
Integrative Taxonomy and Molecular Phylogeny of the Plant-Parasitic Nematode Genus Paratylenchus (Nematoda: Paratylenchinae): Linking Species with Molecular Barcodes


Abstract
:1. Introduction
2. Results
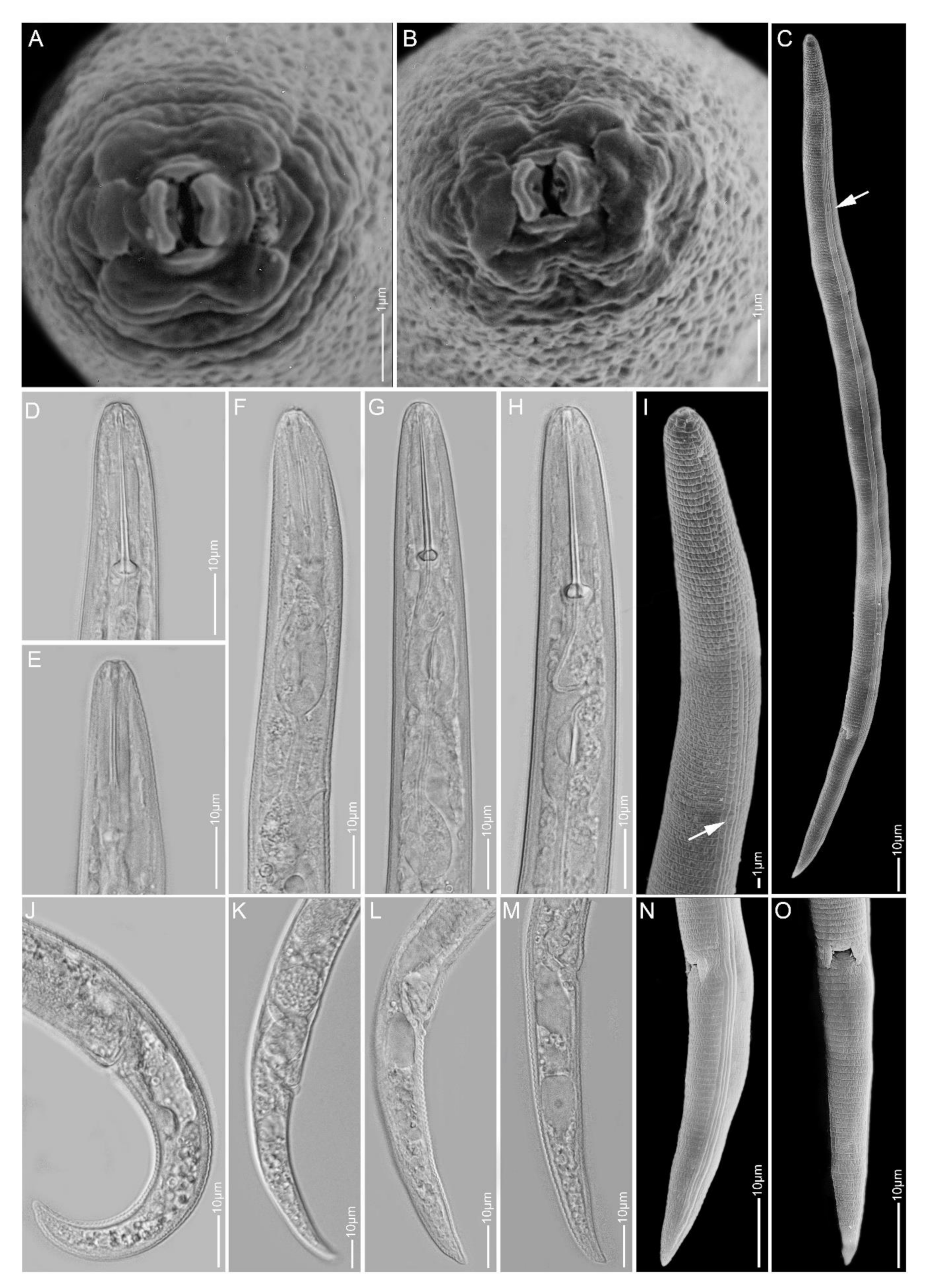
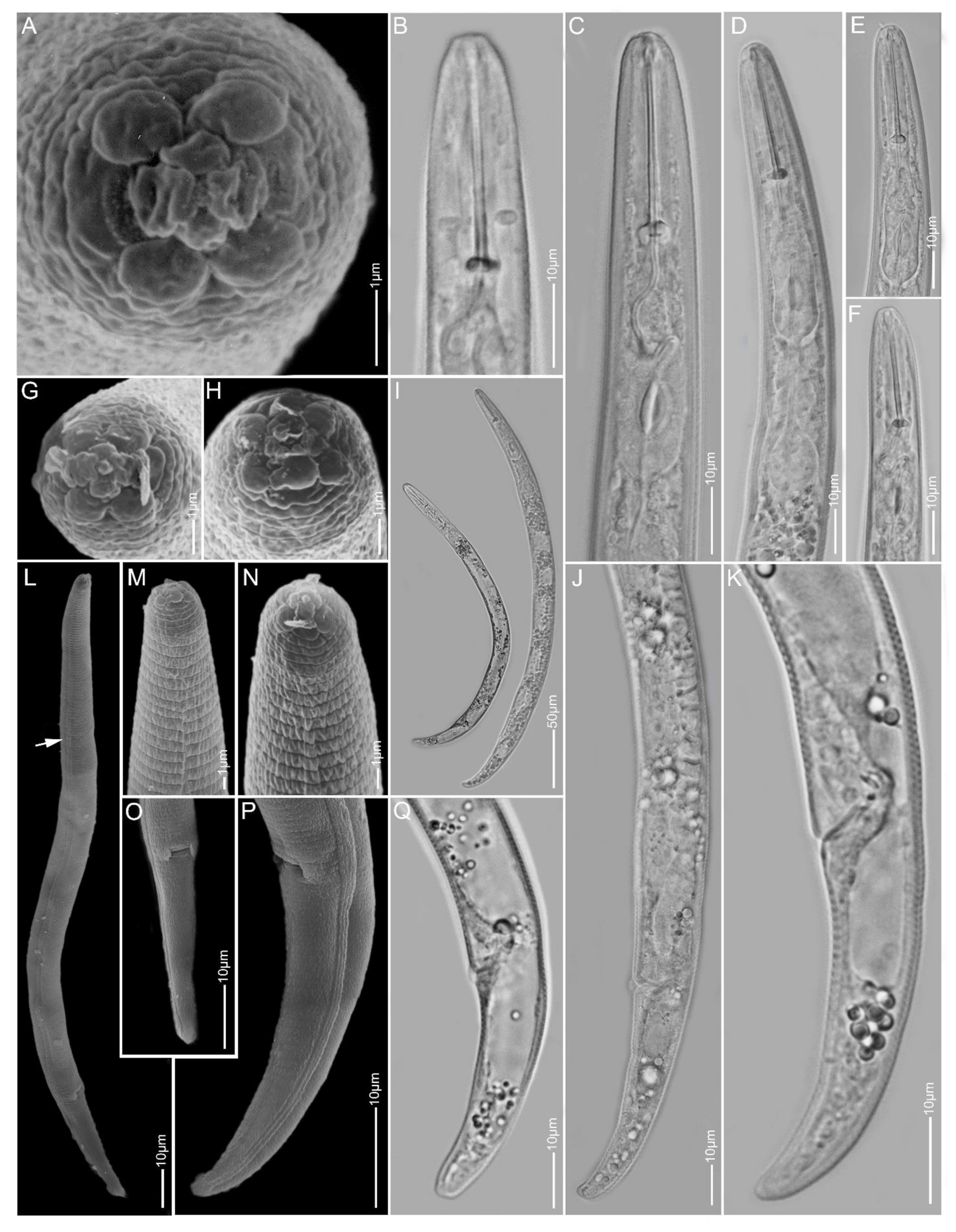
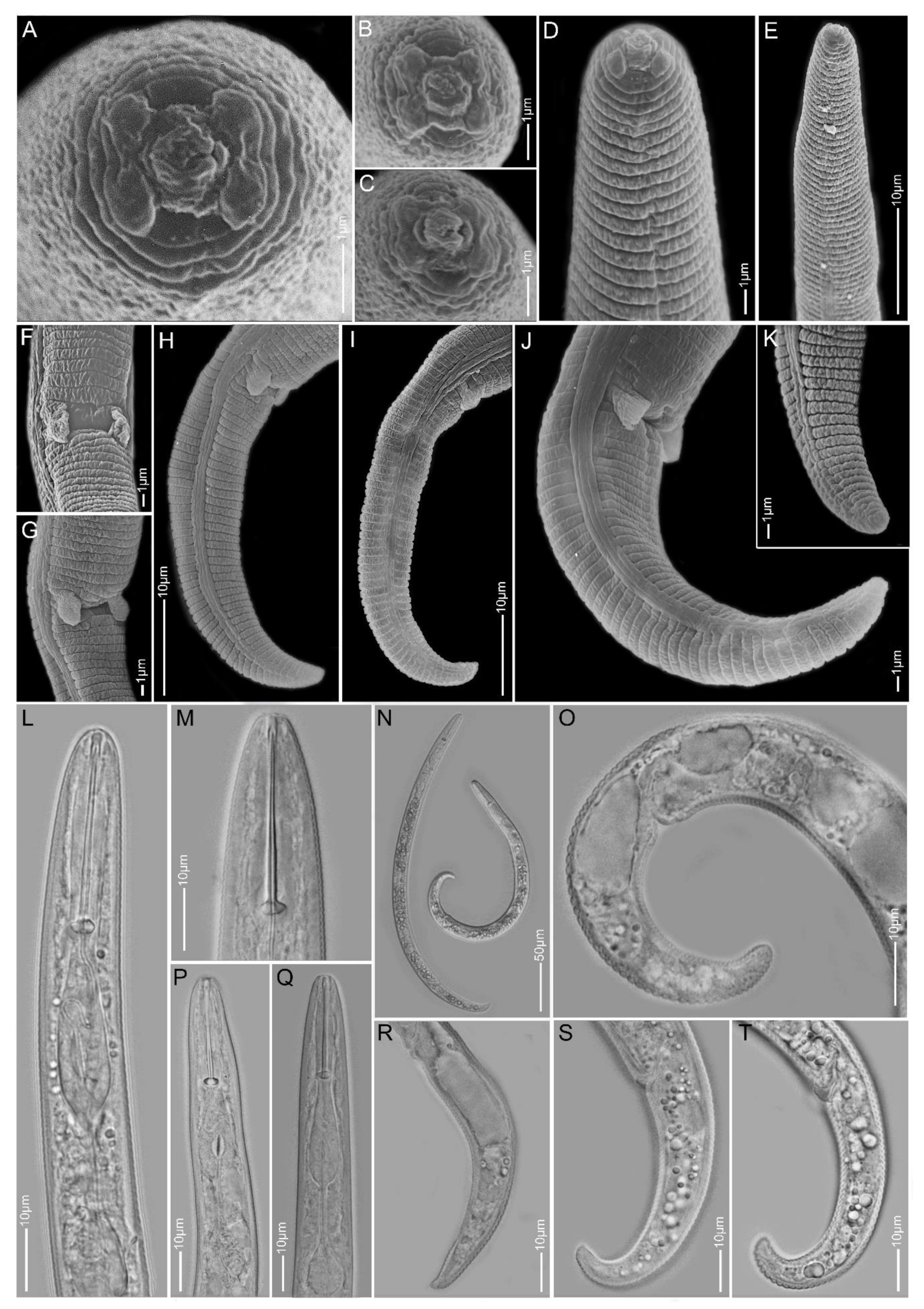
2.1. Species Identification, Characterisation and Delimitation
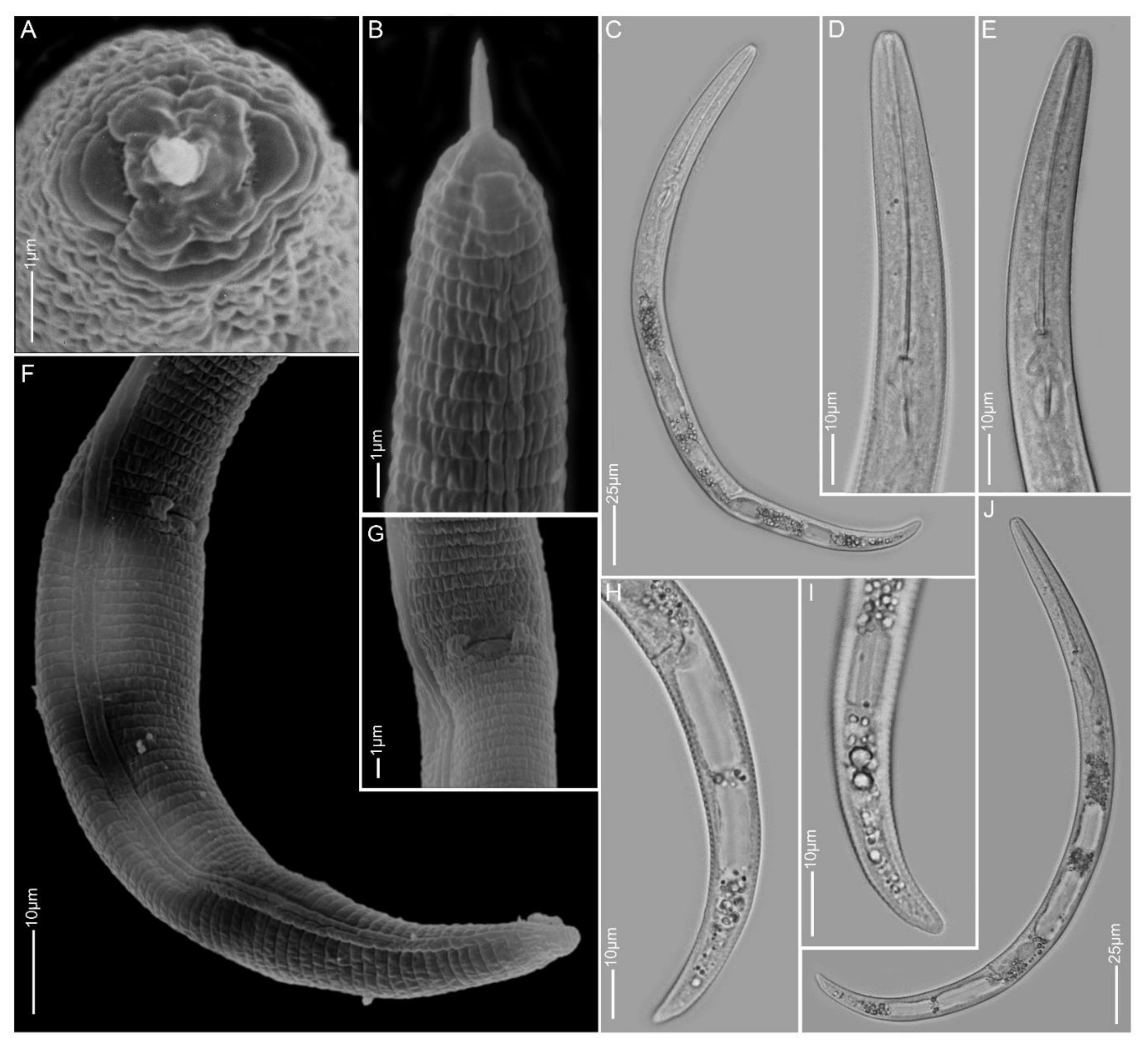
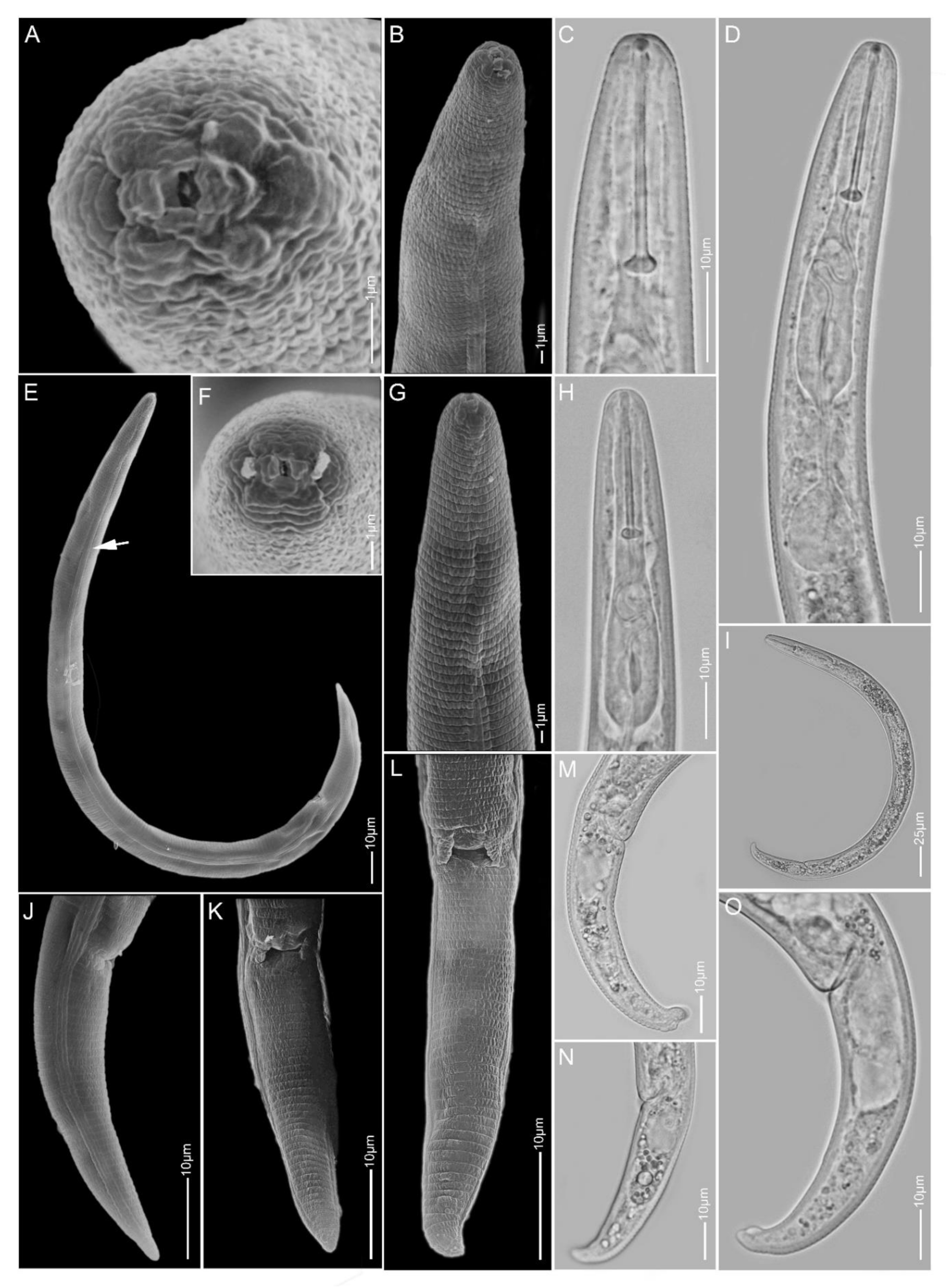
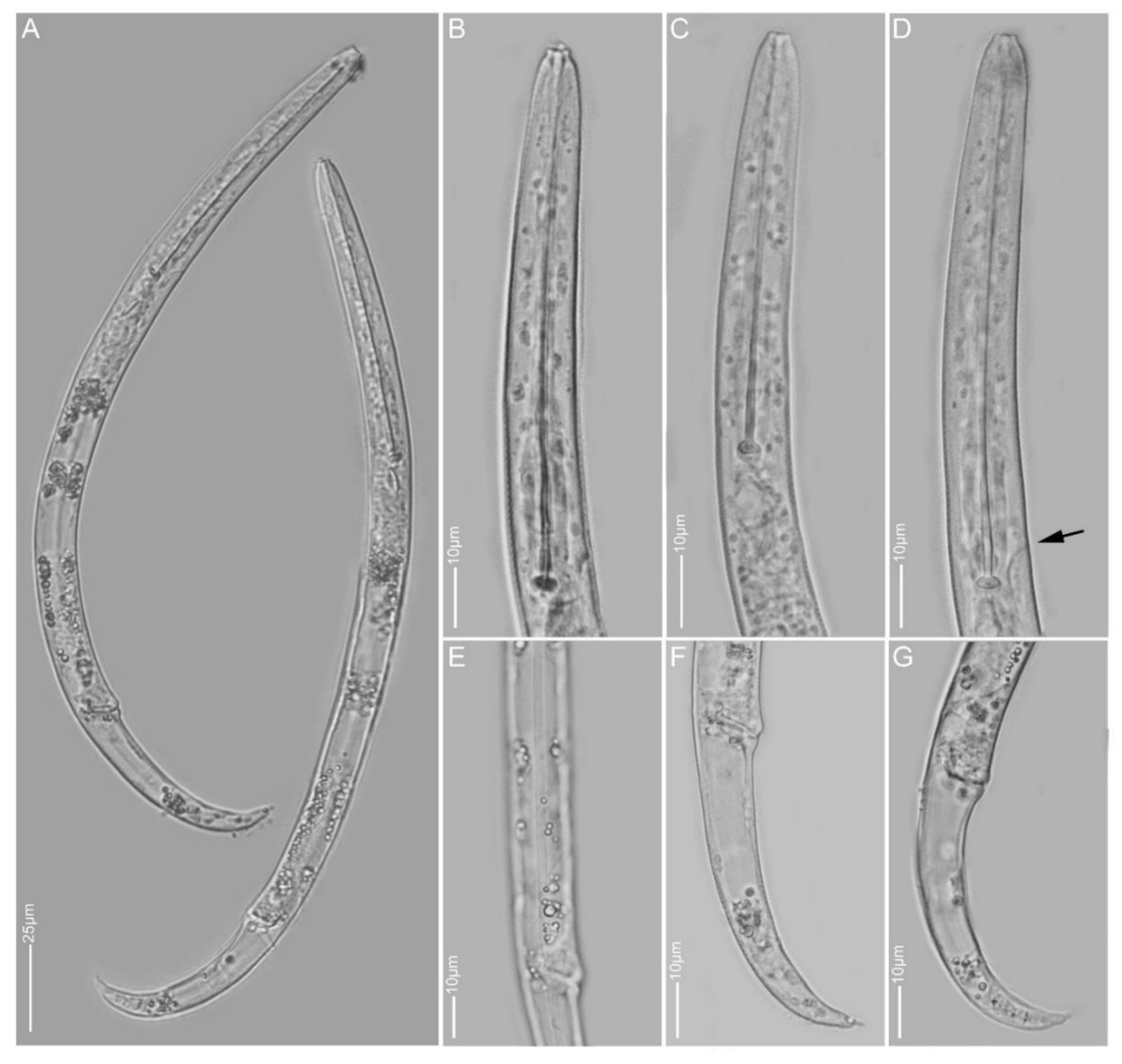
2.1.1. Paratylenchus aculentus
2.1.2. Paratylenchus elachistus
2.1.3. Paratylenchus goodeyi
2.1.4. Paratylenchus holdemani
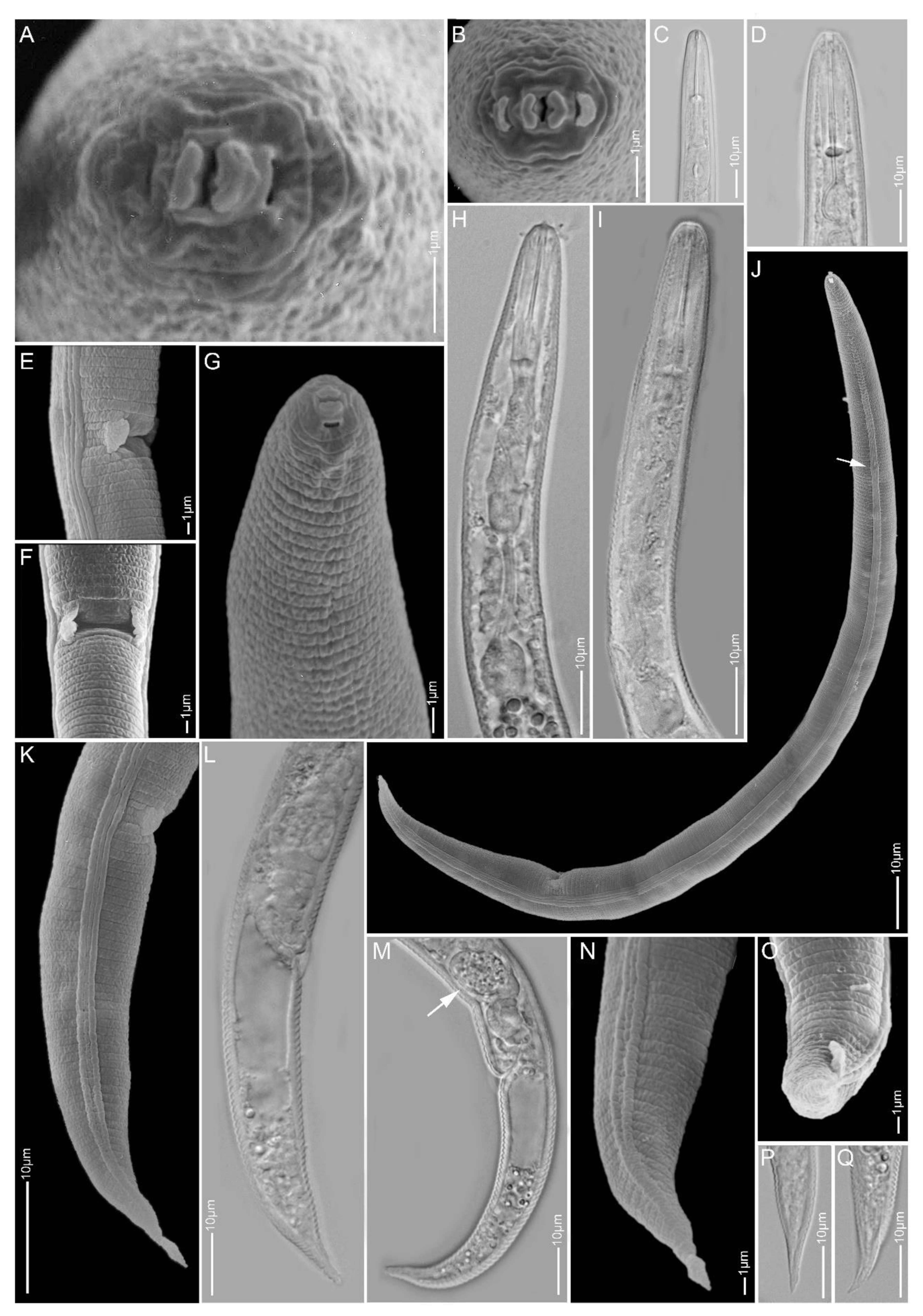
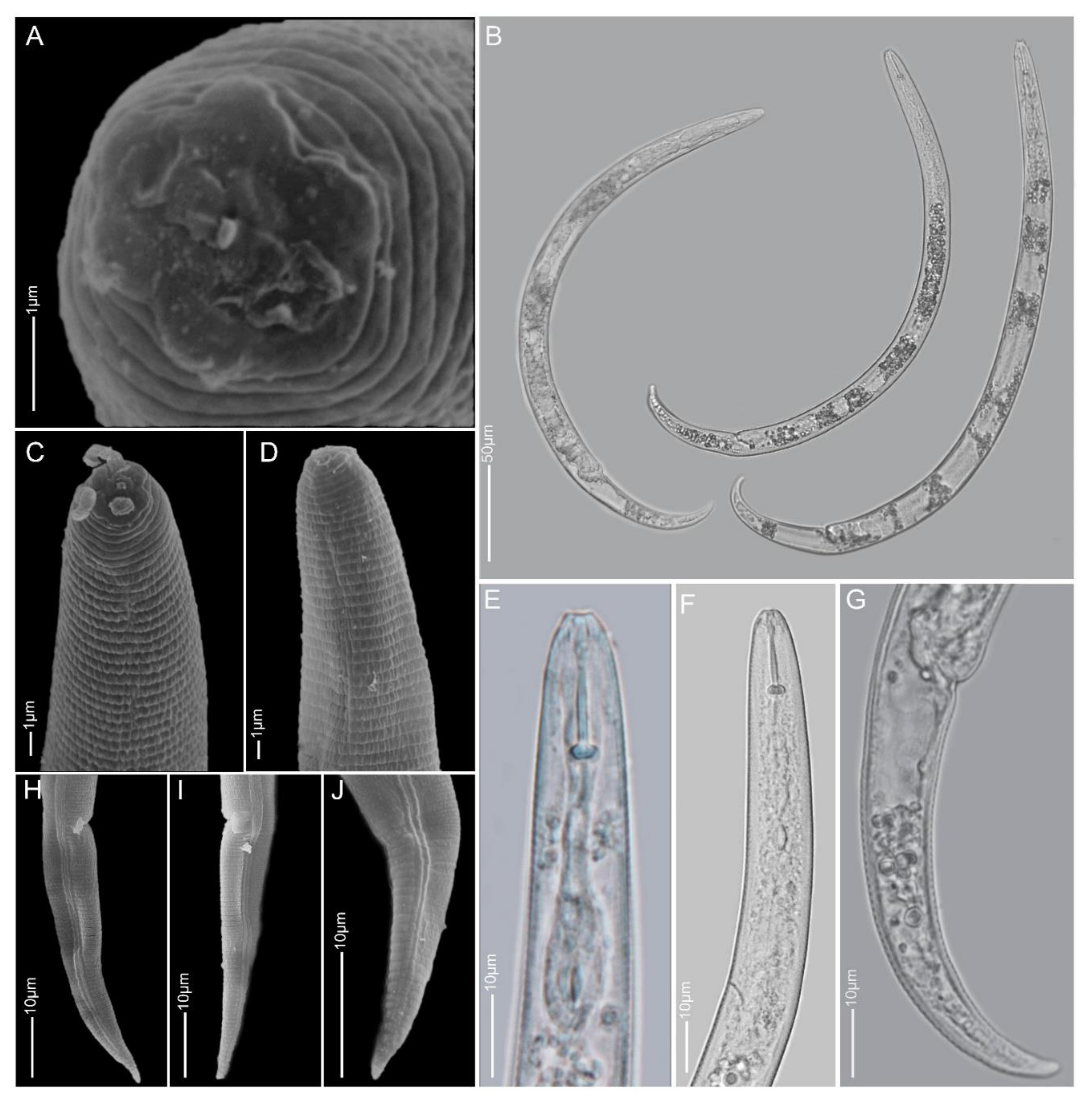
2.1.5. Paratylenchus idalimus
2.1.6. Paratylenchus microdorus
2.1.7. Paratylenchus nanus
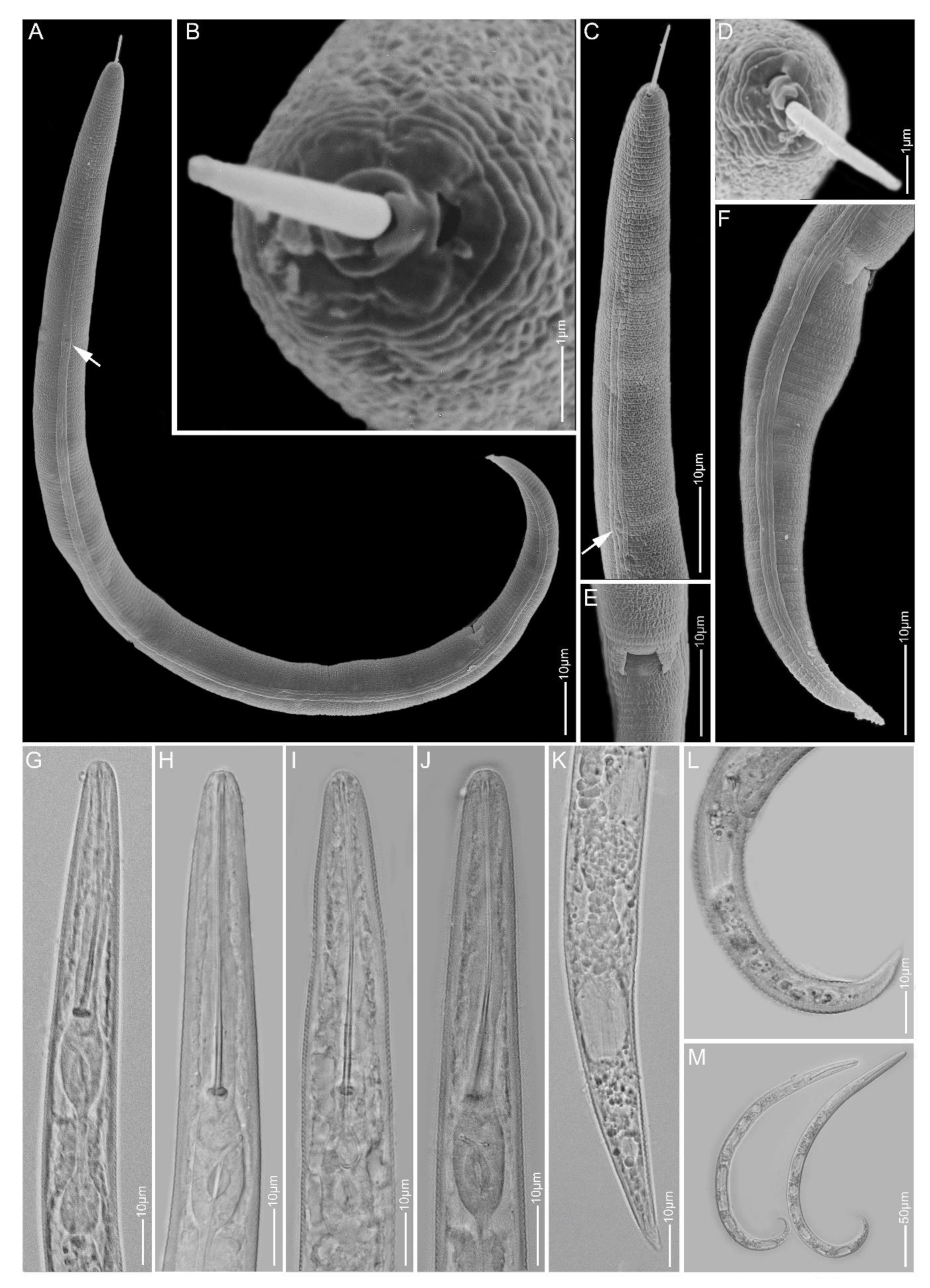
2.1.8. Paratylenchus neoamblycephalus
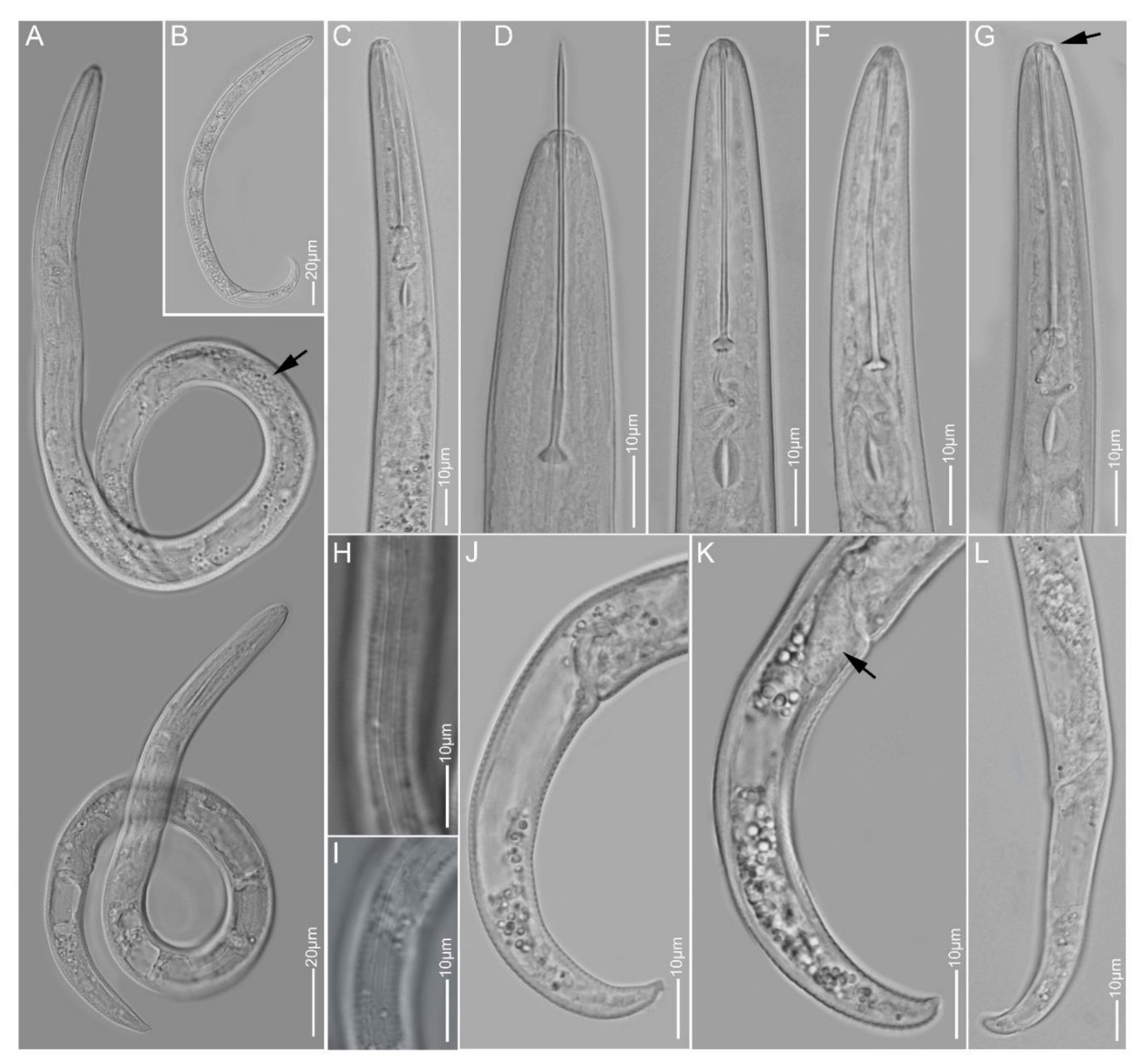
2.1.9. Paratylenchus straeleni
2.1.10. Paratylenchus veruculatus
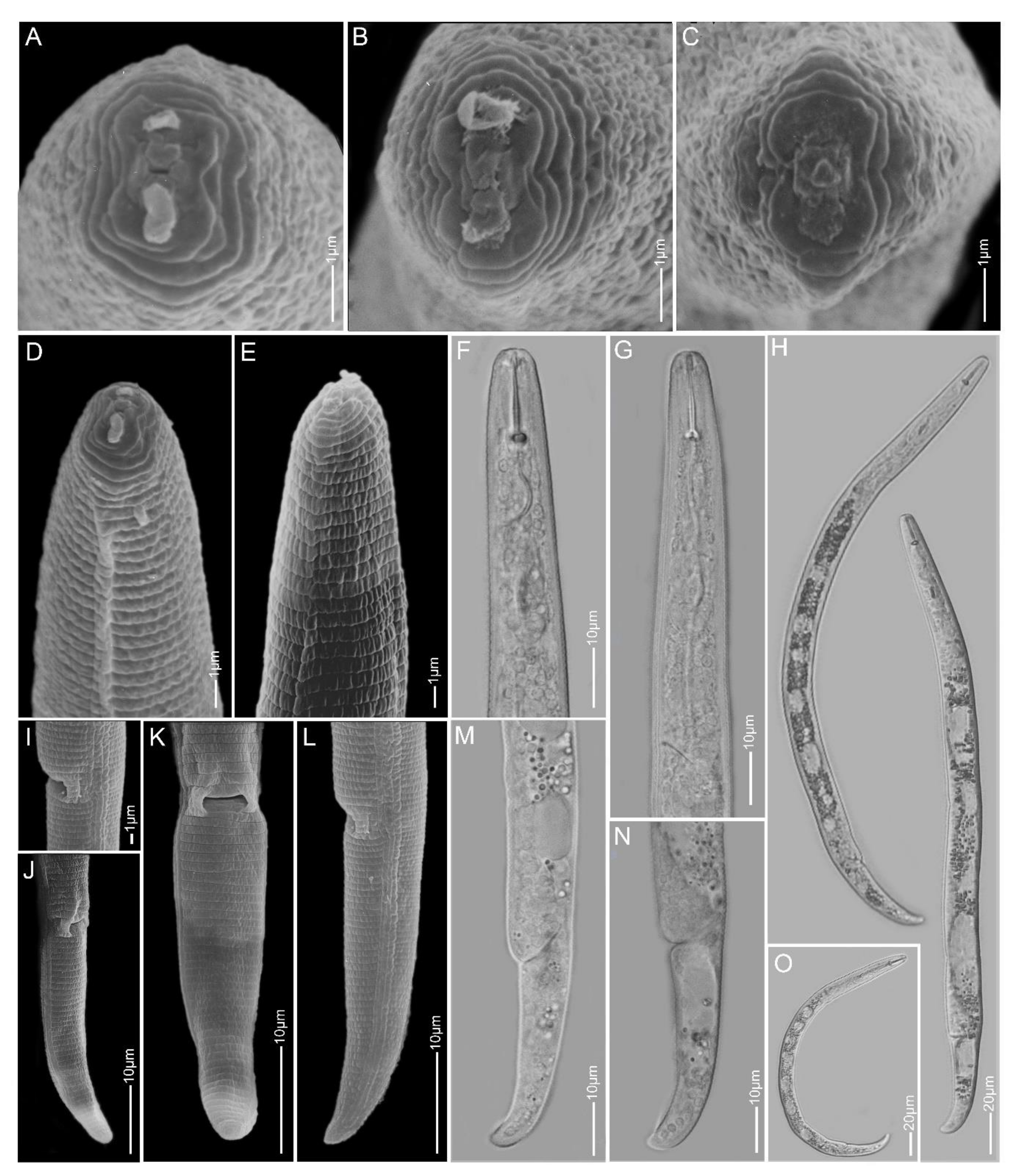
2.1.11. Paratylenchus sp.2
2.1.12. Paratylenchus sp.BE11
2.1.13. Paratylenchus sp.D
2.1.14. Paratylenchus sp.F
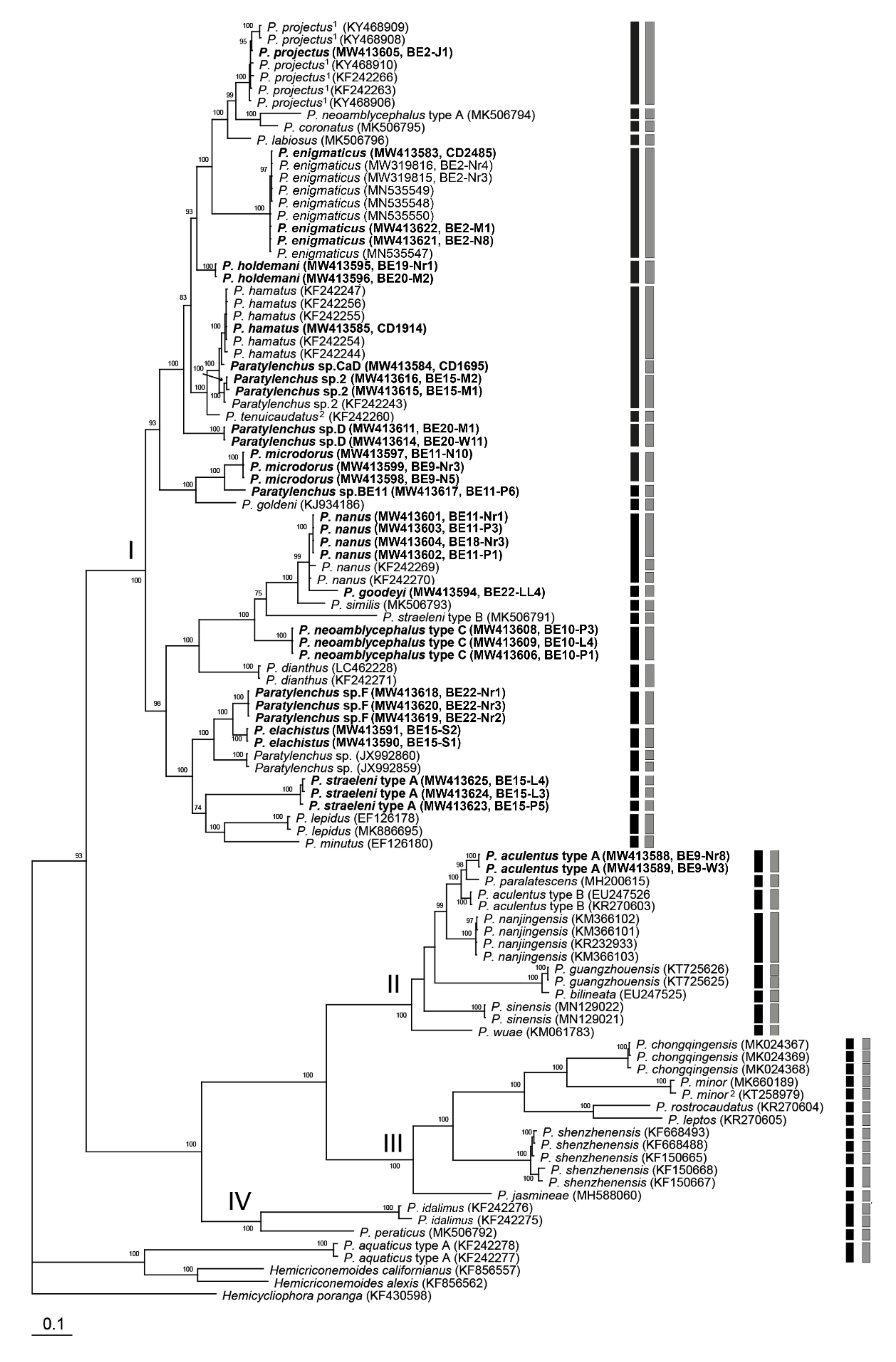
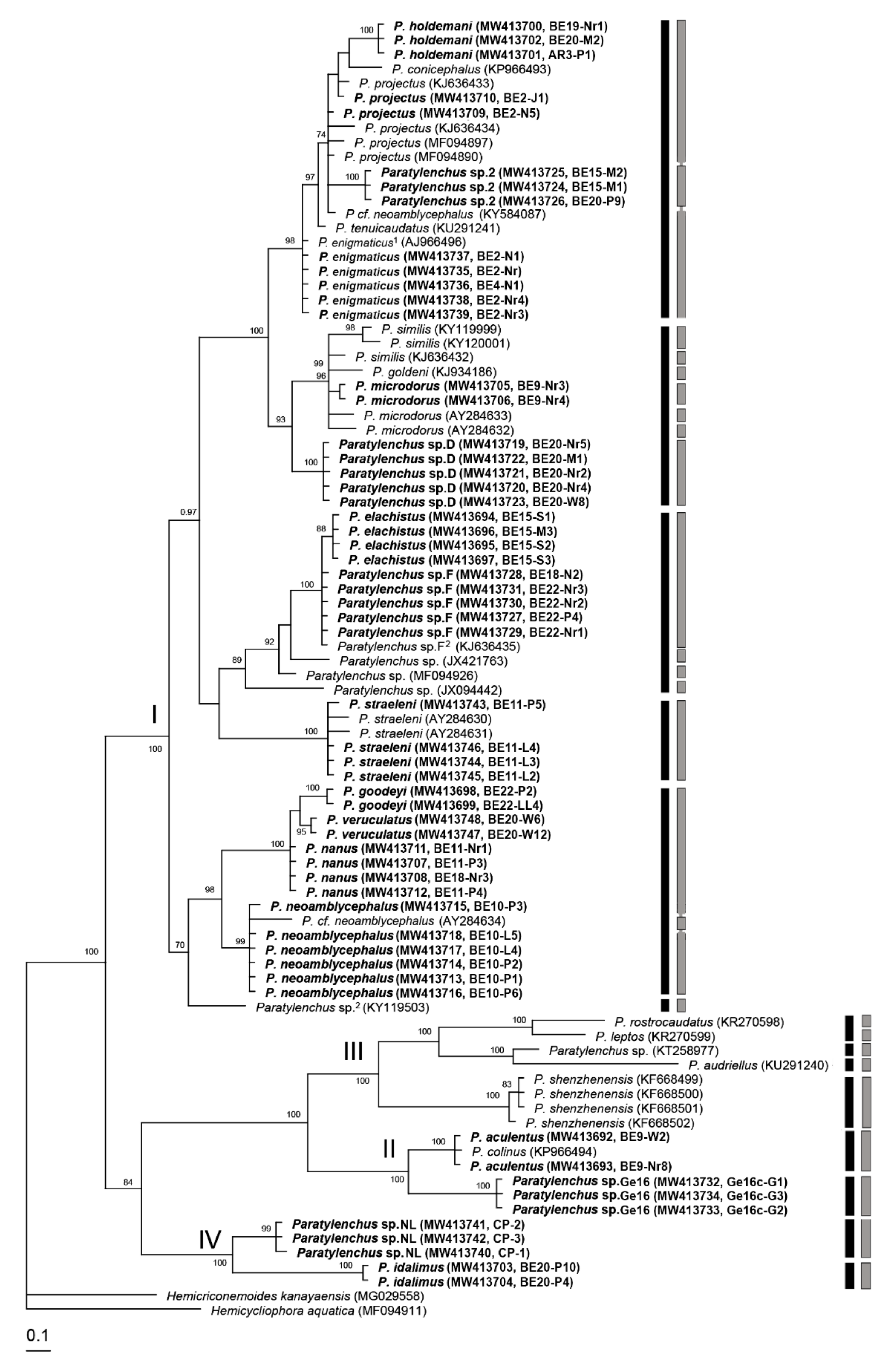
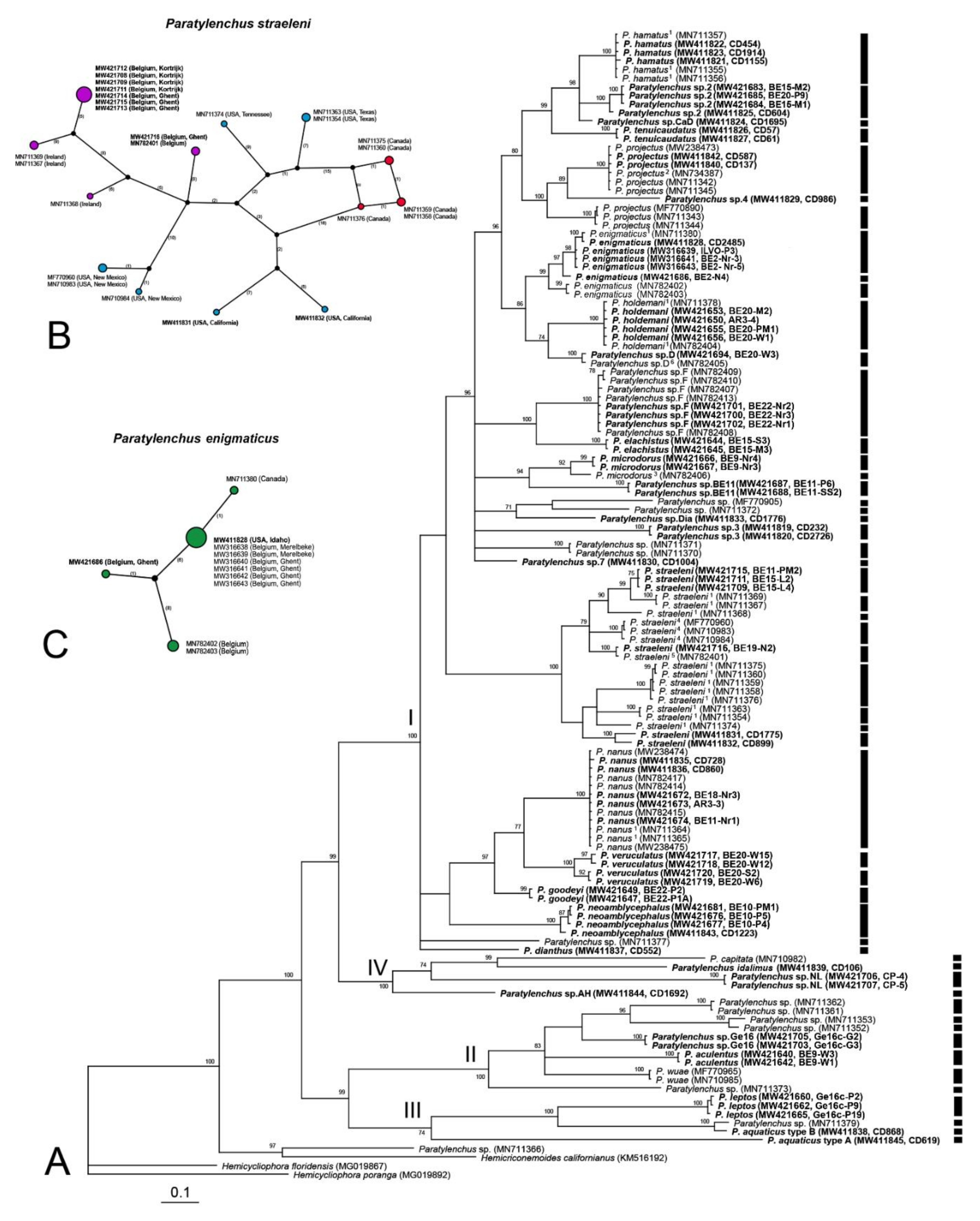
2.2. Phylogenetic and Species Delimitation Analysis
3. Discussion
4. Materials and Methods
4.1. Nematode Populations
4.2. Morphological Study
4.3. Extraction of DNA, PCR and Sequencing
4.4. Phylogenetic and Species Delimitation Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Micoletzky, H. Die freilebenden Erd-Nematoden; Archiv für Naturgeschichte: Berlin, Germany, 1922. [Google Scholar]
- Siddiqi, M.R. Tylenchida: Parasites of Plants and Insects, 2nd ed.; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Manzanilla-López, R.H.; Marbán-Mendoza, N. Practical plant nematology; BBA: Colegio de Postgraduados, Mexico, 2012. [Google Scholar]
- Fourie, H.; Spaull, V.W.; Jones, R.K.; Daneel, M.S.; De Waele, D. Nematology in South Africa: A View from the 21st Century; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Subbotin, S.A.; Chitambar, J.J. Plant Parasitic Nematodes in Sustainable Agriculture of North America; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Tarjan, A.C. A review of the genus Paratylenchus Micoletzky, 1922 (Paratylenchinae: Nematoda) with a description of two new species. Ann. N. Y. Acad. Sci. 1960, 84, 329–390. [Google Scholar] [CrossRef]
- Raski, D.J. Paratylenchidae n. fam. with descriptions of five new species of Gracilacus n. g. and an emendation of Cacopaurus Thorne, 1943, Paratylenchus Micoletzky, 1922 and Criconematidae Thorne, 1943. Proc. Helm. Soc. Wash. 1962, 29, 189–207. [Google Scholar]
- Thorne, G.; Malek, R.B. Nematodes of the Northern Great Plains part I. Tylenchida [Nemata Secernentra]; South Dakota State University, Brookings: Sioux Falls, SD, USA, 1968. [Google Scholar]
- Raski, D.J.; Luc, M. A reappraisal of Tylenchina (Nemata). 10: The superfamily Criconematoidea Taylor, 1936. Rev. Nématol. 1987, 10, 409–444. [Google Scholar]
- Maggenti, A.R.; Luc, M.; Raski, D.J.; Fortuner, R.; Geraert, E. A reappraisal of Tylenchina (Nemata). 11. List of generic and supra-generic taxa, with their junior synonyms. Rev. Nématol. 1988, 11, 177–188. [Google Scholar]
- Raski, D.J. Tylenchulidae of agricultural soils. In Manual of Agricultural Nematology; Nickle, W.R., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1991; pp. 761–794. [Google Scholar]
- Esser, R.P. A diagnostic compendium to species included in Paratylenchinae Thorne, 1949 and Tylenchocriconematinae Raski & Siddiqi, 1975 (Nematoda: Criconematoidea). Nematologica 1992, 38, 146–163. [Google Scholar]
- Andrássy, I. Free-living nematodes of Hungary, II (Nematoda, Errantia); Hungarian Natural History Museum: Budapest, Hungary, 2007. [Google Scholar]
- Yu, Q.; Ye, W.; Powers, T. Morphological and Molecular Characterization of Gracilacus wuae n. sp. (Nematoda: Criconematoidea) Associated with Cow Parsnip (Heracleum maximum) in Ontario, Canada. J. Nematol. 2016, 48, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Siddiqi, M.R. Tylenchida: Parasites of Plants and Insects; Commonwealth Agricultural Bureaux: Slough, UK, 1986. [Google Scholar]
- Brzeski, M.W.; Szczygiel, A. Studies On the Nematodes of the Genus Paratylenchus Micoletzky (Nematoda: Paratylenchinae) in Poland. Nematologica 1963, 9, 613–625. [Google Scholar] [CrossRef]
- Siddiqi, M.R.; Goodey, J.B. The status of the genera and subfamilies of the Criconematidae (Nematoda); with a comment on the position of Fergusobia. Nematologica 1964, 9, 363–377. [Google Scholar] [CrossRef]
- Geraert, E. The genus Paratylenchus. Nematologica 1965, 11, 301–334. [Google Scholar] [CrossRef]
- Brzeski, M.W. Nematodes of Tylenchina in Poland and temperate Europe; Muzeum i Instytut Zoologii, Polska Akademia Nauk: Warszawa, Poland, 1998. [Google Scholar]
- Nguyen, C.N.; Baldwin, J.G.; Choi, Y.E. New records of Paratylenchus Micoletzky, 1922 (Nematoda: Paratylenchinae) from Viet Nam with description of Paratylenchus laocaiensis sp. n. J. Nematode Morphol. Syst. 2004, 7, 51–75. [Google Scholar]
- Decraemer, W.; Hunt, D.J. Structure and classification. In Plant nematology; Perry, R.N., Moens, M., Eds.; CABI Publishing: Wallingford, UK, 2006; pp. 3–32. [Google Scholar]
- Van den Berg, E.; Tiedt, L.R.; Subbotin, S.A. Morphological and molecular characterisation of several Paratylenchus Micoletzky, 1922 (Tylenchida: Paratylenchidae) species from South Africa and USA, together with some taxonomic notes. Nematology 2014, 16, 323–358. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, R.; Kashi, L.; Karegar, A. Contribution to the study of the genus Paratylenchus Micoletzky, 1922 sensu lato (Nematoda: Tylenchulidae). Zootaxa 2014, 3841, 151–187. [Google Scholar] [CrossRef] [Green Version]
