
Recent Efforts to Recover Armeria berlengensis, an Endemic Species from Berlengas Archipelago, Portugal

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
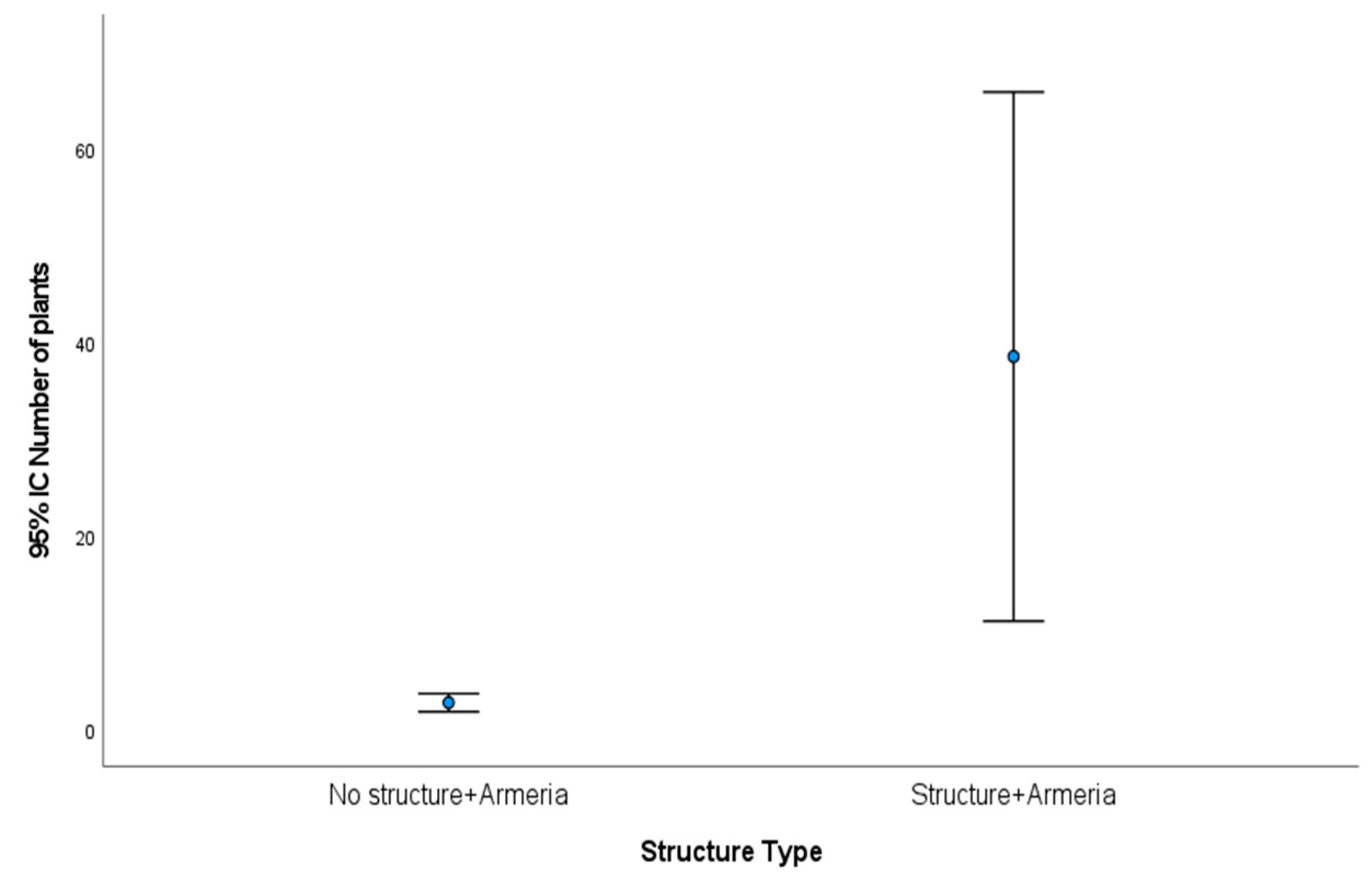
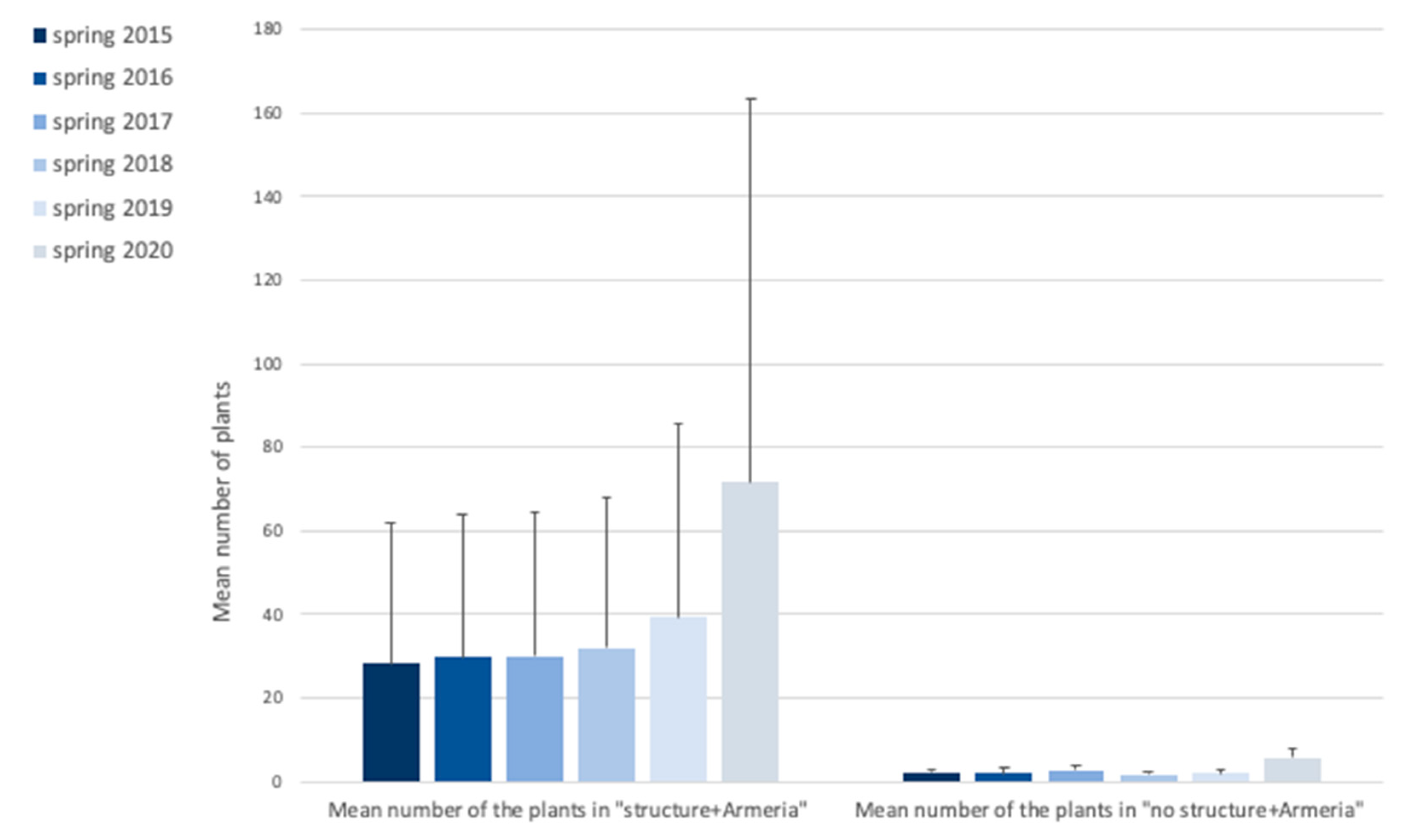
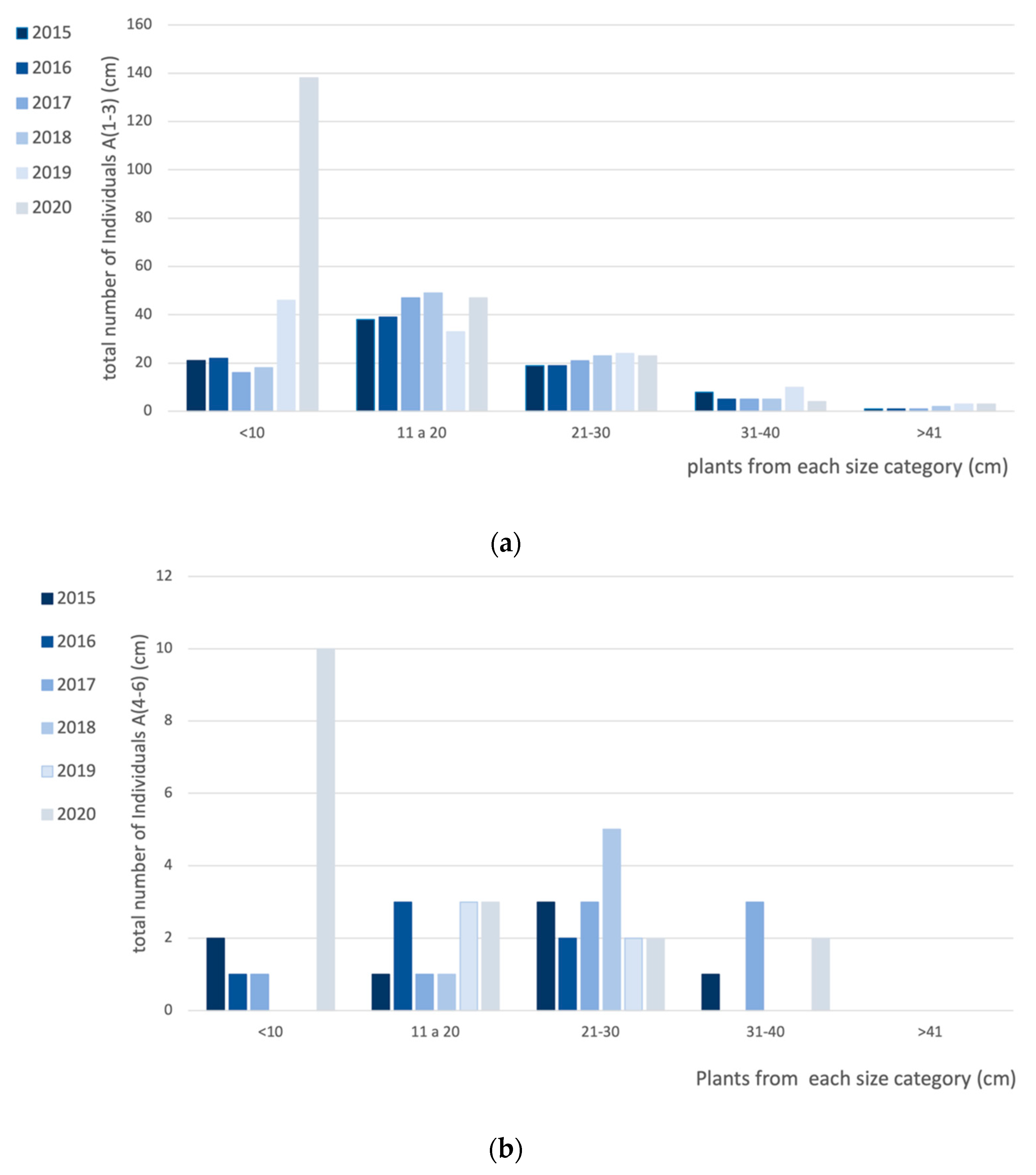
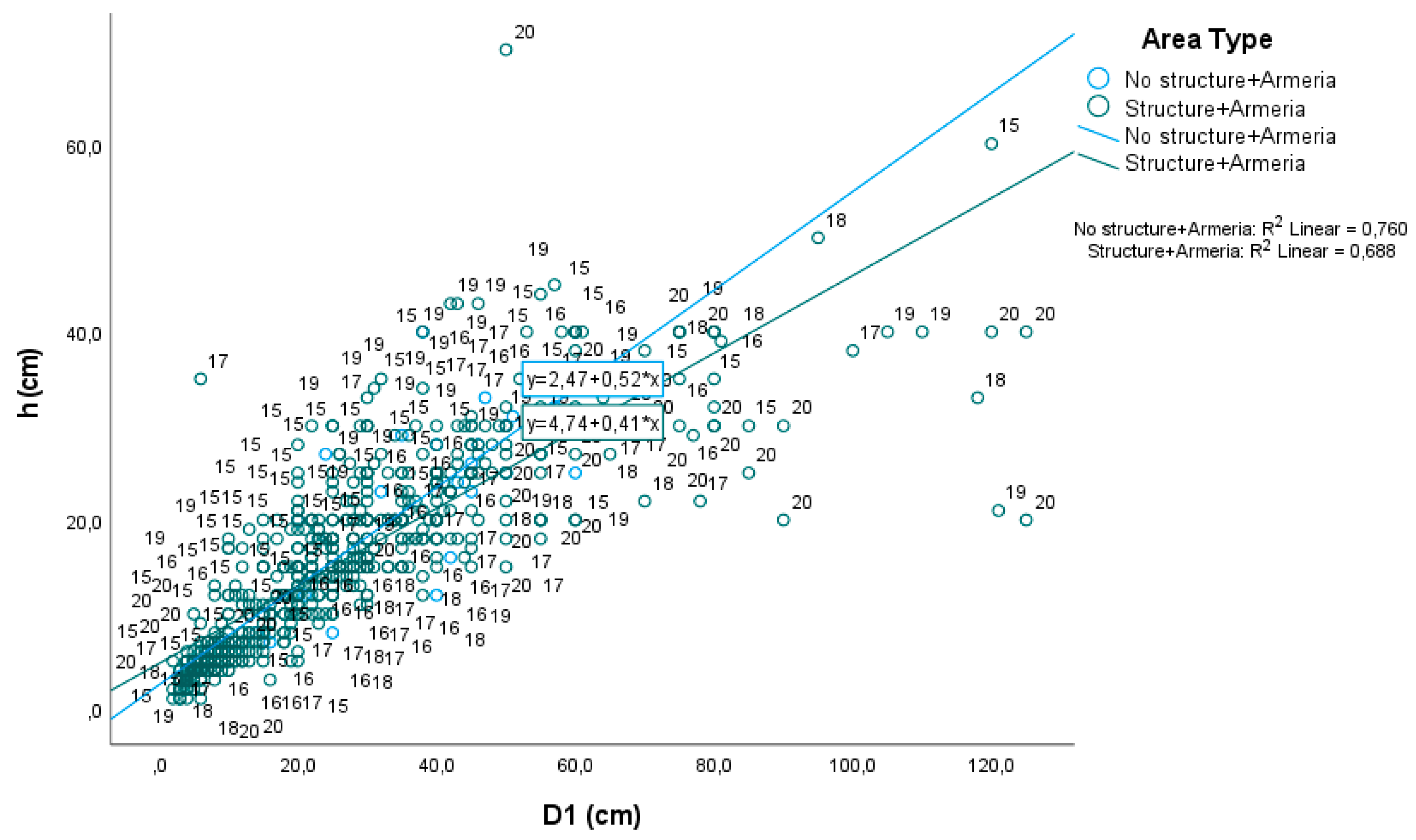
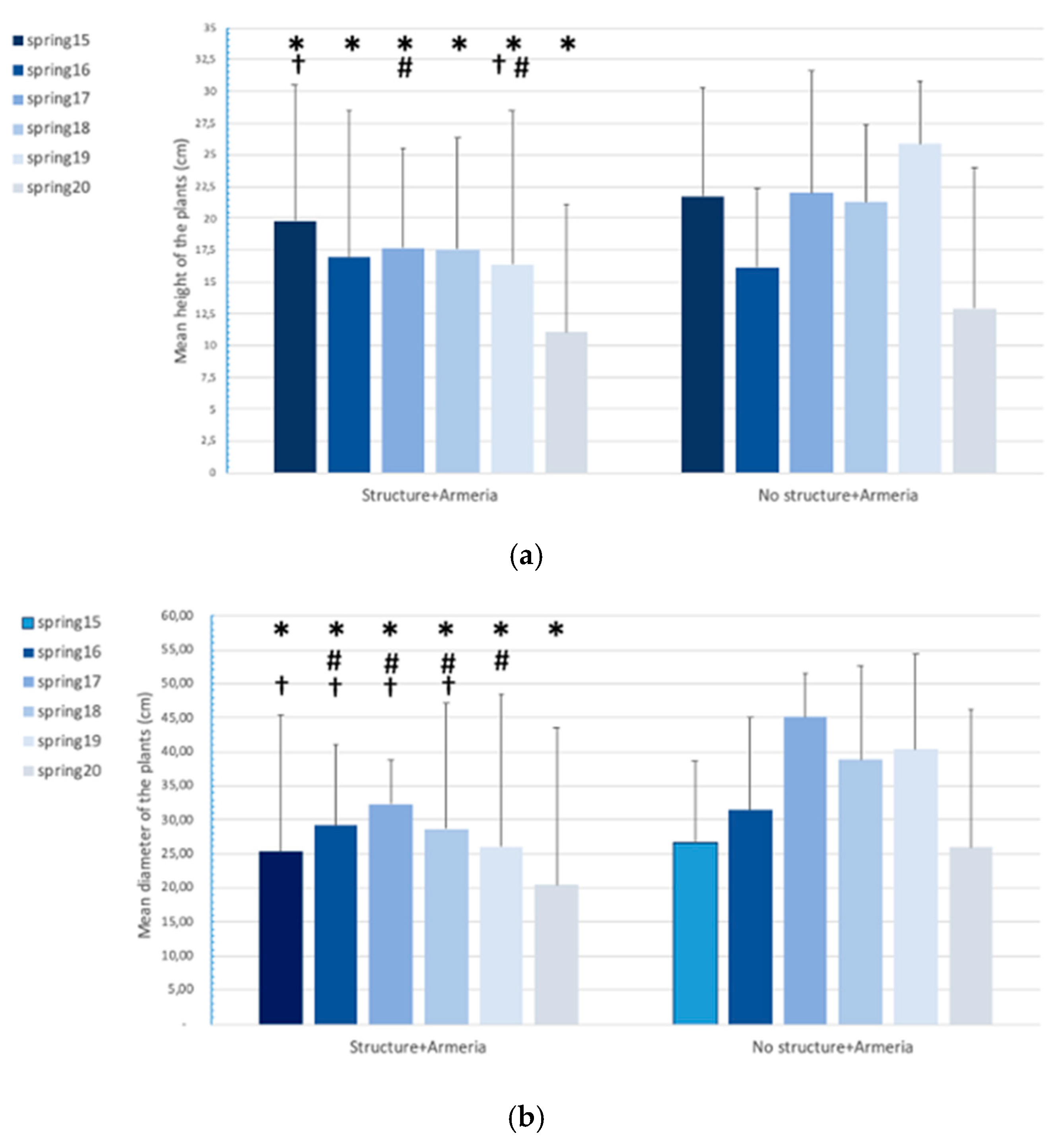
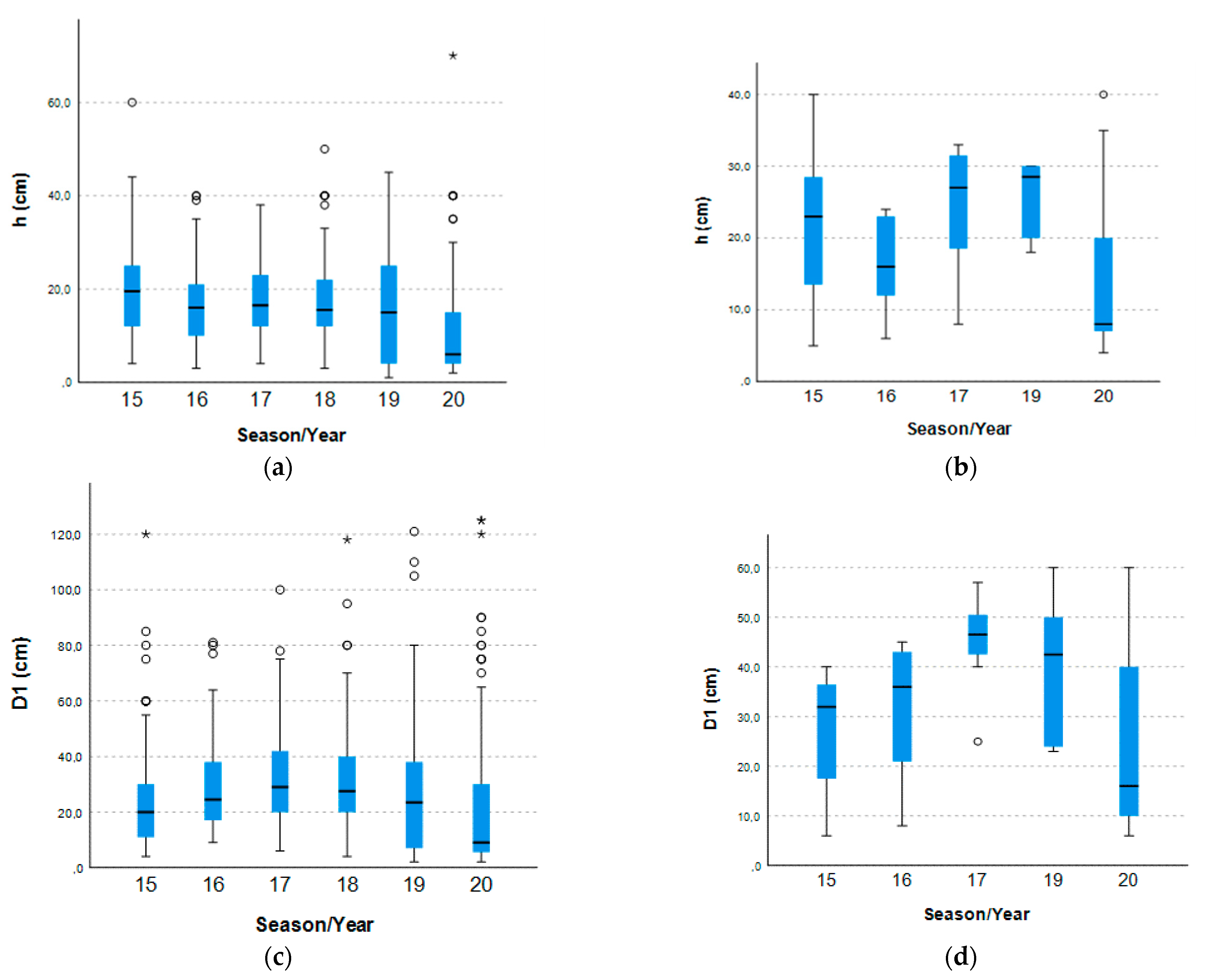
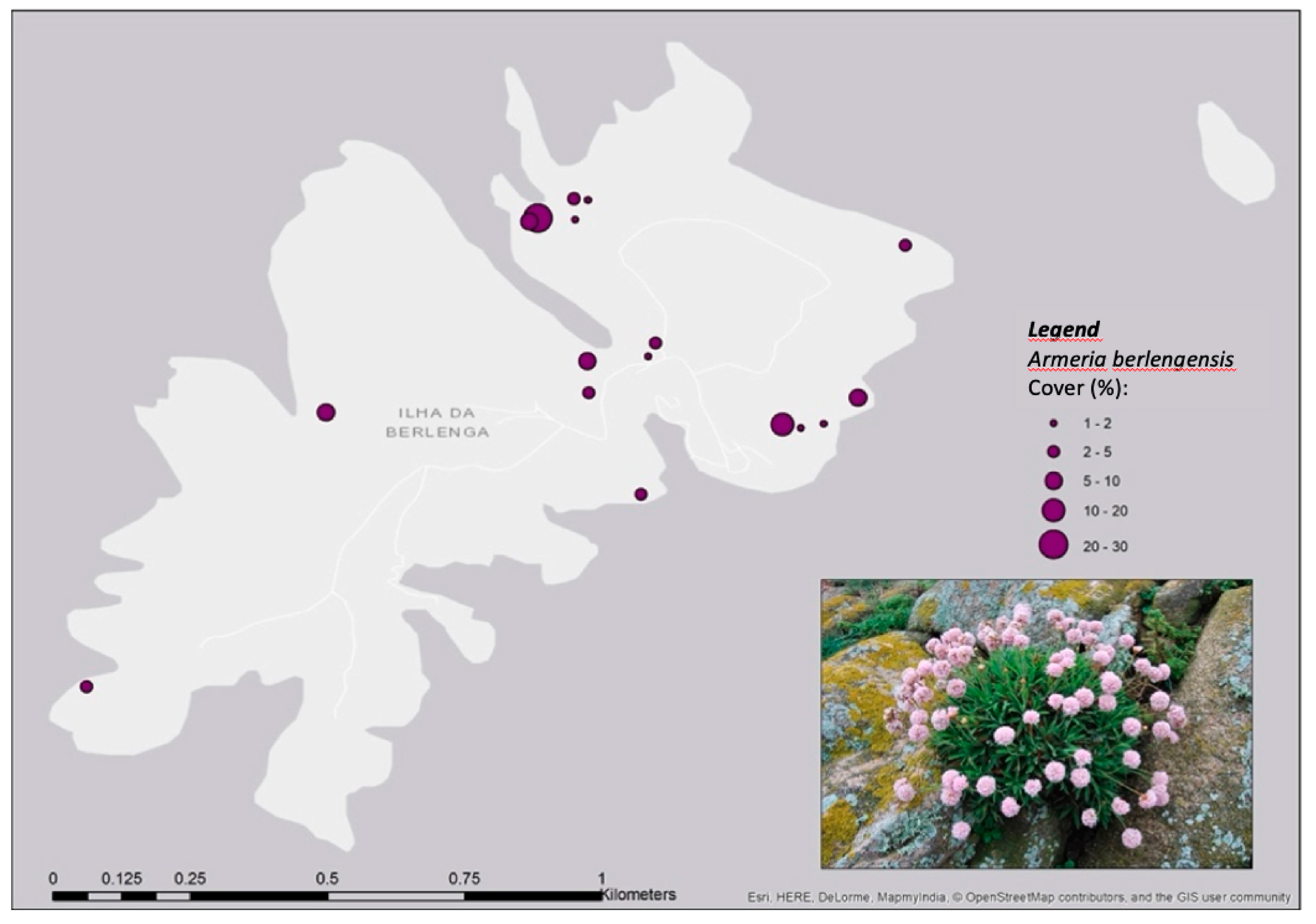
2.1. Armeria Berlengensis Surveys
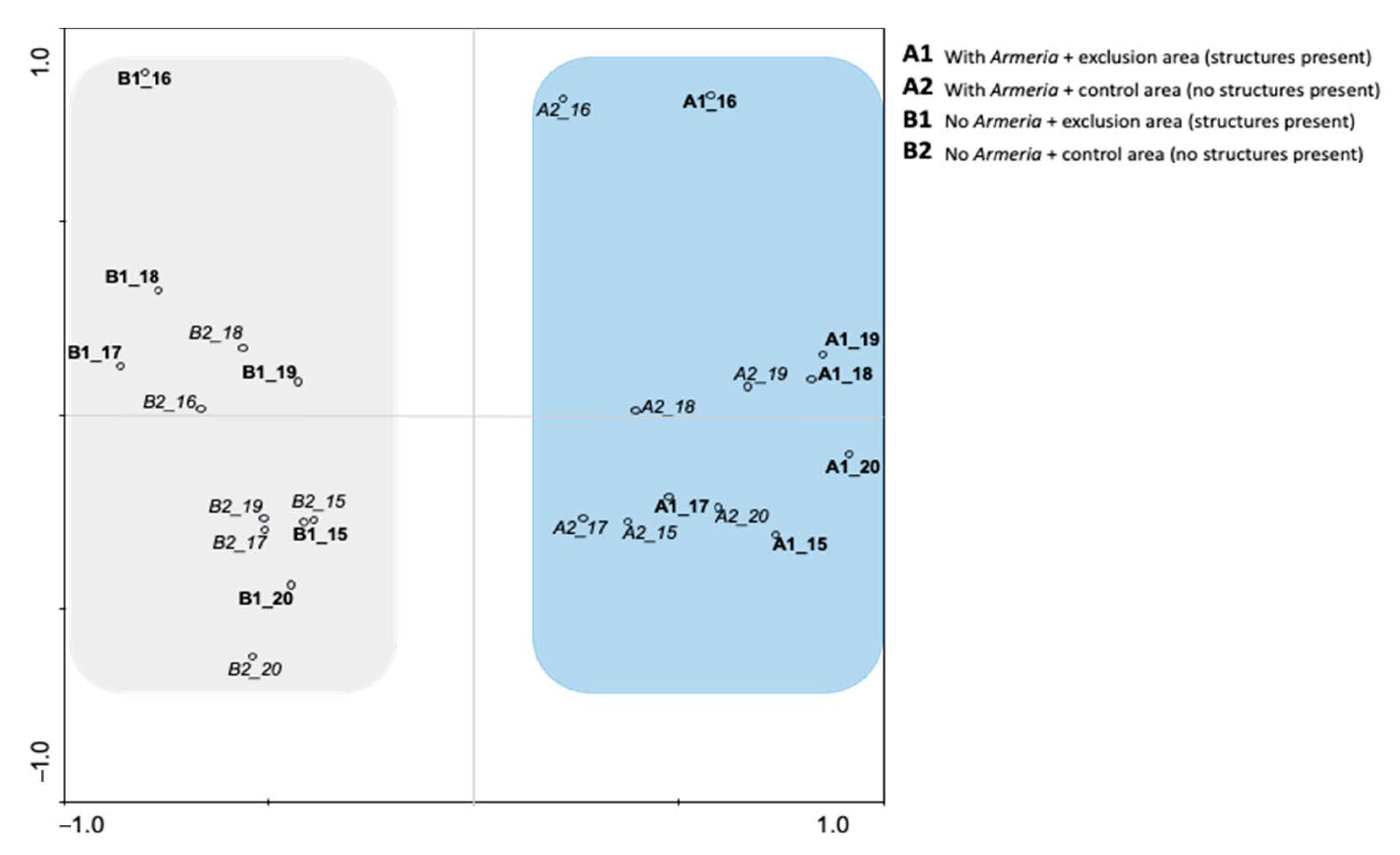
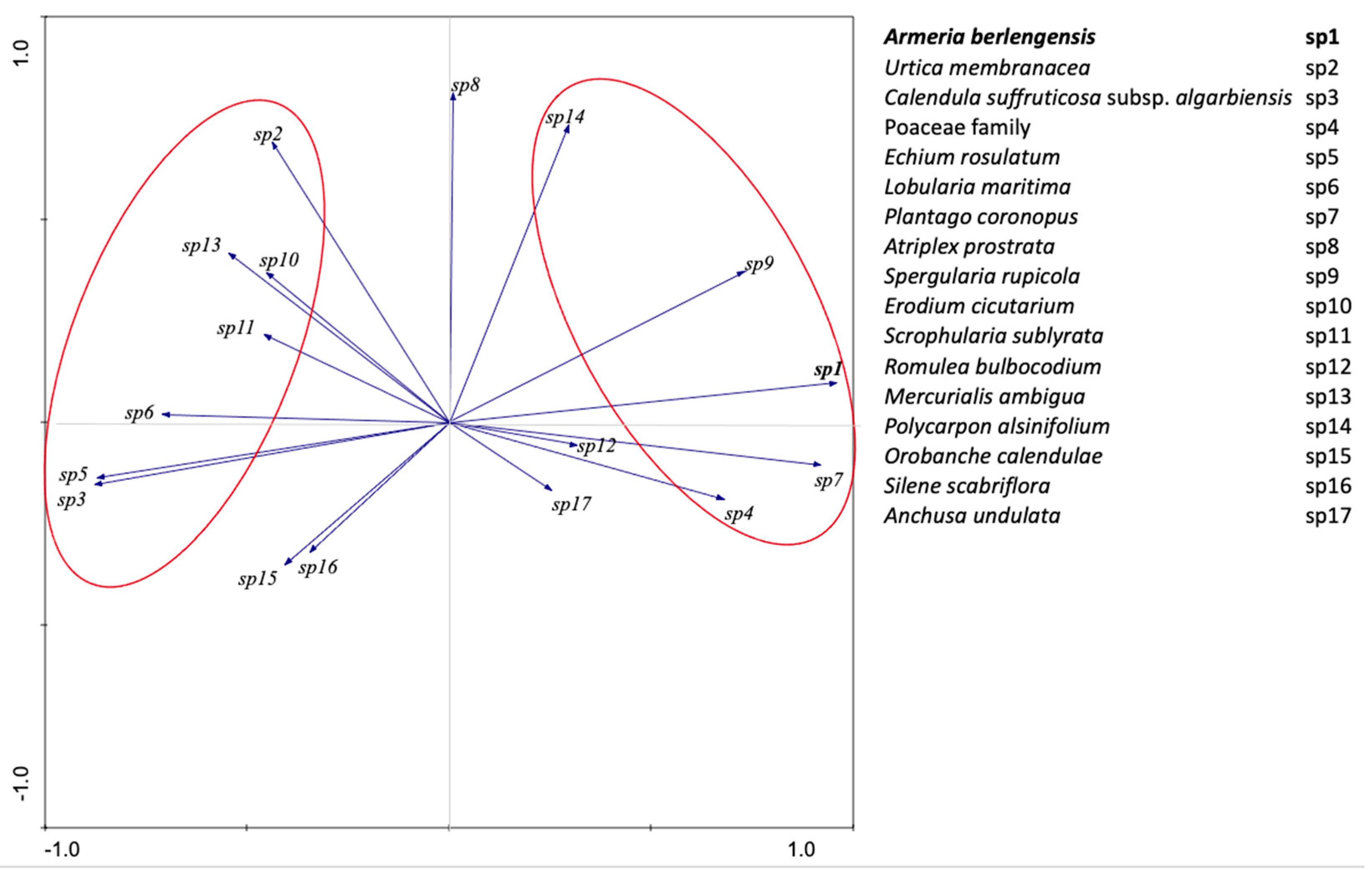
2.2. PCA Analysis
2.3. Armeria berlengensis Seed Set
3. Discussion
4. Materials and Methods
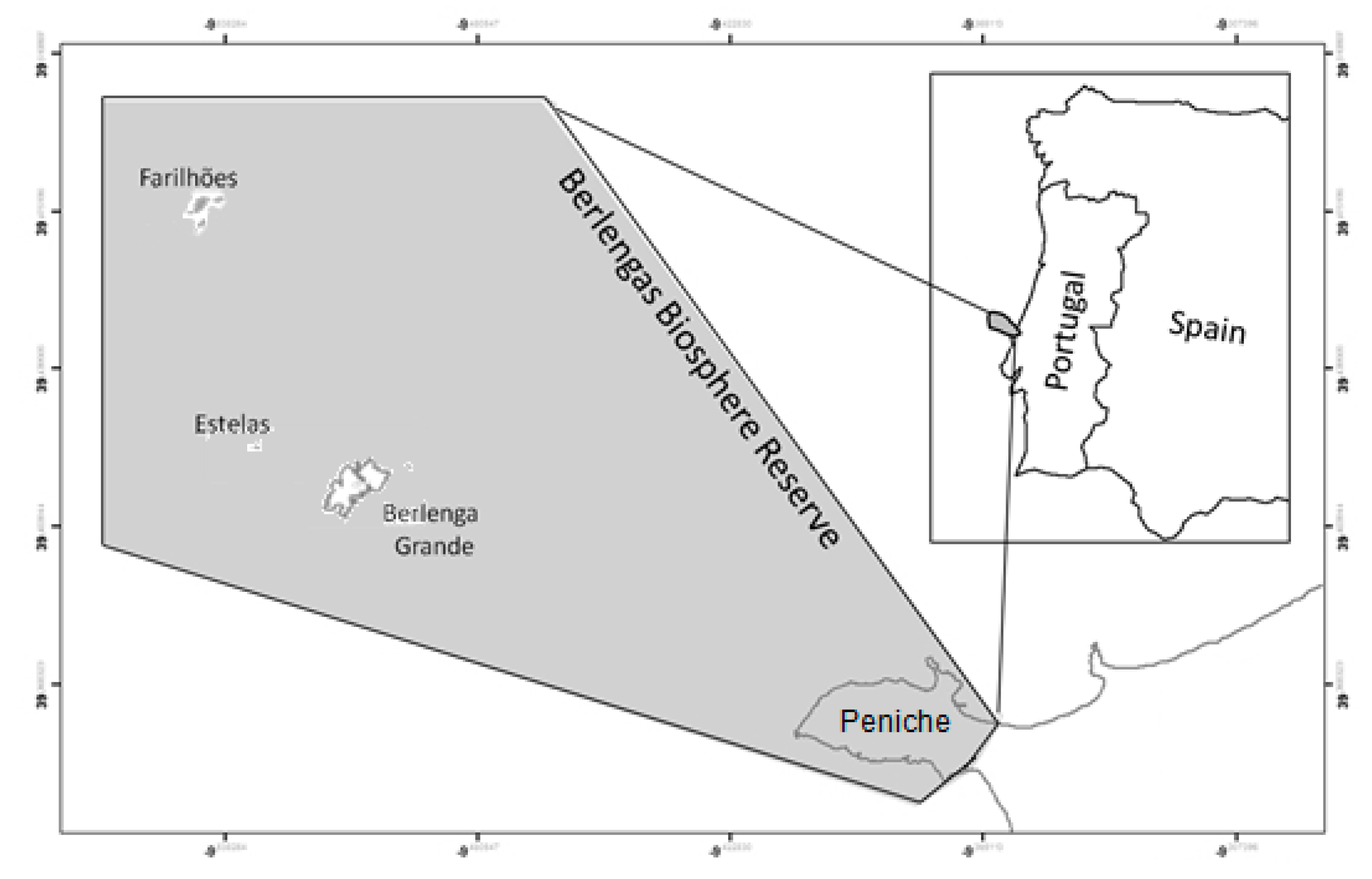
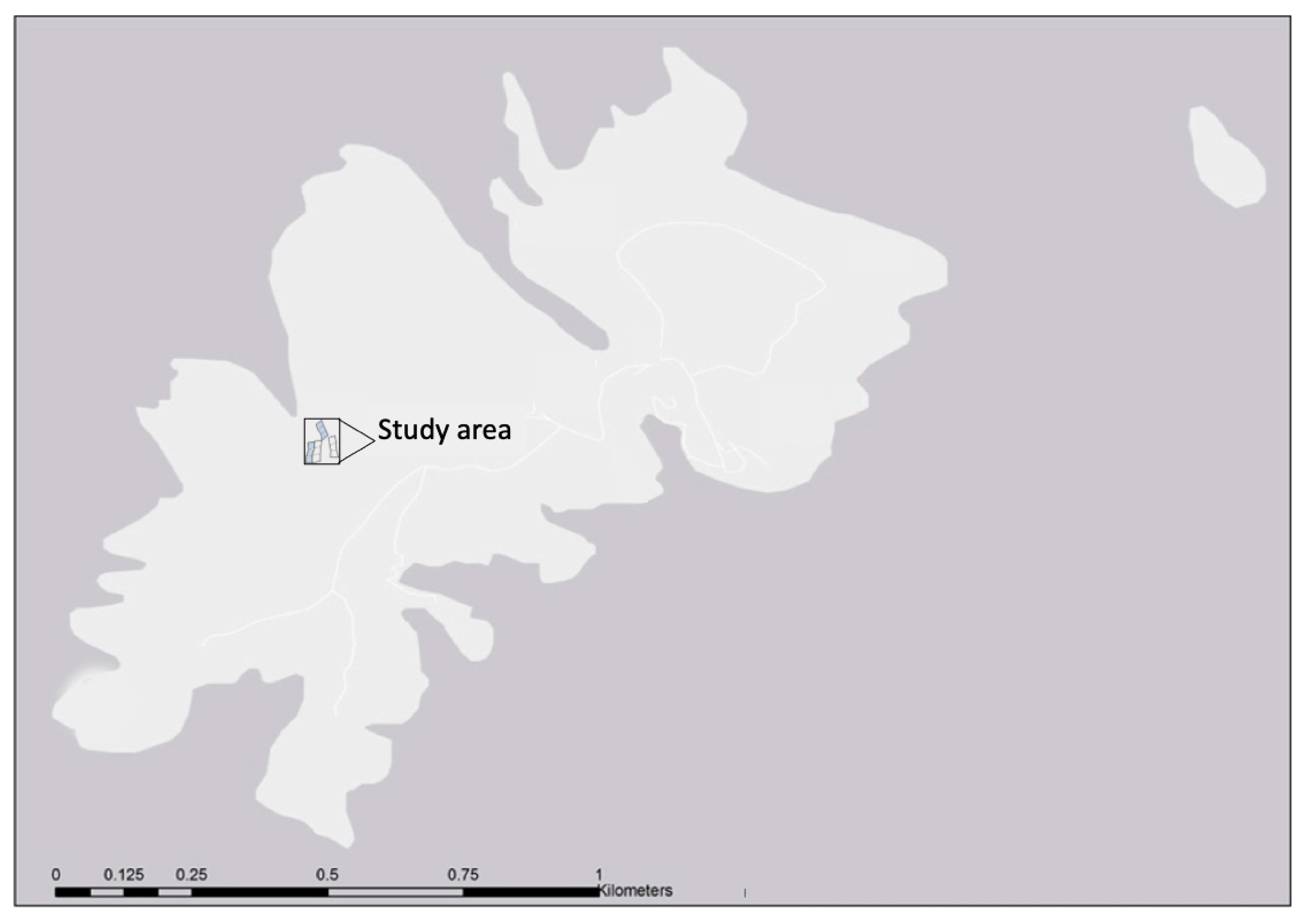
4.1. Study Area
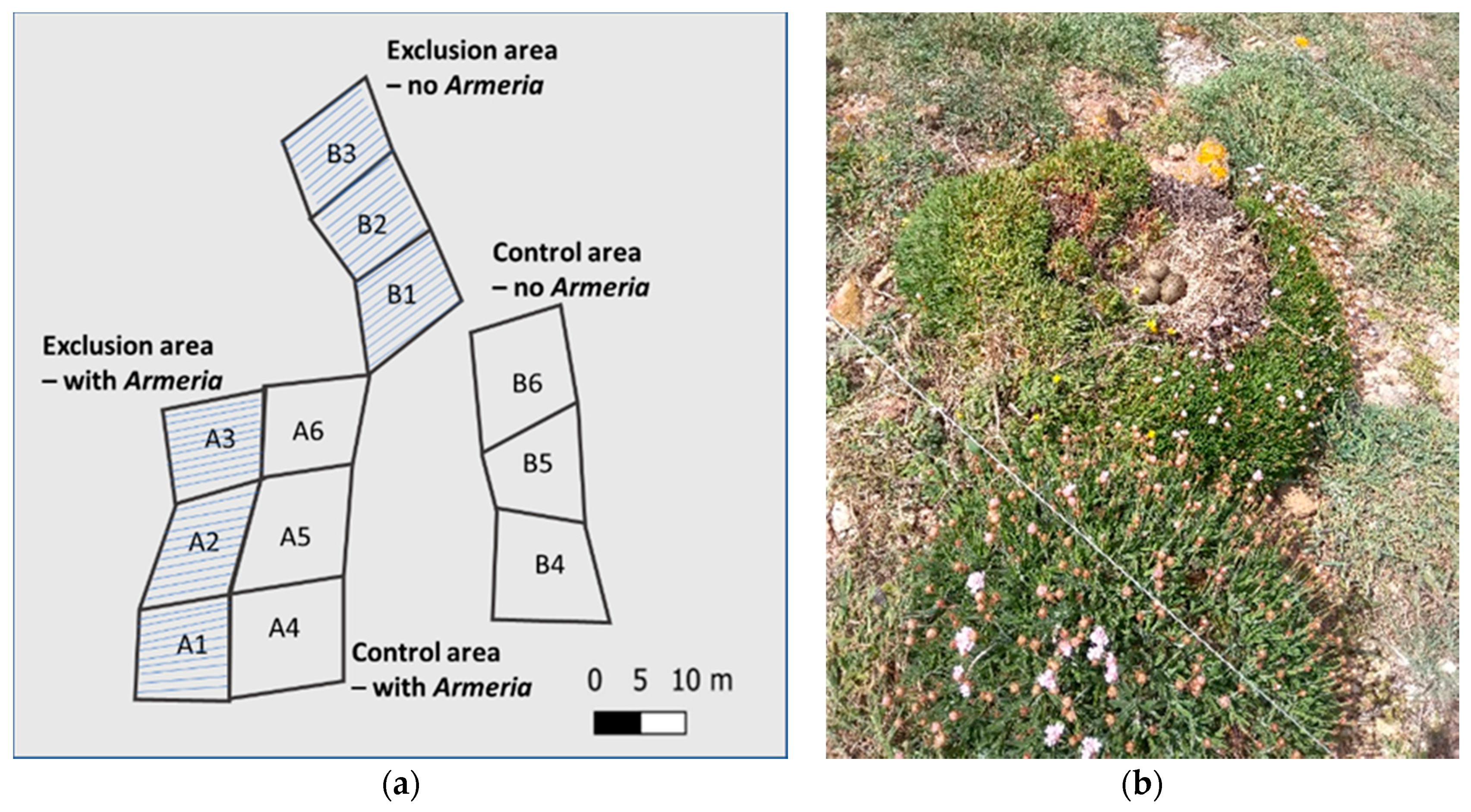
4.2. Armeria berlengensis Morphology and Distribution
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amado, A.; Gafeira, C.; Teixeira, A.; Preto, A.; Bártolo, P.; Fonseca, H.; Moris, L. Plano de Ordenamento da Reserva Natural Das Berlengas; Instituto da Conservação da Natureza e da Biodiversidade: Lisboa, Portugal, 2007. [Google Scholar]
- Queiroga, H.; Leão, F.; Coutinho, M. Candidatura das Berlengas a Reserva da Biosfera da UNESCO; Instituto do Ambiente e Desenvolvimento: Aveiro, Portugal, 2008; p. 128. [Google Scholar]
