
Salinity Effects on Morpho-Physiological and Yield Traits of Soybean (Glycine max L.) as Mediated by Foliar Spray with Brassinolide

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Trial Management
2.2. Observations
2.2.1. Growth Variables
2.2.2. Nutrient Analysis
2.2.3. Plant Physiological Measurements
2.2.4. Biological Yield and Yield Components
2.2.5. Enzyme Extractions and Assays
2.3. Statistics
3. Results
3.1. Soil Analysis
3.2. Plant Growth
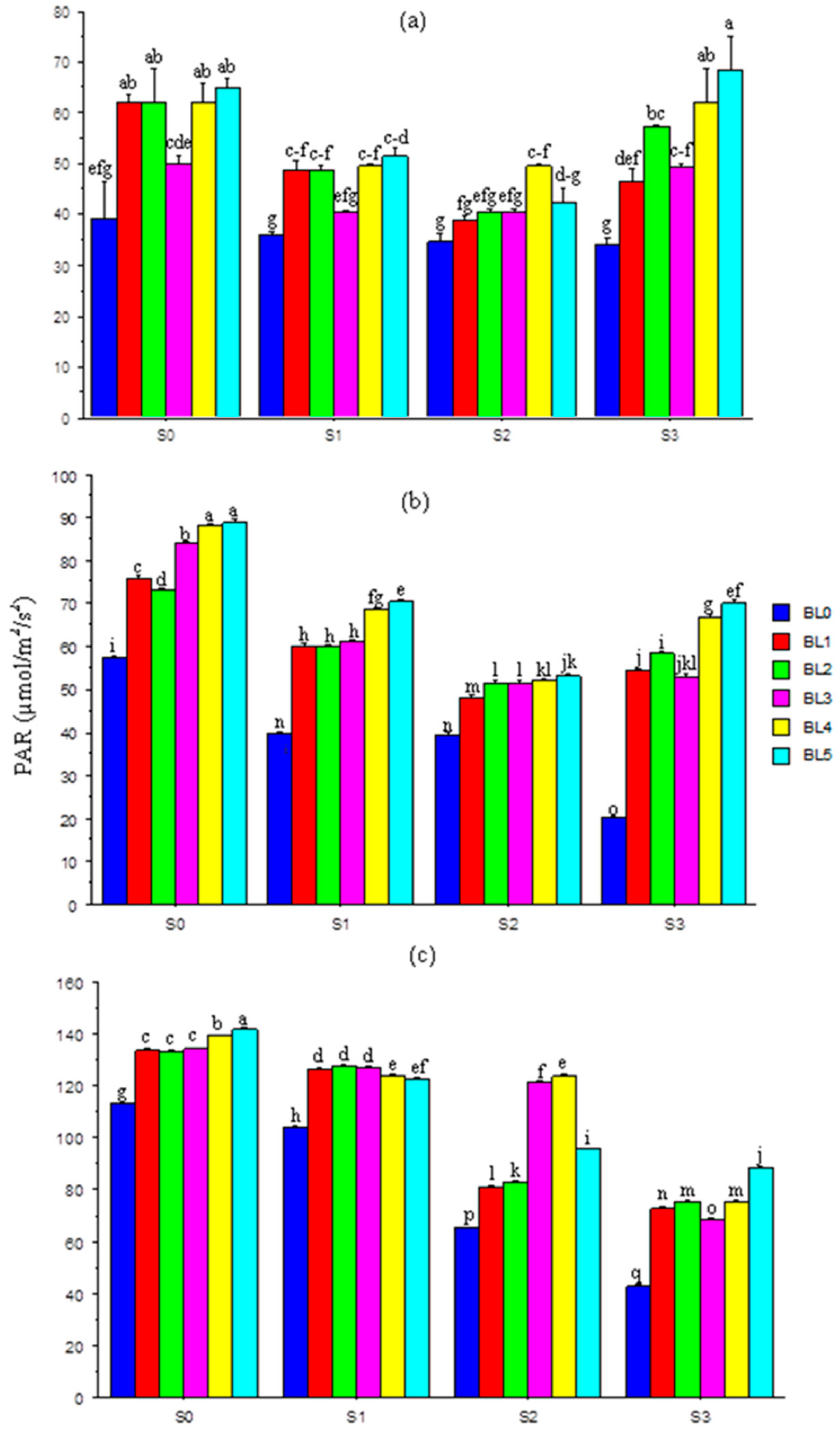
3.3. Photosynthetically Active Radiation
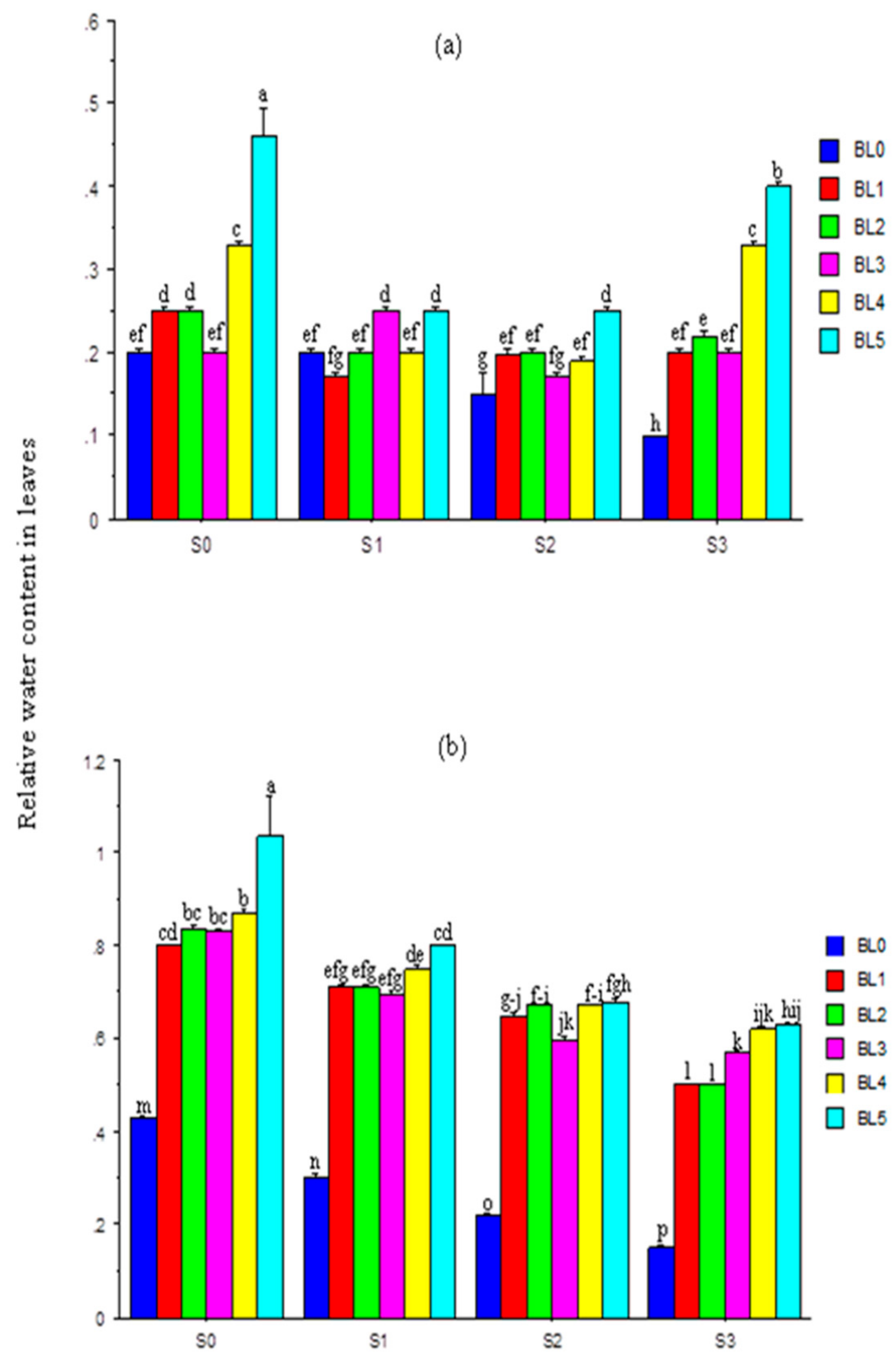
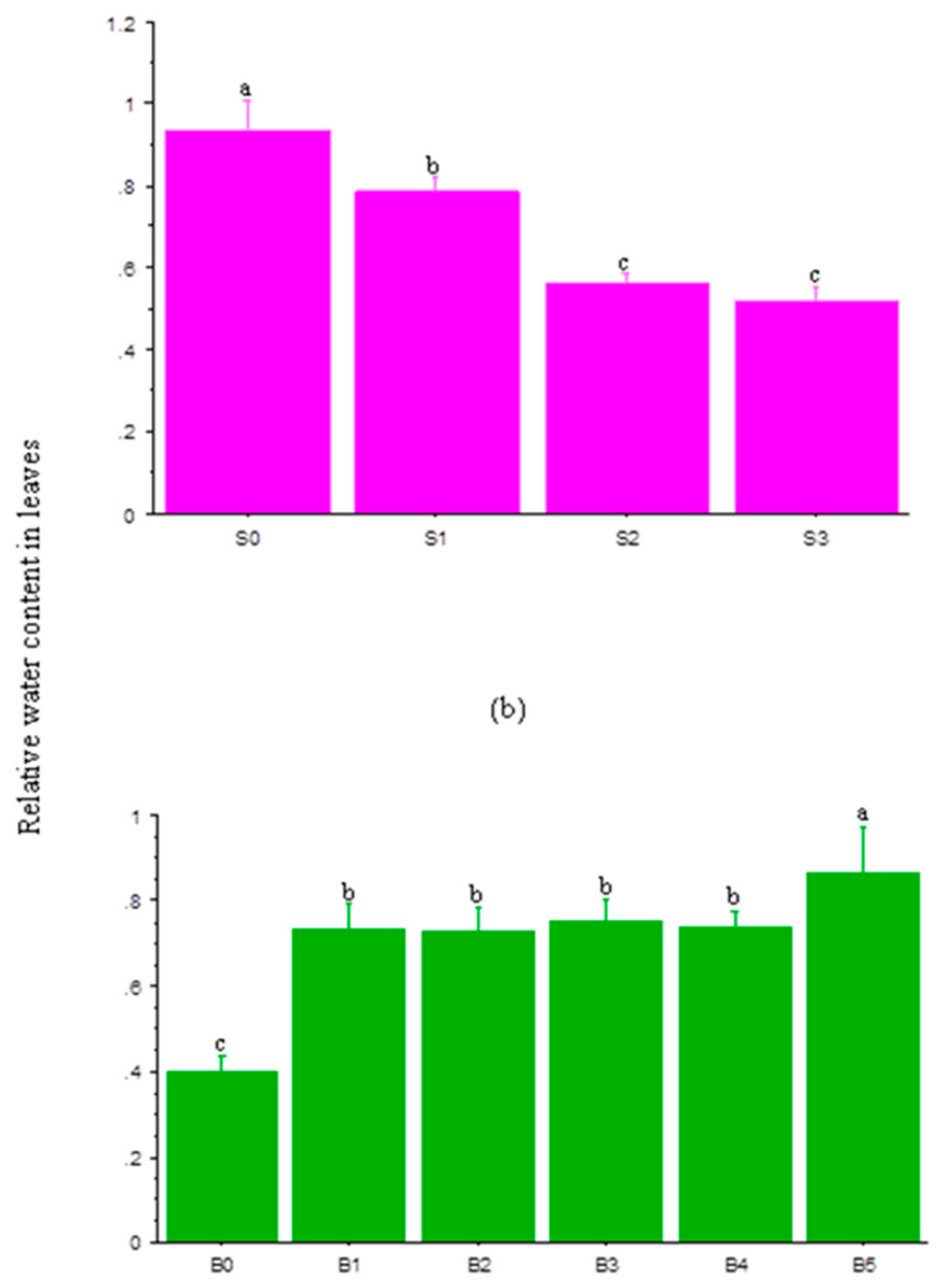
3.4. Relative Water Content in Leaves
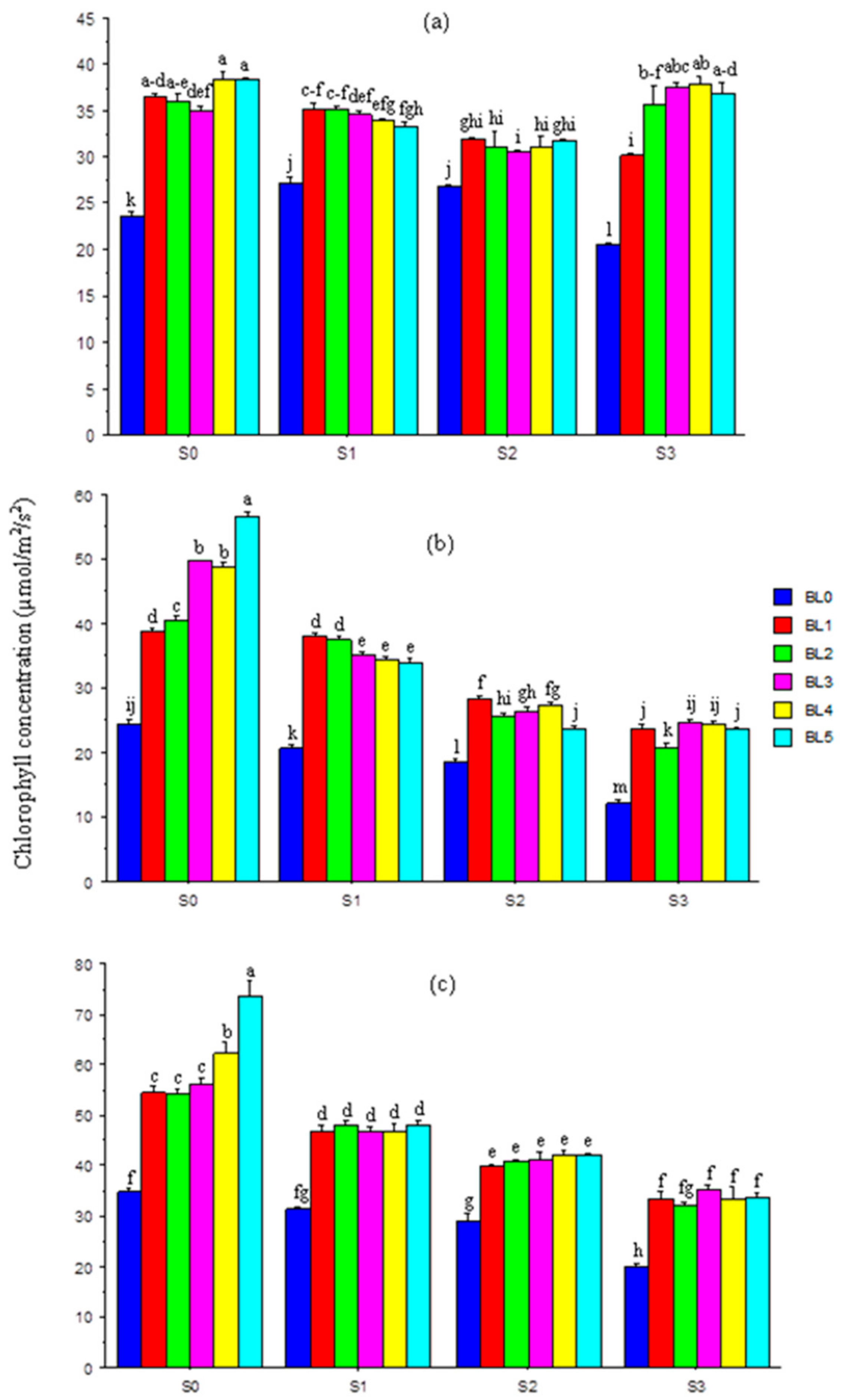
3.5. Chlorophyll Concentration in Leaves
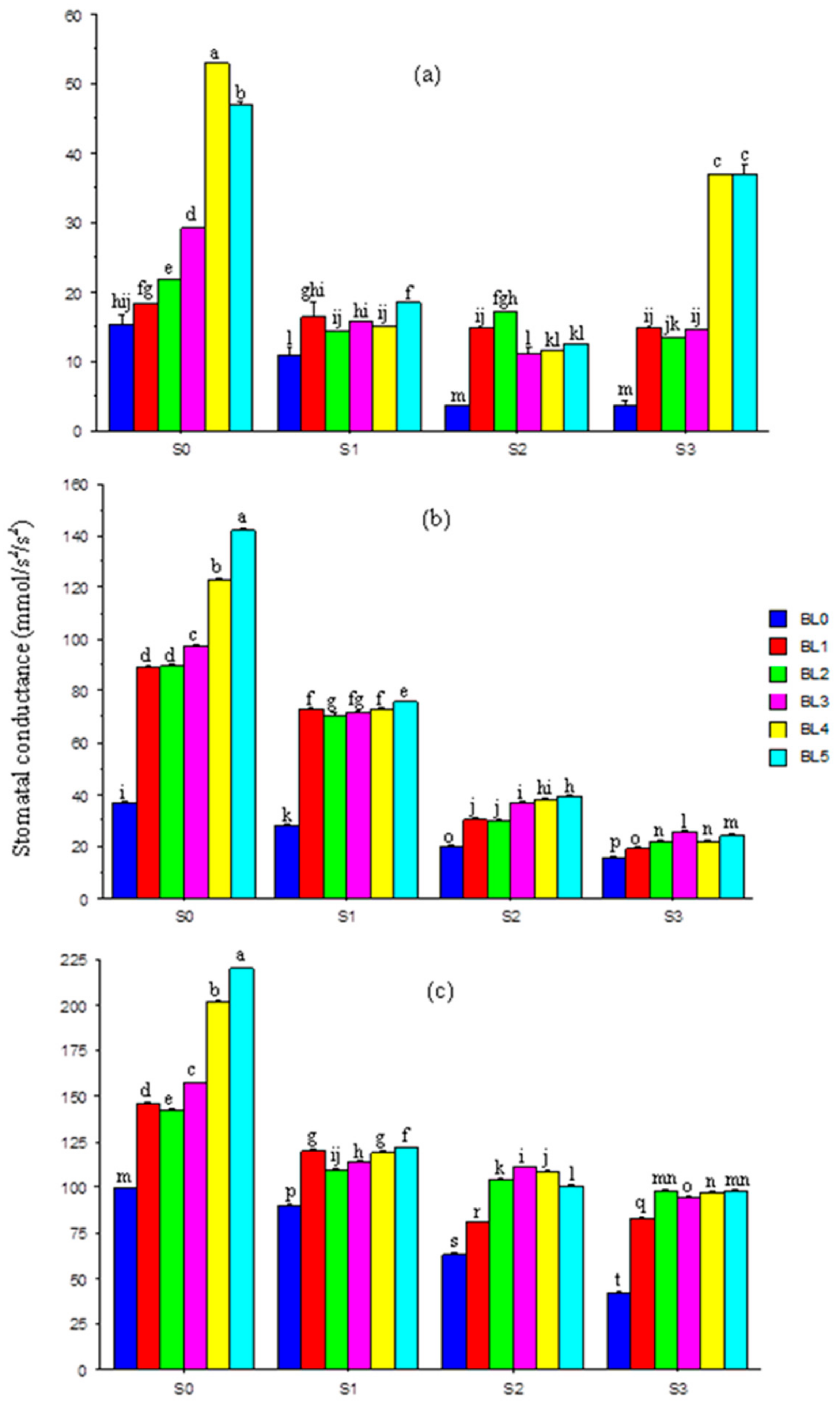
3.6. Leaf Stomatal Conductance
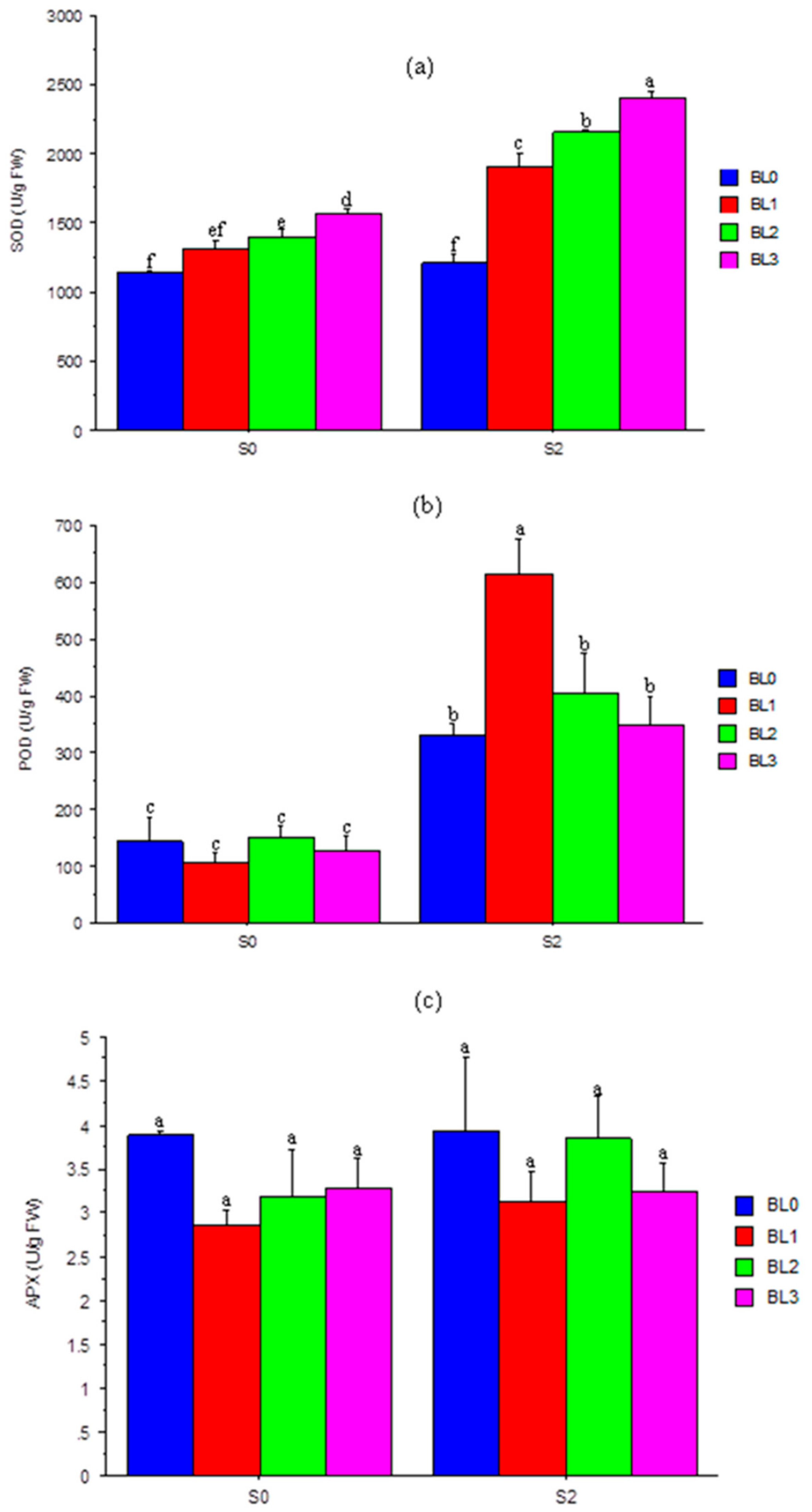
3.7. Superoxide Dismutase (SOD), Peroxidase (POD) and Ascorbate Peroxidase (APX)
3.8. Calcium [Ca2+], Potassium [K+], Magnesium [Mg2+], Sodium [Na+] and K+/Na+ Ratio
3.9. Leaf Dry Weight (LDW), Stem Dry Weight (SDW), Harvest Index (HI), Shoot-to-Root Ratio (S:R) and Protein Concentration
3.10. Number of Branches/Plant, Days to 50% Flowering and Podding, Number of Pods and Seeds/Plant
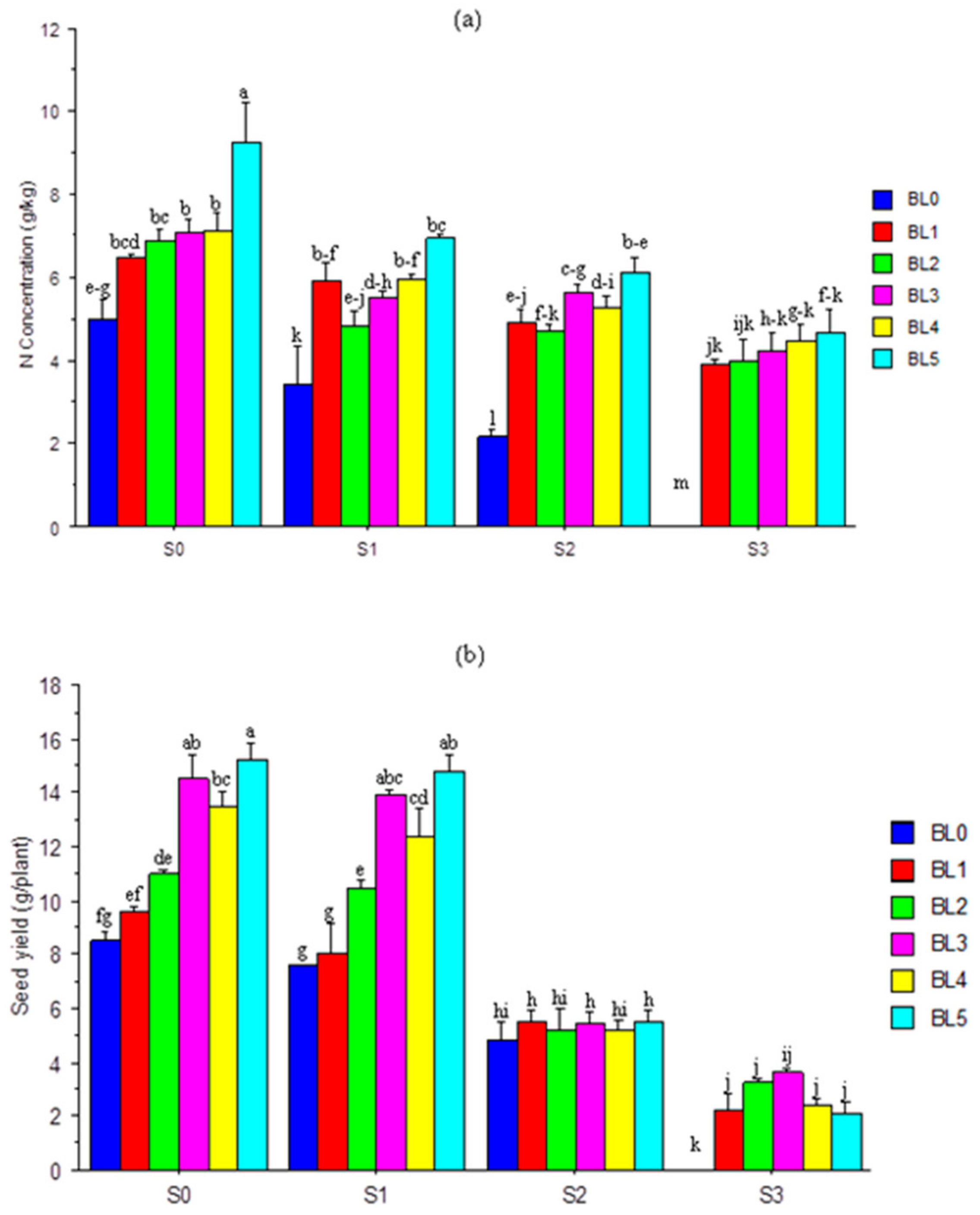
3.11. Nitrogen (N) Concentration and Seed Yield
4. Discussion
4.1. Plant Growth, Harvest Index and Biomass
4.2. Stomatal Conductance, Chlorophyll Concentration and Photosynthetically Active Radiation
4.3. Relative Water Content in Leaves
4.4. Superoxide Dismutase (SOD) Activity
4.5. Peroxidase (POD) Activity
4.6. Ascorbate Peroxidase (APX) Activity
4.7. Mineral [Calcium (Ca2+), Potassium (K+), Magnesium (Mg2+), Sodium (Na+)] Contents and K+/Na+ Ratio
4.8. Protein and Nitrogen Concentrations
4.9. Seed Yield
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Hussain, M.I.; Lyra, D.A.; Farooq, M.; Nikoloudakis, N.; Khalid, N. Salt and drought stresses in safflower: A review. Agron. Sustain. Dev. 2016, 36, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant. Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization Global Network for Integrated Soil Management for Sustainable Use of Salt-Affected Soils; FAO: Rome, Italy, 2005.
- Tian, X.; He, M.; Wang, Z.; Zhang, J.; Song, Y.; He, Z.; Dong, Y. Application of nitric oxide and calcium nitrate enhances tolerance of wheat seedlings to salt stress. Plant. Growth Regul. 2015, 77, 343–356. [Google Scholar] [CrossRef]