- Hesar, A.M.; Karegar, A.; Ghaderi, R. Phylogenetic relationships of Cacopaurus pestis Thorne, 1943 within representatives of the Tylenchulidae Skarbilovich, 1947 as inferred from ITS and D2-D3 expansion segments of 28S-rRNA sequences. Nematology 2019, 21, 971–994. [Google Scholar] [CrossRef]
- Maria, M.; Miao, W.; Castillo, P.; Zheng, J. A new pin nematode, Paratylenchus sinensis n. sp. (Nematoda: Paratylenchinae) in the rhizosphere of white mulberry from Zhejiang Province, China. Eur. J. Plant Pathol. 2020, 156, 1023–1039. [Google Scholar] [CrossRef]
- Raski, D.J. Paratylenchoides gen. n. and two new species (Nematoda: Paratylenchidae). Proc. Helm. Soc. Wash. 1973, 40, 230–233. [Google Scholar]
- Ganguly, S.; Khan, E. Gracilpaurus corbetti gen. n., sp. n. (Nematoda: Tylenchida: Paratylenchidae) from India with a key to the species of this genus. Pak. J. Nematol. 1990, 8, 65–71. [Google Scholar]
- Thorne, G. Cacopaurus pestis, n. g., n. spec. (Nematoda: Criconematinae), a destructive parasite of the walnut, Juglans regia Linn. Proc. Helm. Soc. Wash. 1943, 10, 78–83. [Google Scholar]
- Goodey, T. Soil and Freshwater Nematodes, 2nd ed.; Goodey, J.B., Ed.; Methuen: London, UK, 1963; p. 544. [Google Scholar]
- Ebsary, B.A. Catalog of the Order Tylenchida (Nematoda); Agriculture Canada, Research Branch: Ottawa, ON, Canada, 1991. [Google Scholar]
- Ghaderi, R.; Geraert, E.; Karegar, A. The Tylenchulidae of the world: Identification of the family Tylenchulidae (Nematoda: Tylenchida); Academia Press: Ghent, Belgium, 2016. [Google Scholar]
- Wang, K.; Li, Y.; Xie, H.; Xu, C.L.; Wu, W.J. Morphology and molecular analysis of Paratylenchus guangzhouensis n. sp. (Nematoda: Paratylenchinae) from the soil associated with Bambusa multiplex in China. Eur. J. Plant Pathol. 2016, 145, 255–264. [Google Scholar] [CrossRef]
- Wang, K.; Xie, H.; Li, Y.; Wu, W.J.; Xu, C.L. Morphology and molecular analysis of Paratylenchus nanjingensis n. sp. (Nematoda: Paratylenchinae) from the rhizosphere soil of Pinus massoniana in China. J. Helminthol. 2016, 90, 166–173. [Google Scholar] [CrossRef]
- Maria, M.; Cai, R.; Ye, W.; Powers, T.O.; Zheng, J. Description of Gracilacus paralatescens n. sp. (Nematoda: Paratylenchinae) found from the rhizosphere of Bamboo in Zhejiang, China. J. Nematol. 2018, 50, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, K.; Xie, H.; Xu, C.L. Morphology and molecular analysis of Paratylenchus chongqingensis n. sp. (Nematoda: Paratylenchinae) from soil associated with Ophiopogon japonicus in China. Eur. J. Plant Pathol. 2019, 154, 597–605. [Google Scholar] [CrossRef]
- Phani, V.; Somvanshi, V.S.; Rao, U.; Khan, M.R. Paratylenchus jasmineae sp. n. (Nematoda: Paratylenchinae) from rhizosphere of Jasminum sambac in India. Nematology 2019, 21, 469–478. [Google Scholar] [CrossRef]
- Subbotin, S.A.; Vovlas, N.; Crozzoli, R.; Sturhan, D.; Lamberti, F.; Moens, M.; Baldwin, J.G. Phylogeny of Criconematina Siddiqi, 1980 (Nematoda: Tylenchida) based on morphology and D2-D3 expansion segments of the 28SrRNA gene sequences with application of a secondary structure model. Nematology 2005, 7, 927–944. [Google Scholar]
- Chen, D.Y.; Ni, H.F.; Tsay, T.T.; Yen, J.H. Identification of Gracilacus bilineata and G. aculenta (Nematoda: Criconematoidea, Tylenchulidae) among bamboo plantations in Taiwan. Plant Pathol. Bull. 2008, 17, 209–219. [Google Scholar]
- Chen, D.Y.; Ni, H.F.; Yen, J.H.; Tsay, T.T. Identification of a new recorded pin nematode Paratylenchus minutus (Nematoda: Criconematoidea, Tylenchulidae) in Taiwan. Plant Pathol. Bull. 2009, 18, 167–174. [Google Scholar]
- van Megen, H.; van den Elsen, S.; Holterman, M.; Karssen, G.; Mooyman, P.; Bongers, T.; Holovachov, O.; Bakker, L.; Helder, J. A phylogenetic tree of nematodes based on about 1200 full-length small subunit ribosomal DNA sequences. Nematology 2009, 11, 927–950. [Google Scholar] [CrossRef]
- López, M.A.C.; Robbins, R.T.; Szalanski, A.L. Taxonomic and molecular identification of Hemicaloosia, Hemicycliophora, Gracilacus and Paratylenchus species (Nematoda: Criconematidae). J. Nematol. 2013, 45, 145–171. [Google Scholar]
- Akyazi, F.; Felek, A.F.; Čermák, V.; Čudejková, M.; Foit, J.; Yildiz, S.; Háněl, L. Description of Paratylenchus (Gracilacus) straeleni (De Coninck, 1931) Oostenbrink, 1960 (Nematoda: Criconematoidea, Tylenchulidae) from hazelnut in Turkey and its comparison with other world populations. Helminthologia 2015, 52, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Esmaeili, M.; Heydari, R.; Castillo, P.; Bidhendi, M.Z.; Palomares-Rius, J.E. Molecular characterisation of two known species of Paratylenchus Micoletzky, 1922 from Iran with notes on the validity of Paratylenchus audriellus Brown, 1959. Nematology 2016, 18, 591–604. [Google Scholar] [CrossRef]
- Zhuo, K.; Liu, X.; Tao, Y.; Wang, H.; Lin, B.; Liao, J. Morphological and molecular characterisation of three species of Paratylenchus Micoletzky, 1922 (Tylenchida: Paratylenchidae) from China, with a first description of the male P. rostrocaudatus. Nematology 2018, 20, 837–850. [Google Scholar] [CrossRef]
- Maria, M.; Miao, W.; Ye, W.; Zheng, J. Updated description of Paratylenchus lepidus Raski 1975 and P. minor Sharma, Sharma and Khan, 1986 by integrating molecular and ultra-structural observations. J. Nematol. 2019, 51, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mirbabaei, H.; Eskandari, A.; Ghaderi, R. On the synonymy of Trophotylenchulus asoensis and T. okamotoi with T. arenarius, and intra-generic structure of Paratylenchus (Nematoda: Tylenchulidae). J. Nematol. 2019, 51, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Etongwe, C.M.; Singh, P.R.; Bert, W.; Subbotin, S.A. Molecular characterisation of some plant-parasitic nematodes (Nematoda: Tylenchida) from Belgium. Russ. J. Nematol. 2020, 28, 1–28. [Google Scholar]