- Crawford, D.J.; Archibald, J.K. Island floras as model systems for studies of plant speciation: Prospects and challenges. J. Syst. Evol. 2017, 55, 1–15. [Google Scholar] [CrossRef] [Green Version]
- The Top 50 Mediterranean Island Plants: Wild Plants at the Brink of Extinction, and What is Needed to Save Them; Montmollin, B.; Strahm, W. (Eds.) UCN/SSC Me.: Gland, Switzerland; Cambridge, UK, 2005; ISBN 283170832X. [Google Scholar]
- Thompson, J.D. Population differentiation in Mediterranean plants: Insights into colonization history and the evolution and conservation of endemic species. Heredity 1999, 82, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Givnish, T.J. Ecology of plant speciation. Taxon 2010, 59, 1326–1366. [Google Scholar] [CrossRef]
- Azevêdo, T.; Nunes, E. The evolution of the coastline at Peniche and the Berlengas Islands (Portugal)—State of the art. Egypt. J. Environ. Chang. 2010, 2, 13–23. [Google Scholar]
- Do Amaral Franco, J.; da Rocha Afonso, M.D.L. Nova Flora de Portugal:(Continente e Açores); Escolar: Lisboa, Portugal, 1971; Volume 10. [Google Scholar]
- Castroviejo, S.; Laínz, G.; López-González, G.; Montserrat, P.; Muñoz-Garmendia, F.; Paiva, J.; Villar, L. Plumbaginaceae, Flora iberica 2; Real Jardim Botánico, C.S.I.C.: Madrid, Spain, 1990. [Google Scholar]