- Kong, X.; Luo, Z.; Dong, H.; Eneji, A.E.; Li, W. H2O2 and ABA signaling are responsible for the increased Na+efflux and water uptake inGossypium hirsutumL. roots in the non-saline side under non-uniform root zone salinity. J. Exp. Bot. 2016, 67, 2247–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohsin, S.M.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Fujita, M. Exogenous Tebuconazole and Trifloxystrobin Regulates Reactive Oxygen Species Metabolism Toward Mitigating Salt-Induced Damages in Cucumber Seedling. Plants 2019, 8, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geissler, N.; Hussin, S.; Koyro, H.-W. Interactive effects of NaCl salinity and elevated atmospheric CO2 concentration on growth, photosynthesis, water relations and chemical composition of the potential cash crop halophyte Aster tripolium L. Environ. Exp. Bot. 2009, 65, 220–231. [Google Scholar] [CrossRef]
- Zheng, Q.; Liu, J.; Liu, R.; Wu, H.; Jiang, C.; Wang, C.; Guan, Y. Temporal and spatial distributions of sodium and polyamines regulated by brassinosteroids in enhancing tomato salt resistance. Plant. Soil 2015, 400, 147–164. [Google Scholar] [CrossRef]
- Hu, W.-H.; Yan, X.-H.; Xiao, Y.-A.; Zeng, J.-J.; Qi, H.-J.; Ogweno, J.O. 24-Epibrassinosteroid alleviate drought-induced inhibition of photosynthesis in Capsicum annuum. Sci. Hortic. 2013, 150, 232–237. [Google Scholar] [CrossRef]
- Lv, J.; Zong, X.-F.; Ahmad, A.S.; Wu, X.; Wu, C.; Li, Y.-P.; Wang, S.-G. Alteration in morpho-physiological attributes of Leymus chinensis (Trin.) Tzvelev by exogenous application of brassinolide under varying levels of drought stress. Chil. J. Agric. Res. 2020, 80, 61–71. [Google Scholar] [CrossRef]
- Otie, V.; Ping, A.; Eneji, E. Interactive effect of brassinolide and lime on growth and yield of maize (Zea mays L.) on acid soils of South-East Nigeria. Commun. Soil Sci. Plant. Anal. 2018, 49, 2918–2931. [Google Scholar] [CrossRef]
- Otie, V.; Ping, A.; Ibrahim, A.; Eneji, E. Plant Growth Regulator-Brassinolide for Mitigating Field Waterlogging Stress on Maize. Int. J. Plant. Soil Sci. 2019, 30, 1–14. [Google Scholar] [CrossRef]
- Otie, V.; Ping, A.; Udo, I.; Eneji, E. Brassinolide effects on maize (Zea mays L.) growth and yield under waterlogged conditions. J. Plant. Nutr. 2019, 42, 954–969. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhu, X.H.; Ding, H.D.; Yang, S.J.; Chen, Y.Y. Foliar application of 24-epibrassinolide alleviates high-temperature-induced inhibition of photosynthesis in seedlings of two melon cultivars. Photosynth 2013, 51, 341–349. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Ahmed, M.; Mir, B.A.; Yusuf, M.; Khan, T.A. 24-Epibrassinolide mitigates the adverse effects of manganese induced toxicity through improved antioxidant system and photosynthetic attributes in Brassica juncea. Environ. Sci. Pollut. Res. 2015, 22, 11349–11359. [Google Scholar] [CrossRef]
