0.9A | 27.0 ± 1.1A | 31.2 ± 1.1A |
| T3 | 200 ± 3A | 9.1 ± 0.1A | 17.7 ± 0.3A | 17.7 ± 0.5A | 23.3 ± 0.9A | 23.7 ± 0.8A | 28.2 ± 0.8A | 32.0 ± 1.2A |
| T4 | 193 ± 5A | 8.9 ± 0.3A | 19.3 ± 0.6A | 19.3 ± 0.7A | 25.0 ± 0.5A | 26.3 ± 1.0A | 26.2 ± 0.7A | 31.7 ± 0.9A |
| 75 DIT | ||||||||
| T1 | N/A | 9.1 ± 0.2A | 19.7 ± 0.3A | 23.7 ± 0.3A | 24.2 ± 0.3A | 29.5 ± 0.6A | 27.0 ± 0.5A | 31.3 ± 0.6A |
| T2 | N/A | 9.3 ± 0.1A | 18.8 ± 0.6A | 23.0 ± 0.6A | 23.3 ± 0.5A | 28.3 ± 0.8A | 26.0 ± 0.7A | 30.7 ± 0.7A |
| T3 | N/A | 9.4 ± 0.1A | 19.5 ± 0.4A | 23.3 ± 0.3A | 23.5 ± 0.7A | 27.7 ± 1.1A | 25.7 ± 0.8A | 30.5 ± 0.8A |
| T4 | N/A | 9.3 ± 0.2A | 19.5 ± 0.6A | 24.0 ± 0.8A | 24.5 ± 0.4A | 28.8 ± 0.7A | 26.7 ± 0.8A | 31.0 ± 1.2A |
| 103 DIT | ||||||||
| T1 | N/A | 8.3 ± 0.2A | 17.5 ± 0.4A | 20.8 ± 0.4A | 25.8 ± 0.5A | 30.2 ± 0.9A | 27.8 ± 0.4A | 33.0 ± 0.8A |
| T2 | N/A | 7.9 ± 0.2A | 18.3 ± 0.4A | 20.8 ± 0.3A | 25.3 ± 0.3A | 30.5 ± 0.9A | 26.3 ± 0.5A | 32.3 ± 0.2A |
| T3 | N/A | 8.0 ± 0.2A | 17.0 ± 0.4A | 20.5 ± 0.5A | 24.7 ± 0.7A | 29.3 ± 0.8A | 27.7 ± 0.5A | 31.2 ± 0.7A |
| T4 | N/A | 8.0 ± 0.1A | 17.2 ± 0.2A | 20.5 ± 0.2A | 25.7 ± 0.7A | 30.5 ± 0.8A | 27.3 ± 0.6A | 33.5 ± 1.0A |
| Light Treatment | Respiration (µmol CO2 m−2 s−1) | LCP (µmol m−2 s−1) | QY (µmol CO2 m−2 s−1/ µmol m−2 s−1) | Pnmax (µmol CO2 m−2 s−1) |
|---|---|---|---|---|
| 20 DIT | ||||
| T1 | −1.73 ± 0.49A | 36.94 ± 8.56A | 0.046 ± 0.001A | 25.98 ± 1.34A |
| T2 | −2.65 ± 0.21A | 51.48 ± 4.22A | 0.051 ± 0.004A | 26.12 ± 0.72A |
| T3 | −2.18 ± 0.60A | 38.14 ± 8.70A | 0.055 ± 0.001A | 26.41 ± 1.73A |
| T4 | −1.92 ± 0.10A | 35.13 ± 3.64A | 0.051 ± 0.004A | 25.80 ± 1.21A |
| 99 DIT | ||||
| T1 | −2.13 ± 0.10A | 46.53 ± 0.10A | 0.042 ± 0.002A | 17.69 ± 0.60A |
| T2 | −2.76 ± 0.23A | 57.88 ± 8.00A | 0.049 ± 0.005A | 16.23 ± 1.45A |
| T3 | −2.59 ± 0.06A | 55.39 ± 3.08A | 0.045 ± 0.002A | 17.37 ± 0.89A |
| T4 | −2.41 ± 0.04A | 50.87 ± 1.73A | 0.047 ± 0.002A | 18.01 ± 0.48A |
| Light Treatment | Vcmax (µmol CO2 m−2 s−1) | Jmax (µmol e- m−2 s−1) | Jmax:Vcmax |
|---|---|---|---|
| 19 DIT | |||
| T1 | 40.51 ± 0.92AB | 119.17 ± 3.44A | 2.94 ± 0.03A |
| T2 | 36.89 ± 1.35B | 107.53 ± 4.07B | 2.91 ± 0.02A |
| T3 | 40.05 ± 2.06AB | 116.04 ± 5.15AB | 2.90 ± 0.03A |
| T4 | 42.89 ± 1.63A | 125.44 ± 5.42A | 2.92 ± 0.02A |
| 96 DIT | |||
| T1 | 36.93 ± 0.61A | 108.15 ± 1.83A | 2.93 ± 0.003A |
| T2 | 24.63 ± 1.72C | 71.08 ± 5.29B | 2.88 ± 0.02A |