- Mwamula, A.O.; Kabir, M.F.; Lee, G.; Choi, I.H.; Kim, Y.H.; Bae, E.J.; Lee, D.W. Morphological characterisation and molecular phylogeny of several species of Criconematina (Nematoda: Tylenchida) associated with turfgrass in Korea, as inferred from ribosomal and mitochondrial DNA. Nematology 2020, 22, 939–956. [Google Scholar] [CrossRef]
- Powers, T.; Harris, T.S.; Higgins, R.S.; Mullin, P.G.; Powers, K.S. Nematode biodiversity assessments need vouchered databases: A BOLD reference library for plant-parasitic nematodes in the superfamily Criconematoidea. Genome 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Chen, P.J. First Report of a Pin Nematode (Paratylenchus dianthus) on Chrysanthemum in Taiwan. Plant Dis. 2020, 104, 995. [Google Scholar] [CrossRef]
- Janssen, T.; Karssen, G.; Couvreur, M.; Waeyenberge, L.; Bert, W. The pitfalls of molecular species identification: A case study within the genus Pratylenchus (Nematoda: Pratylenchidae). Nematology 2017, 19, 1179–1199. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Cantalapiedra-Navarrete, C.; Castillo, P. Cryptic species in plant-parasitic nematodes. Nematology 2014, 16, 1105–1118. [Google Scholar] [CrossRef]
- Bert, W.; Geraert, E. Nematode species of the order Tylenchida, new to the Belgian nematofauna with additional morphological data. Belg. J. Zool. 2000, 130, 47–57. [Google Scholar]
- Brzeski, M.W. Paratylenchinae: Morphology of some known species and descriptions of Gracilacus bilineata sp. n. and G. vera sp. n. (Nematoda: Tylenchulidae). Nematologica 1995, 41, 535–565. [Google Scholar] [CrossRef]
- Lišková, M.; Sasanelli, N.; D’Addabbo, T. Some notes on the occurrence of plant parasitic nematodes on fruit trees in Slovakia. Plant Prot. Sci. 2007, 43, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Steiner, G. Plant nematodes the grower should know. Proc. Soil Sci. Soc. 1949, 2, 37–39. [Google Scholar]
- Oostenbrink, M. A note on Paratylenchus in the Netherlands with the description of P. goodeyi n. sp. (Nematoda, Criconematidae). Tijdschr. Plantenziekten 1953, 59, 207–216. [Google Scholar] [CrossRef]
- Raski, D.J. Revision of the genus Paratylenchus Micoletzky, 1922 and descriptions of new species. Part II of three parts. J. Nematol. 1975, 7, 274–295. [Google Scholar] [PubMed]
- Andrássy, I. Neue und wenig bekannte nematoden aus Jugoslawien. Ann. Hist. Nat. Musei Natl. Hung. 1959, 51, 259–275. [Google Scholar]
- Cobb, N.A. Notes on Paratylenchus, a genus of nemas. J. Wash. Acad. Sci. 1923, 13, 254–257. [Google Scholar]
- Subbotin, S.A.; Yan, G.; Kantor, M.; Handoo, Z. On the molecular identity of Paratylenchus nanus Cobb, 1923 (Nematoda: Tylenchida). J. Nematol. 2020, accepted. [Google Scholar]
- De Coninck, L. Sur trois espèces nouvelles de nématodes libres trouvés en Belgique. Bul. Mus. Roy. D’hist. Nat. Belg. 1931, 7, 1–5. [Google Scholar]
- Wu, L.Y. Paratylenchus veruculatus n. sp. (Criconematidae: Nematoda) from Scotland. Can. J. Zool. 1962, 40, 773–775. [Google Scholar] [CrossRef]
- Thorne, G.; Allen, M.W. Paratylenchus hamatus n. sp, and Xiphinema index n. sp., two nematodes associated with fig roots, with a note on Paratylenchus anceps Cobb. Proc. Helm. Soc. Wash. 1950, 17, 27–35. [Google Scholar]
- Meldal, B.H.; Debenham, N.J.; De Ley, P.; De Ley, I.T.; Vanfleteren, J.R.; Vierstraete, A.R.; Bert, W.; Borgonie, G.; Moens, T.; Tyler, P.A.; et al. An improved molecular phylogeny of the Nematoda with special emphasis on marine taxa. Mol. Phylogenetics Evol. 2007, 42, 622–636. [Google Scholar] [CrossRef]
- Ortiz, V.; Phelan, S.; Mullins, E. A temporal assessment of nematode community structure and diversity in the rhizosphere of cisgenic Phytophthora infestans-resistant potatoes. BMC Ecol. 2016, 16, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Háněl, L.; Brzeski, M. Paratylenchinae: Evaluation of diagnostic morpho-biometrical characters of females in the genus Paratylenchus Micoletzky, 1922 (Nematoda: Tylenchulidae). Nematology 2000, 2, 253–261. [Google Scholar] [CrossRef]
- Hirschmann, H. Scanning electron microscopy as a tool in nematode taxonomy. In Concepts in Nematode Systematics; Stone, A.R., Platt, H.M., Kahlil, L.F., Eds.; Academic Press: New York, NY, USA, 1983; pp. 95–111. [Google Scholar]
- Wergin, W.P. Scanning electron microscopic techniques and applications for use in nematology. In Plant Parasitic Nematodes; Zuckerman, B.M., Rhode, R.A., Eds.; Academic Press: New York, NY, USA, 1981; Volume 3, pp. 175–204. [Google Scholar]
- Eisenback, J.D. Techniques for preparing nematodes for scanning electron microscopy. In An Advanced Treatise on Meloidogyne; Barker, K.R., Carter, C.C., Sasser, N.S., Eds.; Raleigh, State University Graphics: Raleigh, NC, USA, 1985; Volume 2, pp. 79–105. [Google Scholar]
- Nassonova, E.; Smirnov, A.; Fahrni, J.; Pawlowski, J. Barcoding amoebae: Comparison of SSU, ITS and COI genes as tools for molecular identification of naked lobose amoebae. Protist 2010, 161, 102–115. [Google Scholar] [CrossRef]
- Morise, H.; Miyazaki, E.; Yoshimitsu, S.; Eki, T. Profiling nematode communities in unmanaged flowerbed and agricultural field soils in Japan by DNA barcode sequencing. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Armenteros, M.; Rojas-Corzo, A.; Ruiz-Abierno, A.; Derycke, S.; Backeljau, T.; Decraemer, W. Systematics and DNA barcoding of free-living marine nematodes with emphasis on tropical desmodorids using nuclear SSU rDNA and mitochondrial COI sequences. Nematology 2014, 16, 979–989. [Google Scholar] [CrossRef]
- Kiewnick, S.; Holterman, M.; van den Elsen, S.; van Megen, H.; Frey, J.E.; Helder, J. Comparison of two short DNA barcoding loci (COI and COII) and two longer ribosomal DNA genes (SSU & LSU rRNA) for specimen identification among quarantine root-knot nematodes (Meloidogyne spp.) and their close relatives. Eur. J. Plant Pathol. 2014, 140, 97–110. [Google Scholar]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstens, B.C.; Pelletier, T.A.; Reid, N.M.; Satler, J.D. How to fail at species delimitation. Mol. Ecol. 2013, 22, 4369–4383. [Google Scholar] [CrossRef]