- Almada, F. Contribuição para a determinação do estatuto taxonómico de Pulicaria microcephala Lange (Asteraceae). Sep. Bol. Soc. Broteriana 1995, 67, 43–54. [Google Scholar]
- Baumel, A.; Youssef, S.; Ongamo, G.; Medail, F. Habitat suitability assessment of the rare perennial plant Armeria arenaria (Pers.) Schult. (Plumbaginaceae) along the French Mediterranean coastline. Candollea 2013, 68, 221–228. [Google Scholar] [CrossRef]
- Trias-Blasi, A.; Eddie, W.M.M.; Hedge, I.C.; Möller, M.; Sales, F. The taxonomy and conservation of Campanula primulifolia (Campanulaceae), a critically endangered species in the Iberian Peninsula. Willdenowia 2011, 41, 35–42. [Google Scholar] [CrossRef]
- Foggi, B.; Viciani, D.; Baldino, R.M.; Carta, A.; Guidi, T. Conservation assessment of the endemic plants of the Tuscan Archipelago, Italy. Oryx 2014, 49, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Wagensommer, R.P.; Medagli, P.; Turco, A.; Perrino, E.V. IUCN red list evaluation of the Orchidaceae endemic to Apulia (Italy) and considerations on the application of the IUCN protocol to rare species. Nat. Conserv. Res. 2020, 5 (Suppl 1.), 90–101. [Google Scholar] [CrossRef]
- Draper, D.; Rosselló-Graell, A.; Garcia, C.; Gomes, C.T.; Sérgio, C. Application of GIS in plant conservation programmes in Portugal. Biol. Conserv. 2003, 113, 337–349. [Google Scholar] [CrossRef]