| T3 | 30.84 ± 0.63B | 89.12 ± 1.69AB | 2.89 ± 0.01A |
| T4 | 36.51 ± 1.56AB | 106.00 ± 5.61A | 2.90 ± 0.04A |
| Yield Period | Light Treatment | Fruit Number (m−2) | Total Fruit Weight (kg m−2) | Average Fruit Weight (g Fruit−1) |
|---|---|---|---|---|
| 19 November 2019–27 December 2019 | T1 | 149A | 12.02A | 80.67A |
| T2 | 145A | 11.78A | 81.24A | |
| T3 | 147A | 11.92A | 81.09A | |
| T4 | 148A | 11.82A | 79.86A | |
| 28 December 2019–7 February 2020 | T1 | 149A | 11.61A | 77.92A |
| T2 | 149A | 11.52A | 77.32A | |
| T3 | 147A | 11.38A | 77.41A | |
| T4 | 152A | 11.64A | 76.58A | |
| 8 February 2020–20 March 2020 | T1 | 149A | 12.87A | 86.38A |
| T2 | 155A | 13.03A | 84.06A | |
| T3 | 155A | 13.19A | 85.10A | |
| T4 | 157A | 13.29A | 84.65A | |
| 19 November 2019–20 March 2020 | T1 | 448A | 36.50A | 81.50A |
| T2 | 449A | 36.34A | 80.94A | |
| T3 | 448A | 36.49A | 81.45A | |
| T4 | 456A | 36.76A | 80.61A |
| Light Treatment | T1 | T2 | T3 | T4 |
|---|---|---|---|---|
| Total electrical consumption (kWh) | 6184 | 6184 | 6184 | 6184 |
| On-peak electrical consumption (kWh) Z | 2319 | 1546 | 1988 | 2282 |
| Mid-peak electrical consumption (kWh) Y | 2319 | 1546 | 1988 | 2650 |
| Off-peak electrical consumption (kWh) X | 1546 | 3092 | 2209 | 1251 |
| Total electricity cost | $105.70 | $95.94 | $101.52 | $106.68 |
| Total electricity cost per m2 of grow area | $4.52 | $4.10 | $4.34 | $4.56 |
| Total electricity cost per kg of production v | ¢12.38 | ¢11.28 | ¢11.89 | ¢12.40 |
| Total electricity cost per fruit W | ¢1.01 | ¢0.91 | ¢0.97 | ¢1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanoue, J.; Zheng, J.; Little, C.; Grodzinski, B.; Hao, X. Continuous Light Does Not Compromise Growth and Yield in Mini-Cucumber Greenhouse Production with Supplemental LED Light. Plants 2021, 10, 378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020378
Lanoue J, Zheng J, Little C, Grodzinski B, Hao X. Continuous Light Does Not Compromise Growth and Yield in Mini-Cucumber Greenhouse Production with Supplemental LED Light. Plants. 2021; 10(2):378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020378
Chicago/Turabian StyleLanoue, Jason, Jingming Zheng, Celeste Little, Bernard Grodzinski, and Xiuming Hao. 2021. "Continuous Light Does Not Compromise Growth and Yield in Mini-Cucumber Greenhouse Production with Supplemental LED Light" Plants 10, no. 2: 378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020378