- Leliaert, F.; Verbruggen, H.; Vanormelingen, P.; Steen, F.; López-Bautista, J.M.; Zuccarello, G.C.; De Clerck, O. DNA-based species delimitation in algae. Eur. J. Phycol. 2014, 49, 179–196. [Google Scholar] [CrossRef] [Green Version]
- Janssen, T.; Karssen, G.; Orlando, V.; Subbotin, S.; Bert, W. Molecular characterization and species delimiting of plant parasitic nematode of the genus Pratylenchus from the Penetrans group (Nematoda: Pratylenchidae). Mol. Phylogenet. Evol. 2017, 117, 30–48. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W.; Gamliel, A.; Bucki, P.; Duvrinin, S.; Alon, T.; Miyara, S.B. Phylogeography and molecular species delimitation of Pratylenchus capsici n. sp., a new root-lesion nematode in Israel on pepper (Capsicum annuum). Phytopathology 2018, 109, 847–858. [Google Scholar] [CrossRef]
- Xu, X.; Qing, X.; Xie, J.L.; Yang, F.; Peng, Y.L.; Ji, H.L. Population structure and species delimitation of rice white tip nematode, Aphelenchoides besseyi (Nematoda: Aphelenchoididae), in China. Plant Pathol. 2020, 69, 159–167. [Google Scholar] [CrossRef]
- Padial, J.M.; De la Riva, I. A paradigm shift in our view of species drives current trends in biological classification. Biol. Rev. 2020, 23, 1–21. [Google Scholar]
- Prévot, V.; Jordaens, K.; Sonet, G.; Backeljau, T. Exploring Species Level Taxonomy and Species Delimitation Methods in the Facultatively Self-Fertilizing Land Snail Genus Rumina (Gastropoda: Pulmonata). PLoS ONE 2013, 8, e60736. [Google Scholar]
- Zink, R.M.; Barrowclough, G.F. Mitochondrial DNA under siege in avian phylogeography. Mol. Ecol. 2008, 17, 2107–2121. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Cipriano, F.; Hare, M.P. Predicting nuclear gene coalescence from mitochondrial data: The three-times rule. Evolution 2001, 55, 859–868. [Google Scholar] [CrossRef]
- Jörger, K.M.; Norenburg, J.L.; Wilson, N.G.; Schrödl, M. Barcoding against a paradox? Combined molecular species delineations reveal multiple cryptic lineages in elusive meiofaunal sea slugs. BMC Evol. Biol. 2012, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; Boisselier, M.C.; Cruaud, C.; Holford, M.; Samadi, S. Large-scale species delimitation method for hyperdiverse groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 1–16. [Google Scholar] [CrossRef]
- Whitehead, A.G.; Hemming, J.R. A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol. 1965, 55, 25–38. [Google Scholar] [CrossRef]
- Jenkins, W.R.B. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Dis. Rep. 1964, 48, 692. [Google Scholar]
- Available online: https://www.youtube.com/watch?v=qWITH1yB1U0&t=2s (accessed on 15 February 2021).
- Singh, P.R.; Nyiragatare, A.; Janssen, T.; Couvreur, M.; Decraemer, W.; Bert, W. Morphological and molecular characterisation of Pratylenchus rwandae n. sp. (Tylenchida: Pratylenchidae) associated with maize in Rwanda. Nematology 2018, 20, 781–794. [Google Scholar] [CrossRef]
- Nunn, G.B. Nematode Molecular Evolution. An Investigation of Evolutionary Patterns among Nematodes Based Upon DNA Sequences. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1992. [Google Scholar]
- Vrain, T.C.; Wakarchuk, D.A.; Levesque, A.C.; Hamilton, R.I. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam. Appl. Nematol. 1992, 15, 563–573. [Google Scholar]
- Curran, J.; Driver, F.; Ballard, J.W.O.; Milner, R.J. Phylogeny of Metarhizium: Analysis of ribosomal DNA sequence data. Mycol. Res. 1994, 98, 547–552. [Google Scholar] [CrossRef]
- Mayer, W.E.; Herrmann, M.; Sommer, R.J. Phylogeny of the nematode genus Pristionchus and implications for biodiversity, biogeography and the evolution of hermaphroditism. BMC Evol. Biol. 2007, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.R.; Couvreur, M.; Decraemer, W.; Bert, W. Survey of slug-parasitic nematodes in East and West Flanders, Belgium and description of Angiostoma gandavensis n. sp. (Nematoda: Angiostomidae) from arionid slugs. J. Helminthol. 2019, 94, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tanha Maafi, Z.; Subbotin, S.A.; Moens, M. Molecular identification of cyst-forming nematodes (Heteroderidae) from Iran and a phylogeny based on the ITS sequences of rDNA. Nematology 2003, 5, 99–111. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Powers, T.O.; Bernard, E.C.; Harris, T.; Higgins, R.; Olson, M.; Lodema, M.; Mullin, P.; Sutton, L.; Powers, K.S. COI haplotype groups in Mesocriconema (Nematoda: Criconematidae) and their morphospecies associations. Zootaxa 2014, 3827, 101–146. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.R.; Karssen, G.; Couvreur, M.; Bert, W. Morphological and molecular characterization of Heterodera dunensis n. sp.(Nematoda: Heteroderidae) from Gran Canaria, Canary Islands. J. Nematol. 2020, 52, 1–14. [Google Scholar]
- Available online: https://dna.macrogen.com (accessed on 30 December 2020).
- Available online: https://www.genewiz.com (accessed on 15 December 2020).
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Subbotin, S.A.; Sturhan, D.; Adams, B.J.; Powers, T.O.; Mullin, P.G.; Chizhov, V.N.; Vovlas, N.; Baldwin, J.G. Molecular phylogeny of the order tylenchida: Analysis of nuclear ribosomal RNA genes. J. Nematol. 2006, 38, 296. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L.; Sullivan, J. Phylogeny inference based on parsimony and other methods using PAUP* Practice. In The Phylogenetic Handbook, a Practical Approach to DNA and Protein Phylogeny; Salemi, M., Vandamme, A.M., Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 182–206. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://species.h-its.org (accessed on 10 December 2020).
- Available online: https://species.h-its.org/gmyc (accessed on 10 December 2020).