- SPEA (Sociedade Portuguesa para o Estudo das Aves). LIFE Berlengas—Gestão Sustentável para a Conservação das Espécies e Habitats Ameaçados na ZPE das Berlengas: Relatório Final; SPEA: Lisboa, Portugal, 2019. [Google Scholar]
- Tauleigne Gomes, C. Nouvelles citations pour la flore de l’archipel des Berlengas (Estremadura, Portugal). Acta Bot. Malacit. 2002, 27, 261–268. [Google Scholar] [CrossRef]
- García, L.V.; Marañón, T.; Ojeda, F.; Clemente, L.; García, R.R.; Ojeda, T.; Redondo, C.L. Seagull influence on soil properties, chenopod shrub distribution, and leaf nutrient status in semi-arid Mediterranean islands. Oikos 2002, 98, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Tauleigne Gomes, C.; Draper, D.; Rosse-Graell, A. Données sur la végétation halophile; l’archipel des Berlengas (Estremadura, Portugal). Bocconea 2001, 13, 537–542. [Google Scholar]
- Otero, X.L.; Tejada, O.; Martín-Pastor, M.; De La Peña, S.; Ferreira, T.O.; Pérez-Alberti, A. Phosphorus in seagull colonies and the effect on the habitats. The case of yellow-legged gulls (Larus michahellis) in the Atlantic Islands National Park (Galicia-NW Spain). Sci. Total Environ. 2015, 532, 383–397. [Google Scholar] [CrossRef]