- Karlidag, H.; Yildirim, E.; Turan, M. Role of 24-epibrassinolide in mitigating the adverse effects of salt stress on stomatal conductance, membrane permeability, and leaf water content, ionic composition in salt stressed strawberry (Fragaria×ananassa). Sci. Hortic. 2011, 130, 133–140. [Google Scholar] [CrossRef]
- Jin-Huan, L.; Anjum, S.A.; Mei-Ru, L.; Jian-Hang, N.; Ran, W.; Ji-Xuan, S.; Jun, L.; Xue-Feng, Z.; Ashraf, U.; San-Gen, W. Modulation of morpho-physiological traits of Leymus chinensis (Trin.) through exogenous appli-cation of brassinolide under salt stress. J. Anim. Plant. Sci. 2015, 25, 1055–1062. [Google Scholar]
- Maas, E.V. Crop tolerance to saline sprinkling water. Plant. Soil 1985, 89, 273–284. [Google Scholar] [CrossRef]
- An, P.; Inanaga, S.; Kafkafi, U.; Lux, A.; Sugimoto, Y. Different Effect of Humidity on Growth and Salt Tolerance of Two Soybean Cultivars. Biol. Plant. 2001, 44, 405–410. [Google Scholar] [CrossRef]
- Inanaga, S.; Cohen, Y.; Kafkafi, U.; An, P.; Sugimoto, Y. Salt tolerance in two soybean cultivars. J. Plant. Nutr. 2002, 25, 407–423. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Ghanehpoor, S.; Mohammadi-Nasab, D. Effects of water limitation on growth and grain filling of faba bean cultivars. J. Food Agric. Environ. 2009, 7, 442–447. [Google Scholar]
- Katerji, N.; van Hoorn, J.; Hamdy, A.; Mastrorilli, M. Salinity effect on crop development and yield, analysis of salt tolerance according to several classification methods. Agric. Water Manag. 2003, 62, 37–66. [Google Scholar] [CrossRef]
- Sharma, I.; Ching, E.; Saini, S.; Bhardwaj, R.; Pati, P.K. Exogenous application of brassinosteroid offers tolerance to salinity by altering stress responses in rice variety Pusa Basmati-1. Plant. Physiol. Biochem. 2013, 69, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Derevyanchuk, M.; Litvinovskaya, R.; Khripach, V.; Martinec, J.; Kravets, V. Effect of 24-epibrassinolide on Arabidopsis thaliana alternative respiratory pathway under salt stress. Acta Physiol. Plant. 2015, 37, 1–10. [Google Scholar] [CrossRef]
- Eneji, A.E.; Yamamoto, S.; Honna, T. Rice growth and nutrient uptake as affected by livestock manure in four Japanese soils. J. Plant. Nutr. 2001, 24, 333–343. [Google Scholar] [CrossRef]
- Fujiyama, H.; Nagai, T. Studies on Improvement of Nutrient and Water Supply in Crop Cultivation on Sand Dune Soil. Soil Sci. Plant. Nutr. 1987, 33, 461–470. [Google Scholar] [CrossRef] [Green Version]