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Associated Plant Host | Sample Code | GenBank Accession Numbers | Source | |||
|---|---|---|---|---|---|---|---|---|
| 28S rRNA | ITS rRNA | 18S rRNA | COI of mtDNA | |||||
| P. aculentus | Belgium, Ghent, Citadel Park; 51°02′05″ N; 3°43′10″ E | Grasses under a tree | BE9 | MW413626–MW41328 | MW413588–MW413589 | MW413692–MW413693 | MW421639–MW421642 | C.M. Etongwe |
| P. aquaticus type A | USA, Florida, Princeton | Aechmea sp. | CD3375 | MW413557 | - | - | - | S.A. Subbotin |
| P. aquaticus type A | USA, Hawaii, Waimanalo | Bromeliad (Neoregelia sp.) | CD619 | KF242239, KF242240 | KF242277, KF242278 | - | MW411845 | S.A. Subbotin, Van den Berg et al. [22] |
| P. aquaticus type B | USA, Kansas, Manhattan, WashintonMarlatt park | Grasses | CD868 | KF242241 | - | - | MW411838 | S.A. Subbotin, Van den Berg et al. [22] |
| P. dianthus | South Africa, Gauteng, Tarlton | Chrysanthemum | CD552 | KF242226–KF242229 | KF242271, KF242272 | - | MW411837 | Van den Berg, Van den Berg et al. [22] |
| P. enigmaticus | Belgium, Ghent, Ghent University Botanical Garden; 51°2′7.53″ N; 3°43′20.07″ E | Leek | BE2 | - | MW413621–MW413622 | MW413735, MW413737–MW413739 | MW421686 | C.M. Etongwe |
| P. enigmaticus | Belgium, Ghent, Ghent University Botanical Garden; 51°2′7.10″ N; 3°43′19.28″ E | Wild oregano | BE4 | - | - | MW413732–MW413734 | - | C.M. Etongwe |
| P. enigmaticus | USA, Idaho | Unknown plant | CD2485 | MW413568 | MW413583 | - | MW411828 | S.A. Subbotin |
| P. elachistus | Belgium, Kortrijk; 50°47′58″ N; 3°11′37″ E | Grasses under a thorny tree | BE15 | MW413629–MW413630 | MW413590–MW413593 | MW413694–MW413697 | MW421643–MW421646 | C.M. Etongwe |
| P. goodeyi | Belgium, Merendree; 51°04′12″ N; 3°34′37″ E | Grasses around a beech tree | BE22 | MW413631–MW413633 | MW413594 | MW413698–MW413699 | MW421647–MW421649 | C.M. Etongwe |
| P. hamatus | USA, California, Merced County, Planada | Fig tree (Ficus carica) | CD1155 | KF242212 | - | - | MW411821 | S.A. Subbotin, Van den Berg et al. [22] |
| P. hamatus | USA, California | Trees | CD1914 | MW413564 | MW413585 | - | MW411823 | S.A. Subbotin |
| P. hamatus | USA, California, Kern county | Grape (Vitis sp.) | CD2534a, b | MW413565, MW413566 | - | - | - | A. Westphal |
| P. hamatus | USA, California, Kern county, Delano | Grape, Cherry | CD3372 | MW413558 | - | - | - | S.A. Subbotin |
| P. hamatus | USA, California, Kern county, Maricopa | Apricot (Prunus sp.) | CD454 | KF242206, KF242216, KF242217 | KF242247, KF242256 | - | MW411822 | S.A. Subbotin, Van den Berg et al. [22] |
| P. holdemani | Belgium, Gouvy, Rogery; 50°14′39.8″ N; 5°57′21.9″ E | Grasses under tree Fraxinus sp. | AR3 | MW413636–MW413638, MW413640, MW413642 | - | MW413701 | MW421650–MW421652 | P.R. Singh |
| P. holdemani | Belgium, Ghent, Blaarmeersen; 51°02′18.9″ N; 3°41′17.2″ E | Grasses under a thorny tree next to a stream | BE19 | MW413634–MW413635 | MW413595 | MW413700 | MW421658 | C.M. Etongwe |
| P. holdemani | Belgium, Ghent, Blaarmeersen; 51°02′14″ N; 3°41′23″ E | Grasses under a tree | BE20 | MW413639, MW413641 | MW413596 | MW413702 | MW421653–MW421657 | C.M. Etongwe |
| P. idalimus | Belgium, Ghent, Blaarmeersen; 51°02′18.9″ N; 3°41′17.2″ E | Grasses under a thorny tree next to a stream | BE19 | MW413644 | - | - | - | C.M. Etongwe |
| P. idalimus | Belgium, Ghent, Blaarmeersen; 51°02′14″ N; 3°41′23″ E | Grassesundera tree | BE20 | MW413643 | - | MW413703–MW413704 | - | C.M. Etongwe |
| P. idalimus | USA, California, Napa county, Napa | Grape (Vitis sp.) | CD106 | KF242237, KF242238 | KF242275, KF242276 | - | MW411839 | S.A. Subbotin, Van den Berg et al. [22] |
| P. leptos | Ethiopia, Jimma Zone, Gera district | Coffee | Ge16c | MW413645–MW413653 | - | - | MW421659–MW421665 | A.W. Aseffa |
| P. microdorus | Belgium, Zwijnaarde; 51°00′19″ N; 3°42′11″ E | Grasses | BE11 | - | MW413597 | - | - | C.M. Etongwe |
| P. microdorus | Belgium, Ghent, Citadel Park; 51°02′05″ N; 3°43′10″ E | Grasses under a tree | BE9 | MW413654–MW413655 | MW413598–MW413600 | MW413705–MW413706 | MW421666–MW421667 | C.M. Etongwe |
| P. nanus | USA, California, Humboldt county, Trinidad, sample 5B; 41°02′40.6″ N; 124°07′18.1″ W | Unknown plant | CD3141 | MW413576 | - | - | - | S.A. Subbotin |
| P. nanus | USA, Washington, Mason County, Skokomish, sample 32B; 47°18′07.0″ N; 123°10′95.6″ W | Unknown plant | CD3217 | MW413575 | - | - | - | S.A. Subbotin |
| P. nanus | USA, California, Riverside | Grasses | CD728 | KF242194, KF242197 | KF242267, KF242268 | - | MW411835 | S.A. Subbotin, Van den Berg et al. [22] |
| P. nanus | USA, California, Marin county | Festuca sp. | CD850 | KF242192, KF242193 | - | - | MW411834 | S.A. Subbotin, Van den Berg et al. [22] |
| P. nanus | USA, California, Marin county | Grasses | CD860 | KF242191, KF242195 | - | - | MW411836 | S.A. Subbotin, Van den Berg et al. [22] |
| P. nanus | Belgium, Gouvy, Rogery;50°14′39.8″ N; 5°57′21.9″ E | Grasses under tree Fraxinus sp. | AR3 | - | - | - | MW421673 | P.R. Singh |
| P. nanus | Belgium, Zwijnaarde; 51°00′19″ N; 3°42′11″ E | Grasses under a tree | BE11 | MW413658–MW413659 | MW413601–MW413603 | MW413707, MW413711–MW413712 | MW421668–MW421671, MW421674 | C.M. Etongwe |
| P. nanus | Belgium, Ghent, Blaarmeersen; 51°07′14″ N; 2°39′29″ E | Grasses under a tree | BE18 | MW413657 | MW413604 | MW413708 | MW421672 | C.M. Etongwe |
| P. neoamblycephalus | Belgium, Ghent, Citadel Park; 51°02′09″ N; 3°43′06″ E | Cypress tree | BE10 | MW413660–MW413663 | MW413606–MW413610 | MW413713–MW413718 | MW421675–MW421682 | C.M. Etongwe |
| P. neoamblycephalus | USA, California, Madera county, Madera | Grasses | CD1223 | KF242190 | - | - | MW411843 | S.A. Subbotin, Van den Berg et al. [22] |