- Vidal, E.; Medail, F.T.; Tatoni, T. Is the yellow-legged gull a superabundant bird species in the Mediterranean? Impact on fauna and flora, conservation measures and research priorities. Biodivers. Conserv. 1998, 7, 1013–1026. [Google Scholar] [CrossRef]
- Vidal, E.; Médail, F.T.; Atoni, T.; Bonnet, V. Seabirds drive plant species turnover on small Mediterranean islands at the expense of native taxa. Oecologia 2000, 122, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Groves, F.R.; Di Castro, F. Biogeography of Mediterranean Invasions; Cambridge University Press: Cambridge, UK, 1991; ISBN 0521360404. [Google Scholar]
- Nascimento, T. Caracterização de Uma População Insular de Roedores Invasores. O Caso Do Rato Preto (Rattus Rattus) NA Ilha Da Berlenga; University of Aveiro: Aveiro, Portugal, 2016. [Google Scholar]
- Novoa, A.; González, L.; Moravcová, L.; Pyšek, P. Constraints to native plant species establishment in coastal dune communities invaded by Carpobrotus edulis: Implications for restoration. Biol. Conserv. 2013, 164, 1–9. [Google Scholar] [CrossRef]
- Pysek, P.; Genovesi, P.; Pergl, J.; Monaco, A.; Wild, J. Plant Invasions in Protected Areas; Springer: Dordrecht, The Netherland, 2013; Volume 7, ISBN 978-94-007-7749-1. [Google Scholar]