- New Books. J. Agric. Food Chem. 1959, 7, 138. [CrossRef]
- Food and Agriculture Organization; World Health Organization Energy and Protein Requirement. Report of FAO/WHO Nutritional Meeting Series, No. 52; FAO: Rome, Italy, 1973. [Google Scholar]
- Smart, R.E.; Bingham, G.E. Rapid Estimates of Relative Water Content. Plant. Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Itam, M.; Abdelrahman, M.; Yamasaki, Y.; Mega, R.; Gorafi, Y.; Akashi, K.; Tsujimoto, H. Aegilops Tauschii Introgressions Improve Physio-Biochemical Traits and Metabolite Plasticity in Bread Wheat under Drought Stress. Agronomy 2020, 10, 1588. [Google Scholar] [CrossRef]
- Fujii, S.; Saka, H. Distribution of Assimilates to Each Organ in Rice Plants Exposed to a Low Temperature at the Ripening Stage, and the Effect of Brassinolide on the Distribution. Plant. Prod. Sci. 2001, 4, 136–144. [Google Scholar] [CrossRef]
- Kumari, A.; Parida, A.K. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of the halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen alleviates salinity toxicity in Solanum lycopersicum seedlings by regulating ROS homeostasis. Plant. Physiol. Biochem. 2019, 141, 466–476. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant. Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, V.P.; Lima, M.D.R.; Da Silva, B.R.S.; Batista, B.L.; Lobato, A.K.D.S. Brassinosteroids Confer Tolerance to Salt Stress in Eucalyptus urophylla Plants Enhancing Homeostasis, Antioxidant Metabolism and Leaf Anatomy. J. Plant. Growth Regul. 2019, 38, 557–573. [Google Scholar] [CrossRef]
- Fàbregas, N.; Caño-Delgado, A.I. Turning on the microscope turret: A new view for the study of brassinosteroid signaling in plant development. Physiol. Plant. 2014, 151, 172–183. [Google Scholar] [CrossRef]
- Kobraei, S.; Etminan, A.; Mohammadi, R.; Kobraee, S. Effects of drought stress on yield and yield components of soybean. Anals Biol. Res. 2011, 2, 504–509. [Google Scholar]
- Esechie, H.A.; Al-Barhi, B.; Al-Gheity, S.; Al-Khanjari, S. Root andshoot growth in salinity-stressed alfalfa in response tonitrogen source. J. Plant. Nutr. 2002, 25, 2559–2569. [Google Scholar] [CrossRef]
- Wani, A.S.; Tahir, I.; Ahmad, S.S.; Dar, R.A.; Nisar, S. Efficacy of 24-epibrassinolide in improving the nitrogen metabolism and antioxidant system in chickpea cultivars under cadmium and/or NaCl stress. Sci. Hortic. 2017, 225, 48–55. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Singh, R.; Thukral, A.K.; Bhardwaj, R. Effect of seed pre-soaking with 24-epibrassinolide on growth and photosynthetic parameters of Brassica juncea L. in imidacloprid soil. Ecotoxicol. Environ. Saf. 2016, 133, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mohsin, S.M.; Fujita, A.M. Quercetin Mediated Salt Tolerance in Tomato through the Enhancement of Plant Antioxidant Defense and Glyoxalase Systems. Plants 2019, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.J.; Anjum, S.A.; Wang, L.; Song, J.X.; Zong, X.F.; Lv, J.; Zohaib, A.; Ali, I.; Yan, R.; Zhang, Y.; et al. Effect of foliar application of brassinolide on photosynthesis and chlorophyll fluorescence traits of Leymus chinensis under varying levels of shade. Photosynth 2018, 56, 873–883. [Google Scholar] [CrossRef]
- Islam, M.T.; Jahan, N.A.; Sen, A.K.; Pramanik, M.H.R. Effects of salinity on morpho-physiological attributes and yield of lentil genotypes. Int. J. Sustain. Crop. Prod. 2012, 1, 12–18. [Google Scholar]
- Taf, V.D.; Kou, J.K.; Tchi, L.M.; Nand, B.A.; Akoa, A. Effects of Salinity Stress on Growth, Ions Partitioning and Yield of Some Cowpea (Vigna unguiculata L. Walp.) Cultivars. Int. J. Bot. 2009, 5, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Mannan, A.M.; Abdul, K.M.; Moynul, H.M.; Abdul, K.Q.; Higuchi, H.; Nawata, E. Response of Soybean to Salinity: II. Growth and Yield of Some Selected Genotypes. Trop. Agric. Dev. 2013, 57, 31–40. [Google Scholar] [CrossRef]
- Mannan, M.A.; Karim, M.A.; Haque, M.M.; Khaliq, Q.A.; Higuchi, H.; Nawata, E. Response of Soybean to Salinity: III. Water Status and Accumulation of Mineral Ions. Trop. Agric. Dev. 2013, 57, 41–48. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Chen, Y.; Li, R. Effects of brassinolide on fruit growth and quality of pitaya. J. South. Agric. 2013, 44, 1150–1153. [Google Scholar]
- Piñero, M.C.; Houdusse, F.; Garcia-Mina, J.M.; Garnica, M.; Del Amor, F.M. Regulation of hormonal responses of sweet pepper as affected by salinity and elevated CO2 concentration. Physiol. Plant. 2013, 151, 375–389. [Google Scholar] [CrossRef]
- González, D.; Fuentes, S.; Serna, L. Interactions among gibberellins, brassinosteroids and genes regulate sto-matal development in the arabidopsis hypocotyl. Int. J. Dev. Biol. 2017, 61, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, L.R.; Da Silva, B.R.S.; Pedron, T.; Batista, B.L.; Lobato, A.K.D.S. 24-Epibrassinolide Improves Root Anatomy and Antioxidant Enzymes in Soybean Plants Subjected to Zinc Stress. J. soil Sci. Plant. Nutr. 2019, 20, 105–124. [Google Scholar] [CrossRef]