| P. projectus | Belgium, Ghent, Ghent University Botanical Garden; 51°2′7.53″ N; 3°43′20.07″ E | Leek | BE2 | MW413656 | MW413605 | MW413709–MW413710 | - | C.M. Etongwe |
| P. projectus | USA, California, Butte county, Gridley | Walnut (Juglans sp.) | CD137 | KF242199 | KF242265, KF242266 | - | MW411840 | S.A. Subbotin, Van den Berg et al. [22] |
| P. projectus | South Africa, Western Cape, George | Bent grass | CD587 | KF242198, KF242200 | KF242263, KF242264 | - | MW411842 | Van den Berg, Van den Berg et al. [22] |
| P. shenzhenensis | USA, Florida, Apopka | Unknown plant | CD2728 | MW413579 | - | - | - | S.A. Subbotin |
| P. straeleni | Belgium, Zwijnaarde; 51°00′19″ N; 3°42′11″ E | Grasses under a tree | BE11 | MW413686 | MW413623 | - | MW421713–MW421715 | C.M. Etongwe |
| P. straeleni | Belgium, Kortrijk; 50°47′58″ N; 3°11′37″ E | Grasses under a thorny tree | BE15 | MW413685 | MW413624–MW413625 | MW413743–MW413746 | MW421708–MW421712 | C.M. Etongwe |
| P. straeleni | Belgium, Ghent, Blaarmeersen; 51°02′18.9″ N; 3°41′17.2″ E | Grasses under a thorny tree next to a stream | BE19 | - | - | - | MW421716 | C.M. Etongwe |
| P. straeleni | USA, North Carolina | Unknown plant | CD1433 | MW413577 | - | - | - | W. Ye |
| P. straeleni | USA, California, Monterey county | Oak | CD1775 | MW413578 | - | - | MW411831 | S.A. Subbotin |
| P. straeleni | USA, California, Napa county | Tree | CD899 | KF242236 | - | - | MW411832 | S.A. Subbotin, Van den Berg et al. [22] |
| P. tenuicaudatus | USA, California, Glenn county, Orland | Prune (Prunus sp.) | CD57 | KF242223, KF242225 | KF242261, KF242262 | - | MW411826 | S.A. Subbotin, Van den Berg et al. [22] |
| P. tenuicaudatus | USA, California, Glenn county, Butte City | Prune (Prunus sp.) | CD61 | KF242224 | KF242259, KF242260 | - | MW411827 | S.A. Subbotin, Van den Berg et al. [22] |
| P. veruculatus | Belgium, Ghent, Blaarmeersen; 51°02′14″ N; 3°41′23″ E | Grasses under a tree | BE20 | MW413687–MW413691 | - | MW413747–MW413748 | MW421717–MW421722 | C.M. Etongwe |
| Paratylenchus sp.2 | Belgium, Kortrijk; 50°47′58″ N; 3°11′37″ E | Grasses under a thorny tree | BE15 | MW413670–MW413671 | MW413615–MW413616 | MW413724–MW413725 | MW421683–MW421684 | C.M. Etongwe |
| Paratylenchus sp.2 | Belgium, Ghent, Blaarmeersen; 51°02′14″ N; 3°41′23″ E | Grasses under a tree | BE20 | - | - | MW413726 | MW421685 | C.M. Etongwe |
| Paratylenchus sp.2 | Kyrgyzstan | Trees and grasses | CD2139 | MW413567 | - | - | - | S.A. Subbotin |
| Paratylenchus sp.2 | USA, California, Yolo county, Davis | Grasses under a willow tree | CD604 | KF242220–KF242222 | KF242243 | - | MW411825 | S.A. Subbotin, Van den Berg et al. [22] |
| Paratylenchus sp.3 | USA, California, Santa Barbara county, Goleta | Lemon (Citrus sp.) | CD232 | KF242231, KF242232 | KF242273, KF242274 | - | MW411819 | S.A. Subbotin, Van den Berg et al. [22] |
| Paratylenchus sp.3 | USA, Florida | Unknown plant | CD2726 | MW413573 | - | - | MW411820 | S.A. Subbotin |
| Paratylenchus sp.4 | USA, Oregon | Trees | CD986 | KF242203 | - | - | MW411829 | S.A. Subbotin, Van den Berg et al. [22] |
| Paratylenchus sp.7 | USA, California, Riverside, UCR campus | Unknown plant | CD1004 | KF242242 | - | - | MW411830 | S.A. Subbotin |
| Paratylenchus sp.AH | USA, California, El Dorado county, Placerville, Apple hills, sample N12 | Unknown plant | CD1692 | MW420921 | - | - | MW411844 | S.A. Subbotin |
| Paratylenchus sp.BE11 | Belgium, Zwijnaarde; 51°00′19″ N; 3°42′11″ E | Grasses under a tree | BE11 | MW413672–MW413674 | MW413617 | - | MW421687–MW421688 | C.M. Etongwe |
| Paratylenchus sp.CaD | USA, California, El Dorado county, Placerville, Apple hills, sample, N7 | Unknown plant | CD1686 | MW413561 | - | - | MW411841 | S.A. Subbotin |
| Paratylenchus sp.CaD | USA, California, El Dorado county, Placerville, Apple hills, sample N16 | Unknown plant | CD1695a, b | MW413560, MW413562 | MW413584 | - | MW411824 | S.A. Subbotin |
| Paratylenchus sp.CaD | USA, California, El Dorado county, Placerville, Apple hills, sample N20 | Unknown plant | CD1696 | MW413563 | - | - | - | S.A. Subbotin |
| Paratylenchus sp.CaD | USA, California, Yolo county, Putah Creek | Rubus sp. | CD1791 | MW413559 | - | - | - | S.A. Subbotin |
| Paratylenchus sp.D | Belgium, Ghent, Blaarmeersen; 51°02′14″ N; 3°41′23″ E | Grasses under a tree | BE20 | MW413664–MW413669 | MW413611–MW413614 | MW413719–MW413723 | MW421689–MW421699 | C.M. Etongwe |
| Paratylenchus sp.Dia | USA, California, Contra Costa County, Mount Diablo State Park | Unknown plant | CD1776 | MW413574 | - | - | MW411833 | S.A. Subbotin |
| Paratylenchus sp.F | Belgium, Ghent, Blaarmeersen; 51°07′14″ N; 2°39′29″ E | Grasses under a tree | BE18 | - | - | MW413728 | - | C.M. Etongwe |
| Paratylenchus sp.F | Belgium, Merendree; 51°04′12″ N; 3°34′37″ E | Grasses around a beech tree | BE22 | MW413675–MW413679 | MW413618–MW413620 | MW413727, MW413729–MW413731 | MW421700–MW421702 | C.M. Etongwe |
| Paratylenchus sp.F | Russia, Primorsky Krai, Olginsky district | Unknown plant | CD1842, CD1844 | MW413571, MW413572 | - | - | J. Zograf | |
| Paratylenchus sp.Ge16 | Ethiopia, Jimma Zone, Gera district | Coffee | Ge16c | MW413680–MW413682 | - | MW413732–MW413734 | MW421703–MW421705 | C.M. Etongwe |
| Paratylenchus sp.J | USA, Washington, Mason County, Skokomish, sample 32E; 47°18′07.0″ N 123°10′95.6″ W | Unknown plant | CD3216 | MW413570 | - | - | - | S.A. Subbotin |
| Paratylenchus sp.J | USA, Oregon, Douglas County, Oakland, sample 35; 43°28′59.9″ N 123°19′24.5″ W | Unknown plant | CD3220 | MW413569 | - | - | - | S.A. Subbotin |
| Paratylenchus sp.NL | The Netherlands, Hilversum | Holly | NL | MW413683–MW413684 | - | MW413740–MW413742 | MW421706–MW421707 | G. Karssen |
| Paratylenchus sp.SK | South Korea | Pinus sp. | CD1384 | MW413580 | - | - | S.A. Subbotin | |
| Population | P. aculentus (BE9) | P. goodeyi (BE22) | P. idalimus (BE19 and BE20) | P. straeleni (BE15) |