- Van Der Maarel, E.; Van Der Maarel-Versluys, M. Distribution and conservation status of littoral vascular plant species along the European coasts. J. Coast. Conserv. 1996, 2, 73–92. [Google Scholar] [CrossRef]
- Caldas, F.B. Armeria Berlengensis. The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/details/162004/0 (accessed on 9 April 2018).
- Tauleigne Gomes, C.; Draper, D.; Marques, I. Componente Vegetal do Plano de Ordenamento da Reserva Natural das Berlengas; Flora e Vegetação do Arquipélago as Berlengas: Lisboa, Portugal, 2004. [Google Scholar]
- Philipp, M.; Erik, H.; Madsen, S.; Redlef Siegismund, H. Gene flow and population structure in Armeria maritima. Heredity 1992, 69, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Albert, M.J.; Escudero, A.; Iriondo, J.M. Environmental range of narrow endemic Erodium paularense Fern. Gonz. & Izeo and its vulnerability to changing climatic conditions. Bocconea 2001, 13, 549–556. [Google Scholar]
- Catarino, F.M.; Correia, O.A.; Correia, A.I.V.D. Structure and dynamics of Serra da Arrábida mediterranean vegetation. Ecol. Mediterr. 1982, 8, 203–222. [Google Scholar] [CrossRef]
- Asensi, A.; Diez-Garretas, B. Coastal Vegetation. In The Vegetation of the Iberian Peninsula, Plant and Vegetation; Loidi, J., Ed.; Springer International Publishing: Cham, Switzerland, 2017; Volume 13, pp. 397–432. ISBN 978-3-319-54866-1. [Google Scholar]