- Hayat, S.; Hasan, S.A.; Yusuf, M.; Hayat, Q.; Ahmad, A. Effect of 28-homobrassinolide on photosynthesis, fluorescence and antioxidant system in the presence or absence of salinity and temperature in Vigna radiata. Environ. Exp. Bot. 2010, 69, 105–112. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant. Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ghobadi, M.; Taherabadi, S.; Ghobadi, M.-E.; Mohammadi, G.-R.; Jalali-Honarmand, S. Antioxidant capacity, photosynthetic characteristics and water relations of sunflower (Helianthus annuus L.) cultivars in response to drought stress. Ind. Crop. Prod. 2013, 50, 29–38. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium Supplementation Improves Na+/K+ Ratio, Antioxidant Defense and Glyoxalase Systems in Salt-Stressed Rice Seedlings. Front. Plant. Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [Green Version]
- Xuan, T.D.; Khang, D.T. Effects of Exogenous Application of Protocatechuic Acid and Vanillic Acid to Chlorophylls, Phenolics and Antioxidant Enzymes of Rice (Oryza sativa L.) in Submergence. Molecules 2018, 23, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological Mechanism of Enhancing Salt Stress Tolerance of Perennial Ryegrass by 24-Epibrassinolide. Front. Plant. Sci. 2017, 8, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Gao, H.; Zheng, Q.; Wang, C.; Wang, X.; Wang, Q. Effects of 24-epibrassinolide on plant growth, osmotic regulation and ion homeostasis of salt-stressed canola. Plant. Biol. 2013, 16, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Morales, L.M.M.; Senn, M.E.; Grozeff, G.E.G.; Fanello, D.D.; Carrión, C.A.; Núñez, M.; Bishop, G.J.; Bartoli, C.G. Impact of brassinosteroids and ethylene on ascorbic acid accumulation in tomato leaves. Plant. Physiol. Biochem. 2014, 74, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, F.R.; Lima, J.P.M.S.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. Roots and leaves display contrasting oxidative response during salt stress and recovery in cowpea. J. Plant. Physiol. 2007, 164, 591–600. [Google Scholar] [CrossRef]
- Yadava, P.; Kaushal, J.; Gautam, A.; Parmar, H.; Singh, I. Physiological and Biochemical Effects of 24-Epibrassinolide on Heat-Stress Adaptation in Maize (Zea mays L.). Nat. Sci. 2016, 8, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R.C. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol. Plant. 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Zhou, Y.; Diao, M.; Chen, X.; Cui, J.; Pang, S.; Li, Y.; Hou, C.; Liu, H.-Y. Application of exogenous glutathione confers salinity stress tolerance in tomato seedlings by modulating ions homeostasis and polyamine metabolism. Sci. Hortic. 2019, 250, 45–58. [Google Scholar] [CrossRef]