|---|---|---|---|---|
| n | 12 | 17 | 7 | 11 |
| L | 266 ± 20.1 (233–03) | 348 ± 42.5 (266–452) | 299 ± 20.7 (278–332) | 358 ± 13.1 (330–379) |
| a | 19.6 ± 2.0 (16.3–23.2) | 20.7 ± 1.7 (16.7–23.2) | 21.0 ± 1.3 (20–23) | 22.6 ± 0.9 (20.7–24.3) |
| b | 2.6 ± 0.1 (2.4–2.8) | 3.2 ± 0.3 (2.9–3.7) | 2 ± 0.2 (2.0–2.4) | 3.6 ± 0.1 (3.4–3.7) |
| c | 12.4 ± 1.5 (10.8–15.2) | 11.9 ± 1.4 (10.1–13.5) | 12.1 ± 0.6 (12.1–13.1) | 10.6 ± 1.0 (8.9–11.8) |
| c | 2.8 ± 0.3 (2.4–3.1) | 3.0 ± 0.2 (2.8–3.3) | 4.0 ± 0.6 (3.1–4.1) | 3.4 ± 0.3 (3.0–3.9) |
| Maximum body width | 13.6 ± 1.3 (11.6–15.5) | 17.0 ± 3.3 (13.0–27.0) | 14.0 ± 1.4 (13.0–16.1) | 15.9 ± 0.7 (14.6–16.7) |
| Stylet length | 56.0 ± 3.3 (52.4–61.2) | 52.1 ± 2.8 (47.0–58.6) | 89.0 ± 3.5 (84.1–93.0) | 55.7 ± 1.7 (53.5–58.6) |
| Cone length | 49.1 ± 3.6 (43.0–54.9) | 43.0 ± 2.7 (48.2–48.5) | 78.0 ± 2.9 (74.0–83.1) | 44.7 ± 1.7 (42.2–47.4) |
| Cone%stylet | 87.5 ± 3.8 (80.1–91.0) | 82.4 ± 2.5 (78.0–89.6) | 88.0 ± 2.2 (83–89) | 80 ± 1.8 (76–83) |
| Knob width | 3.2 ± 0.5 (2.3–4.0) | 4.1 ± 0.6 (3.3–5.2) | 4.0 ± 0.2 (4.0–4.3) | 3.9 ± 0.4 (3.1–4.6) |
| Pharynx length | 101 ± 8.3 (87.0–113) | 109 ± 11.7 (92.7–133) | 130 ± 12.8 (114–147) | 100 ± 3.8 (92.1–105) |
| Anterior end to SE pore | 66.7 ± 5.2 (54.3–74.4) | 80.7 ± 8.6 (68.5–99.0) | 93.0 ± 11.7 (82.0–115) | 82.5 ± 2.9 (79.4–87.6) |
| SE pore%L | 25.2 ± 0.9 (23.3–26.4) | 22.9 ± 1.4 (21.0–25.8) | 31 ± 4.2 (28–40) | 23 ± 0.7 (22–24) |
| Anterior end to vulva | 193 ± 16.1 (165–218) | 279 ± 31.9 (216–356) | 233 ± 17.9 (214–260) | 270 ± 18.8 (249–330) |
| V% | 72.5 ± 1.5 (70.8–75.7) | 80.1 ± 1.4 (77.8–82.3) | 78.0 ± 0.9 (77–79) | 81 ± 1.9 (80–84) |
| Body width at anus | 7.6 ± 0.5 (7.0–8.3) | 9.7 ± 0.5 (9.7–10.0) | 7.0 ± 0.1 (7.0–7.1) | 10.1 ± 0.4 (9.1–10.5) |
| Tail length | 20.9 ± 2.3 (18.1–25.1) | 29.0 ± 2.4 (25.8–32.1) | 25.0 ± 2.2 (22.0–28.1) | 34.4 ± 3.7 (31–40.8) |
| Population | P. elachistus (BE15) | P. holdemani (AR3) | P. microdorus (BE9) | P. veruculatus (BE20) |
|---|---|---|---|---|
| n | 24 | 31 | 10 | 15 |
| L | 301 ± 12.5 (283–329) | 359 ± 47 (285–475) | 330.7 ± 20 (297–355) | 286 ± 24.7 (251–331) |
| a | 20.4 ± 1.1 (17.7–22.6) | 20.9 ± 1.9 (16.4–25.2) | 21.4 ± 1.4 (18.7–23.1) | 19.8 ± 1.9 (17.2–23.3) |
| b | 4.1 ± 0.2 (3.8–4.3) | 4.1 ± 0.7 (2.2–5.1) | 5.0 ± 0.5 (4.5–6.2) | 3.8 ± 0.3 (3.3–4.2) |
| c | 12.1 ± 0.9 (10.9–14.3) | 14.8 ± 1.4 (12.4–17.7) | 10.6 ± 0.9 (9.2–12) | 17.8 ± 1.8 (14.6–20.6) |
| c’ | 2.8 ± 0.2 (2.4–3.3) | 2.5 ± 0.3 (2.1–3.2) | 3.8 ± 0.5 (2.8–4.5) | 2.2 ± 0.2 (1.8–2.6) |
| Maximum body width | 14.8 ± 1.0 (12.7–16.6) | 17.3 ± 3.0 (11.3–23.8) | 15.5 ± 1.4 (13.3–17.1) | 14.5 ± 1.4 (12.5–16.5) |
| Stylet length | 20.9 ± 0.7 (19.7–22.2) | 22.5 ± 2.0 (19.0–26.1) | 12.4 ± 1.3 (10.6–14.7) | 14.2 ± 0.5 (13.1–14.8) |
| Cone length | 13.3 ± 0.4 (12.4–13.9) | 15.1 ± 1.1 (13.2–18.5) | 6.7 ± 1.3 (4.8–8.1) | 8.9 ± 0.3 (8.3–9.3) |
| Cone%stylet | 63.7 ± 1.5 (60.5–67.5) | 67.3 ± 3.5 (60.9–77.4) | 53.4 ± 5.7 (45.3–60.4) | 62.8 ± 1.1 (60.3–64.8) |
| Knob width | 3.5 ± 0.2 (3.1–4.1) | 3.3 ± 0.4 (2.9–4.2) | - | 3.1 ± 0.3 (2.7–3.5) |
| Pharynx length | 74.5 ± 2.6 (70.4–80.6) | 89.7 ± 21.5 (66.1–161) | 66.6 ± 6.4 (56.2–76.1) | 75.8 ± 7.0 (60.8–88.4) |
| Anterior end to SE pore | 60.7 ± 3.9 (54.0–68.5) | 74.8 ± 9.1 (60.1–99.0) | 63.6 ± 4.7 (57.5–71.3) | 62.3 ± 6.5 (51.2–74.4) |
| SE pore%L | 20.1 ± 0.9 (18.6–21.9) | 21.2 ± 1.8 (16.4–23.7) | 19.2 ± 1.1 (17.3–20.6) | 21.8 ± 2.0 (17.5–25.6) |
| Anterior end to vulva | 245 ± 10.2 (226–269) | 303 ± 40.9 (238–391) | - | 245 ± 22.0 (215–284) |
| V% | 81.3 ± 0.9 (79.7–83.2) | 84.3 ± 1.8 (81.3–90.5) | 81 ± 1.6 (79.1–82.8) | 85.7 ± 1.4 (83.8–89.7) |
| Body width at anus | 8.9 ± 0.7 (7.5–9.9) | 10.0 ± 1.3 (7.2–12.3) | 8.5 ± 1.3 (6.8–10.8) | 7.3 ± 0.6 (6.3–8.8) |
| Tail length | 24.8 ± 2.2 (20.9–29.1) | 25.2 ± 2.8 (20.0–29.5) | 31.8 ± 3.1 (28.1–35.9) | 16.1 ± 1.7 (13.6–19.1) |
| Population | P. nanus (BE11) | P. neoamblycephalus (BE10) | Paratylenchus sp.2 (BE15) | Paratylenchus sp.D (BE20) | Paratylenchus sp.F (BE22) |
|---|---|---|---|---|---|
| n | 30 | 15 | 16 | 11 | 17 |
| L | 318 ± 15.8 (287–352) | 337 ± 20.2 (301–367) | 347 ± 20.7 (308–389) | 328 ± 36.1 (285–387) | 300 ± 21.1 (264–339) |
| a | 18.1 ± 1.3 (15.6–20.4) | 18.4 ± 1.0 (16.8–19.9) | 23.2 ± 1.8 (20.1–28.7) | 21.5 ± 1.6 (19.1–24.4) | 20.3 ± 1.3 (18.4–23.0) |
| b | 3.9 ± 0.3 (3.5–4.8) | 4.3 ± 0.5 (3.4–4.9) | 3.9 ± 0.3 (3.5–4.4) | 3.6 ± 0.3 (3.3–4.1) | 4.0 ± 0.3 (3.7–4.6) |
| c | 14.8 ± 1.3 (12.8–16.8) | 14.6 ± 1.5 (12.9–16.6) | 13.2 ± 0.7 (12.3–14.4) | 14.5 ± 1.4 (12.0–15.8) | 12.9 ± 0.8 (11.8–14.0) |
| c’ | 2.0 ± 0.2 (1.7–2.6) | 2.2 ± 0.2 (1.9–2.4) | 3.1 ± 0.2 (2.9–3.5) | 2.7 ± 0.3 (2.5–3.2) | 2.6 ± 0.2 (2.3–2.9) |
| Max. body width | 17.6 ± 1.2 (15.4–20.5) | 18.3 ± 1.4 (16.8–20.7) | 15.1 ± 1.3 (13.2–16.8) | 15.3 ± 2.2 (13.2–19.9) | 14.8 ± 1.1 (13.3–16.6) |
| Stylet length | 28.8 ± 1.2 (26.7–31.2) | 33.4 ± 0.9 (32.0–34.3) | 28.4 ± 1.5 (26.5–31.4) | 27.5 ± 1.0 (25.7–28.9) | 27.6 ± 1.2 (25.3–29.6) |
| Cone length | 20.2 ± 1.4 (17.7–23.4) | 22.6 ± 0.9 (21.4–24.5) | 19.2 ± 1.0 (17.5–20.8) | 17.7 ± 0.8 (17.0–19.2) | 18.5 ± 0.9 (17.1–20.3) |
| Cone%stylet | 70.1 ± 3.6 (64.9–78.5) | 67.8 ± 2.8 (63.0–72.5) | 67.5 ± 2.0 (64.4–71.2) | 64.3 ± 2.0 (60.7–67.3) | 67.1 ± 1.3 (65.2–69.5) |
| Knob width | 3.9 ± 0.4 (3.1–4.6) | 4.8 ± 0.2 (4.4–5.1) | 4.1 ± 0.3 (3.5–4.7) | 4.0 ± 0.3 (3.5–4.6) | 3.6 ± 0.3 (3.3–4.1) |
| Pharynx length | 81.6 ± 5.4 (65.4–91.1) | 79.7 ± 9.1 (65.7–93.8) | 88.2 ± 4.1 (78.0–96.7) | 89.6 ± 7.3 (76.0–104) | 74.8 ± 4.9 (67.7–83.1) |
| Ant. end to SE pore | 65.7 ± 6.0 (54.7–75.0) | 63.9 ± 4.7 (52.2–70.0) | 72.3 ± 4.5 (64.2–90.0) | 74.6 ± 6.8 (66.7–90.2) | 63.0 ± 5.4 (51.5–70.6) |
| SE pore%L | 20.6 ± 1.4 (17.0–22.5) | 19.0 ± 1.5 (15.8–21.7) | 20.8 ± 1.3 (19.4–23.5) | 23.6 ± 0.9 (22.1–24.8) | 21.0 ± 1.6 (18.6–24.2) |
| Ant. end to vulva | 270 ± 18.8 (249–330) | 276 ± 15.0 (247–296) | 286 ± 17.1 (252–313) | 270 ± 27.9 (239–320) | 217 ± 17.2 (217–278) |