- Vitousek, P.M. Diversity and biological invasions of oceanic islands. In Biodiversity; Wilson, E.O., Peter, F.M., Eds.; National Academy Press (US): Washington, DC, USA, 1988; ISBN 0-309-03783-2. [Google Scholar]
- Le Corre, M.; Danckwerts, D.K.; Ringler, D.; Bastien, M.; Orlowski, S.; Morey Rubio, C.; Pinaud, D.; Micol, T. Seabird recovery and vegetation dynamics after Norway rat eradication at Tromelin Island, western Indian Ocean. Biol. Conserv. 2015, 185, 85–94. [Google Scholar] [CrossRef]
- Scott, J.J.; Kirkpatrick, J.B. Rabbits, landslips and vegetation change on the coastal slopes of subantarctic Macquarie Island, 1980-2007: Implications for management. Polar Biol. 2008, 31, 409–419. [Google Scholar] [CrossRef]
- Canale, D.E.; Dio, V.D.; Massa, B.; Mori, E. First successful eradication of invasive Norway rats Rattus norvegicus from a small Mediterranean island (Isola delle Femmine, Italy). Folia Zool. 2019, 68, 29. [Google Scholar] [CrossRef]
- Eijzenga, H. Vegetation change following rabbit eradication on Lehua Island, Hawaiian Islands. In Proceedings of the International Conference on Island Invasives, Auckland, New Zealand, 8–12 February 2010; Veitch, C.R., Clout, M.N., Towns, D.R., Eds.; IUCN: Gland, Switzerland; pp. 290–294.