- Mekawy, A.M.M.; Abdelaziz, M.N.; Ueda, A. Apigenin pretreatment enhances growth and salinity tolerance of rice seedlings. Plant. Physiol. Biochem. 2018, 130, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Akhtar, K.; Sarwar, G. Role of the rooting system in salt tolerance potential of different guar accessions. Agron. Sustain. Dev. 2005, 25, 243–249. [Google Scholar] [CrossRef]
- Sarwar, G.; Ashraf, M.; Naeem, M. Genetic variability of some primitive bread wheat varieties to salt tolerance. Pak. J. Bot. 2003, 35, 771–777. [Google Scholar]
- Ali, B.; Hayat, S.; Fariduddin, Q.; Ahmad, A. 24-Epibrassinolide protects against the stress generated by salinity and nickel in Brassica juncea. Chemosphere 2008, 72, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Shawky, B.T. 24-Epibrassinolide alleviates salt-induced inhibition of productivity by increasing nutrients and compatible solutes accumulation and enhancing antioxidant system in wheat (Triticum aestivum L.). Acta Physiol. Plant. 2013, 35, 729–740. [Google Scholar] [CrossRef]
- Azhar, N.; Su, N.; Shabala, L.; Shabala, S. Exogenously Applied 24-Epibrassinolide (EBL) Ameliorates Detrimental Effects of Salinity by Reducing K+ Efflux via Depolarization-Activated K+ Channels. Plant. Cell Physiol. 2017, 58, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Wang, W.; Hu, G.; Chen, W.; Zhuge, Y.; Wang, Z.; He, M. Role of Exogenous 24-Epibrassinolide in Enhancing the Salt Tolerance of Wheat Seedlings. J. Soil Sci. Plant. Nutr. 2017, 17, 554–569. [Google Scholar] [CrossRef] [Green Version]
- Elzeiny, H.A.; Abou, L.B.; Gaballah, M.S.; Khalil, S. Anti-transpirant application to sesame plant for salinity stress augmentation. Res. J. Agric. Biol. Sci. 2007, 3, 950–959. [Google Scholar]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- Divi, U.K.; Rahman, T.; Krishna, P. Gene expression and functional analyses in brassinosteroid-mediated stress tolerance. Plant. Biotechnol. J. 2015, 14, 419–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xia, X.; Yu, G.; Wang, J.; Wu, J.; Wang, M.; Yang, Y.; Shi, K.; Yu, Y.; Chen, Z.; et al. Brassinosteroids play a critical role in the regulation of pesticide metabolism in crop plants. Sci. Rep. 2015, 5, 9018. [Google Scholar] [CrossRef]
- Sahni, S.; Prasad, B.D.; Liu, Q.; Grbic, V.; Sharpe, A.; Singh, S.P.; Krishna, P. Overexpression of the brassinosteroid biosynthetic gene DWF4 in Brassica napus simultaneously increases seed yield and stress tolerance. Sci. Rep. 2016, 6, 28298. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources of Variance | Plant Height (cm) | Number of Leaves | Leaf Area Index (LAI) | Specific Leaf Area (cm2/g) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S × BL | 2 WAS | 4 WAS | 6 WAS | 8 WAS | 2 WAS | 4 WAS | 6 WAS | 8 WAS | 4WAS | 6WAS | 8WAS | 6WAS |