| V% | 83.8 ± 1.1 (81.7–85.7) | 81.9 ± 0.9 (80.7–83.8) | 82.2 ± 0.8 (81.2–83.5) | 83.7 ± 1.0 (81.8–85.2) | 82 ± 0.7 (80.9–83.5) |
| Body width at anus | 10.9 ± 0.9 (9.3–12.6) | 10.6 ± 0.8 (9.4–12.0) | 8.6 ± 0.7 (7.6–9.8) | 8.0 ± 0.6 (7.3–8.7) | 9.0 ± 0.6 (8.0–9.9) |
| Tail length | 21.7 ± 1.9 (18.9–25.5) | 23.2 ± 2.6 (20.2–27.8) | 26.1 ± 2.0 (23.0–28.7) | 22.0 ± 3.0 (18.0–25.6) | 23.1 ± 2.4 (20.0–26.5) |
| Gene | GenBank Accession No. | Linked Species | Country of Origin | Reference | Reassigned Species Name |
|---|---|---|---|---|---|
| D2-D3 | MN437514 | Gracilacus sp. | Myanmar | Du, Y. (Unpublished) | P. sinensis |
| D2-D3 | AY780943 | P. bukowinensis | Italy | Subbotin et al. [37] | P. holdemani |
| D2-D3 | MN088372 | P. bukowinensis | Iran | Mirbabaei et al. [46] | P. holdemani |
| D2-D3 | MN783703 | P. bukowinensis | Belgium | Etongwe et al. [47] | P. holdemani |
| D2-D3 | AY780944 | Paratylenchus sp. | Italy | Subbotin et al. [37] | P. tenuicaudatus |
| D2-D3 | MH156807 | Paratylenchus sp. | China | Fan et al. (Unpublished) | P. lepidus |
| D2-D3 | KF242223 | Paratylenchus sp.1 | USA | Van den Berg et al. [22] | P. tenuicaudatus |
| D2-D3 | KF242224 | Paratylenchus sp.1 | USA | Van den Berg et al. [22] | P. tenuicaudatus |
| D2-D3 | KF242225 | Paratylenchus sp.1 | USA | Van den Berg et al. [22] | P. tenuicaudatus |
| D2-D3 | KF242237 | Paratylenchus sp.5 | USA | Van den Berg et al. [22] | P. idalimus |
| D2-D3 | KF242238 | Paratylenchus sp.5 | USA | Van den Berg et al. [22] | P. idalimus |
| D2-D3 | KT258978 | Paratylenchus sp. | China | Liu et al. (Unpublished) | P. minor |
| D2-D3 | KF242189 | Paratylenchus sp.6 | USA | Van den Berg et al. [22] | P. neoamblycephalus |
| D2-D3 | KF242190 | Paratylenchus sp.6 | USA | Van den Berg et al. [22] | P. neoamblycephalus |
| D2-D3 | KF242233 | Paratylenchus sp.8 | USA | Van den Berg et al. [22] | P. straeleni |
| D2-D3 | KF242234 | Paratylenchus sp.8 | USA | Van den Berg et al. [22] | P. straeleni |
| D2-D3 | MN783711 | Paratylenchus sp.8 | Belgium | Etongwe et al. [47] | P. straeleni |
| D2-D3 | MN783712 | Paratylenchus sp.E | Belgium | Etongwe et al. [47] | P. microdorus |
| ITS | KT258979 | Paratylenchus sp. | China | Liu et al. (Unpublished) | P. minor |
| ITS | KF242260 | Paratylenchus sp.1 | USA | Van den Berg et al. [22] | P. tenuicaudatus |
| ITS | KF242259 | Paratylenchus sp.1 | USA | Van den Berg et al. [22] | P. tenuicaudatus |
| 18S | AJ966496 | P. dianthus | Belgium | Meldal et al. [65] | P. enigmaticus |
| 18S | KJ636435 | P. nanus | The Netherlands | Van Megen et al. (Unpublished) | Paratylenchus sp.F |
| 18S | KY119503 | P. nanus | Ireland | Ortiz et al. [66] | Paratylenchus sp. |
| COI | MF770960 | Gracilacus sp. | USA | Munawar et al. (Unpublished) | P. straeleni |
| COI | MN710983 | Gracilacus sp. | USA | Powers et al. [49] | P. straeleni |
| COI | MN710984 | Gracilacus sp. | USA | Powers et al. [49] | P. straeleni |
| COI | MN711354 | Paratylenchus sp. | USA | Powers et al. [49] | P. straeleni |
| COI | MN711355 | Paratylenchus sp. | USA | Powers et al. [49] | P. hamatus |
| COI | MN711356 | Paratylenchus sp. | USA | Powers et al. [49] | P. hamatus |
| COI | MN711357 | Paratylenchus sp. | USA | Powers et al. [49] | P. hamatus |
| COI | MN711358 | Paratylenchus sp. | Canada | Powers et al. [49] | P. straeleni |
| COI | MN711359 | Paratylenchus sp. | Canada | Powers et al. [49] | P. straeleni |
| COI | MN711360 | Paratylenchus sp. | Canada | Powers et al. [49] | P. straeleni |
| COI | MN711363 | Paratylenchus sp. | USA | Powers et al. [49] | P. straeleni |
| COI | MN711367 | Paratylenchus sp. | Ireland | Powers et al. [49] | P. straeleni |
| COI | MN711368 | Paratylenchus sp. | Ireland | Powers et al. [49] | P. straeleni |
| COI | MN711369 | Paratylenchus sp. | Ireland | Powers et al. [49] | P. straeleni |
| COI | MN711374 | Paratylenchus sp. | USA | Powers et al. [49] | P. straeleni |
| COI | MN711375 | Paratylenchus sp. | Canada | Powers et al. [49] | P. straeleni |
| COI | MN711376 | Paratylenchus sp. | Canada | Powers et al. [49] | P. straeleni |
| COI | MN711378 | Paratylenchus sp. | Poland | Powers et al. [49] | P. holdemani |
| COI | MN711380 | Paratylenchus sp. | Canada | Powers et al. [49] | P. enigmaticus |
| COI | MN711364 | Paratylenchus sp. | Ireland | Powers et al. [49] | P. nanus |
| COI | MN711365 | Paratylenchus sp. | Ireland | Powers et al. [49] | P. nanus |
| COI | MN782401 | Paratylenchus sp.8 | Belgium | Etongwe et al. [47] | P. straeleni |
| COI | MN782404 | Paratylenchus sp.B | Belgium | Etongwe et al. [47] | P. holdemani |
| COI | MN782405 | Paratylenchus sp.B | Belgium | Etongwe et al. [47] | Paratylenchus sp.D |
| COI | MN782406 | Paratylenchus sp.E | Belgium | Etongwe et al. [47] | P. microdorus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.R.; Karssen, G.; Couvreur, M.; Subbotin, S.A.; Bert, W. Integrative Taxonomy and Molecular Phylogeny of the Plant-Parasitic Nematode Genus Paratylenchus (Nematoda: Paratylenchinae): Linking Species with Molecular Barcodes. Plants 2021, 10, 408. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020408
Singh PR, Karssen G, Couvreur M, Subbotin SA, Bert W. Integrative Taxonomy and Molecular Phylogeny of the Plant-Parasitic Nematode Genus Paratylenchus (Nematoda: Paratylenchinae): Linking Species with Molecular Barcodes. Plants. 2021; 10(2):408. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020408
Chicago/Turabian StyleSingh, Phougeishangbam Rolish, Gerrit Karssen, Marjolein Couvreur, Sergei A. Subbotin, and Wim Bert. 2021. "Integrative Taxonomy and Molecular Phylogeny of the Plant-Parasitic Nematode Genus Paratylenchus (Nematoda: Paratylenchinae): Linking Species with Molecular Barcodes" Plants 10, no. 2: 408. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020408