- Bell, B.D. The eradication of alien mammals from five offshore islands, Mauritius, Indian Ocean. In Turning the Tide: The Eradication of Invasive Species; Veitch, C.R., Clout., M.N., Eds.; IUCN: Gland, Switzerland; Cambridge, UK, 2002; pp. 40–45. [Google Scholar]
- Verdú, J.R.; Crespo, M.B.; Galante, E. Conservation strategy of a nature reserve in Mediterranean ecosystems: The effects of protection from grazing on biodiversity. Biodivers. Conserv. 2000, 9, 1707–1721. [Google Scholar] [CrossRef]
- Rita, J.; Biblioni, G. The flora of the islets of the Balearic Islands. In Islands and Plants: Preservation and Understanding of Flora on Mediterranean Islands: 2nd Botanical Conference in Menorca: Proceedings and abstracts; Pons, E.C., Clarisó, I.E., Casademont, M.C., Arguimbau, P.F.I., Eds.; Institut Menorquí d’Estudis: Menorca, Spain, 2013; pp. 309–322. ISBN 978-8495718952. [Google Scholar]
- Espírito-Santo, M.D.; Capelo, J.; Neto, C.; Pinto-Gomes, C.; Ribeiro, S.; Quinto Canas, R.; Costa, J.C. Lusitania. In The Vegetation of the Iberian Peninsula, Plant and Vegetation 13, Loidi, J., Ed.; Springer: Cham, Switzerland, 2017; pp. 35–82. [Google Scholar]
- Costa, J.C.; Lousã, M.; Capelo, J.; Espírito-Santo, M.D.; Sevillano, J.I.; Arsénio, P. The coastal vegetation of the portuguese divisory sector: Dunes cliffs and low-scrub communities. Finisterra 2000, 69, 69–93. [Google Scholar] [CrossRef] [Green Version]
- Tomassen, H.B.M.; Smolders, A.J.P.; Lamers, L.P.M.; Roelofs, J.G.M. How bird droppings can affect the vegetation composition of ombrotrophic bogs. Can. J. Bot. 2005, 83, 1046–1056. [Google Scholar] [CrossRef]
- Sigurdsson, B.D. Effects of sea birds and soil development on plant and soil nutritional parameters after 50 years of succession on Surtsey. Surtsey Res. 2020, 14, 85–90. [Google Scholar]
- De La Peña-Lastra, S.; Gómez-Rodríguez, C.; Pérez-Alberti, A.; Torre, F.; Otero, X.L. Effects of a yellow legged gull (Larus michahellis) colony on soils and cliff vegetation in the Atlantic Islands of Galicia National Park (NW Spain). Catena 2021, 199, 105115. [Google Scholar] [CrossRef]
- Tauleigne-Gomes, C. Armeria X cintrana nothoespecie nova para o género Armeria Willd. (Plumbaginaceae). Acta Bot. Malacit. 2011, 36, 183–185. [Google Scholar] [CrossRef]
- Alonso, H.; Almeida, A.; Granadeiro, J.P.; Catry, P. Temporal and age-related dietary variations in a large population of yellow-legged gulls Larus michahellis: Implications for management and conservation. Eur. J. Wildl. Res. 2015, 61, 819–829. [Google Scholar] [CrossRef]
- Tauleigne-Gomes, C.; Lefèvre, C. Natural hybridisation between two coastal endemic species of Armeria (Plumbaginaceae) from Portugal. 1. Populational in situ investigations. Plant Syst. Evol. 2005, 250, 215–230. [Google Scholar] [CrossRef]
- Baker, K.; Richards, A.J.; Tremayne, M. Fitness constraints on flower number, seed number and seed size in the dimorphic species Primula farinosa L. and Armeria maritima (Miller) Willd. New Phytol. 1994, 128, 563–570. [Google Scholar] [CrossRef]
- Daveau, J. Excursion botanique aux îles Berlengas et Farilhões par J. Daveau. Bol. Soc. Broteriana 1883, 2, 13–31. [Google Scholar]
- Coutinho, A.X.P. A Flora de Portugal; Aillaud, Alves: Paris, France, 1913. [Google Scholar]
- Do Amaral Franco, J.; Afonso, M.R. Nova Flora de Portugal: (Continente e Açores); Escolar Editora: Lisboa, Portugal, 1984; Volume 20. [Google Scholar]
- Fuertes Aguilar, J.; Nieto, G.; Real, F.; Bot, J. Additive polymorphisms and reticulation in an ITS phylogeny of thrifts (Armeria, Plumbaginaceae). Mol. Phylogenet. Evol. 2003, 28, 430–447. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Legendre, L.; Legendre, P. Écologie Numérique. 1. Le Traitement Multiple des Données Écologiques; 12, 2nd ed.; D’écologie, C., Ed.; Masson: Paris, France, 1984. [Google Scholar]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and Canodraw forWindows User’s Guide: Software for Canonical Community Ordination; Version 4; Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Fagundes, A.I.; Filipe, A.; Oliveira, N.; Andrade, J. Control and Eradication of Invasive Exotic Plants, Report for Task c5; Project LIFE+ Berlengas: Lisboa, Portugal, 2018. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Number of Individuals | Spring15 | Spring16 | Spring17 | Spring18 | Spring19 | Spring20 |
|---|---|---|---|---|---|---|
| A(1–3): Structure + Armeria | 85 | 88 | 90 | 96 | 118 | 201 |
| A(4–6): No structure + Armeria | 7 | 6 | 8 | 5 | 6 | 17 |
| B(4–6): No strucuture + no Armeria | 1 | 0 | 0 | 0 | 0 | 0 |
| Total | 93 | 94 | 98 | 101 | 124 | 218 |
| 1 | 2 | 3 | 4 | Total Variance | |
|---|---|---|---|---|---|
| Eigenvalues | 0.367 | 0.149 | 0.119 | 0.083 | 1 |
| Cumulative percentage variance of species data | 36.7 | 51.6 | 63.5 | 71.8 | -- |
| Data Analyzed | # | |
|---|---|---|
| Fruits | Total number of fruits opened | 4721 |
| Mean number of fruits per cluster | 48.17 | |
| Seeds | Total number of seeds collected | 164 |
| Mean number of seeds collected | 1.67 | |
| Seed set | 3.47% |
| # | % | |
|---|---|---|
| Number of fruit clusters analyzes | 19 | -- |
| Seed set | 23 | 3.39 |
| Empty calyxes | 207 | 30.53 |
| Non-fertilized ovaries | 343 | 50.59 |
| Rotten calyxes | 105 | 15.49 |
| Total number of calyxes | 678 | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouga, T.; Mendes, S.; Franco, I.; Fagundes, A.I.; Oliveira, N.; Crisóstomo, P.; Morais, L.; Afonso, C. Recent Efforts to Recover Armeria berlengensis, an Endemic Species from Berlengas Archipelago, Portugal. Plants 2021, 10, 498. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030498
Mouga T, Mendes S, Franco I, Fagundes AI, Oliveira N, Crisóstomo P, Morais L, Afonso C. Recent Efforts to Recover Armeria berlengensis, an Endemic Species from Berlengas Archipelago, Portugal. Plants. 2021; 10(3):498. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030498
Chicago/Turabian StyleMouga, Teresa, Susana Mendes, Inês Franco, Ana Isabel Fagundes, Nuno Oliveira, Paulo Crisóstomo, Lurdes Morais, and Clélia Afonso. 2021. "Recent Efforts to Recover Armeria berlengensis, an Endemic Species from Berlengas Archipelago, Portugal" Plants 10, no. 3: 498. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030498