| S0BL0 | 14.33 efg | 21.00 j | 40.15 ef | 48.90 bcd | 7.00 de | 17.33 j–m | 30.33 l | 38.67 gh | 2.91 gh | 4.64 fgh | 5.67 def | 152.40 l |
| S1 BL0 | 13.67 efg | 17.99 k | 41.10 ef | 42.73 e–i | 10.00 b | 13.67 n | 30.00 l | 36.67 h–k | 2.40 hi | 3.81 ghi | 4.45 ghi | 142.40 mn |
| S2 BL0 | 14.00 efg | 18.25 k | 30.82 m | 37.87 i | 9.00 b–e | 10.46 o | 21.00 l | 31.67 klm | 2.13 i | 2.78 ij | 3.12 j | 133.50 p |
| S3 BL0 | 11.67 g | 13.34 l | 20.10 n | 24.57 j | 8.00 cde | 6.00 p | 12.00 n | 20.00 p | 1.07 j | 2.38 j | 3.13 j | 119.50 q |
| S0 BL1 | 13.00 fg | 34.70 c | 43.37 d | 66.53 a | 8.00 cde | 23.33 cd | 58.00 e | 69.33 c | 4.36 dc | 6.38 bcd | 7.70 c | 208.10 de |
| S1 BL1 | 13.00 fg | 34.94 c | 41.50 e | 44.33 d–h | 14.00 a | 19.33 f–l | 39.00 gh | 41.67 fg | 3.95 ef | 5.59 def | 5.91 def | 196.80 g |
| S2 BL1 | 16.00 cde | 31.52 de | 38.07 gh | 40.57 f–i | 8.00 cde | 17.00 j–n | 26.00 j | 34.33 h–l | 2.75 ghi | 4.73 efg | 5.31 efg | 166.80 k |
| S3 BL1 | 17.00 bcd | 23.62 i | 32.53 l | 44.87 d–g | 8.00 cde | 19.67 e–k | 20.00 l | 33.00 i–m | 2.67 ghi | 3.59 g–j | 4.52 ghi | 137.30 o |
| S0 BL2 | 12.00 g | 40.13 b | 46.70 c | 66.73 a | 5.00 f | 24.00 c | 64.00 d | 71.33 c | 3.70 ef | 6.40 bcd | 8.64 b | 209.30 d |
| S1 BL2 | 14.00 efg | 31.26 de | 41.77 e | 45.57 d–g | 8.00 cde | 20.33 d–j | 37.00 h | 46.37 e | 3.86 ef | 5.29 def | 5.78 def | 195.90 g |
| S2 BL2 | 14.00 efg | 28.98 fg | 34.63 k | 38.70 hi | 9.00 b–e | 17.00 i–n | 27.00 j | 33.00 i–m | 2.90 gh | 5.47 def | 5.42 ef | 151.70 l |
| S3 BL2 | 12.00 g | 25.14 hi | 39.47 fg | 42.87 e–i | 9.00 b–e | 21.00 c–f | 18.00 m | 24.67 no | 2.61 ghi | 4.55 fgh | 5.01 f–i | 135.50 op |
| S0 BL3 | 16.00 cde | 38.77 b | 46.80 c | 69.40 a | 10.00 b | 28.00 b | 69.00 c | 72.33 c | 4.71 cd | 6.88 bc | 9.07 b | 215.50 c |
| S1 BL3 | 8.00 h | 32.04 de | 43.63 d | 46.77 cde | 8.00 cde | 21.67 c–f | 39.00 gh | 46.00 ef | 3.30 fg | 5.94 cde | 6.10 de | 196.40 g |
| S2 BL3 | 15.00 def | 28.34 g | 37.33 hi | 39.87 ghi | 7.00 de | 17.00 h–n | 26.00 j | 31.33 lm | 2.93 gh | 4.75 efg | 5.13 fgh | 174.70 i |
| S3 BL3 | 13.00 fg | 28.89 fg | 36.30 ij | 39.67 ghi | 8.00 cde | 15.67 mn | 18.00 m | 22.67 op | 2.73 ghi | 3.78 g–j | 4.40 hi | 140.40 n |
| S0 BL4 | 17.00 bcd | 40.14 b | 53.57 b | 67.43 a | 8.00 cde | 28.33 b | 72.00 b | 81.67 b | 5.53 b | 7.51 b | 9.33 b | 227.30 b |
| S1 BL4 | 15.00def | 30.42 ef | 43.51 d | 51.90 bc | 8.00 cde | 22.33 c–f | 38.00 gh | 44.00 ef | 3.94 ef | 5.54 def | 6.20 de | 201.40 f |
| S2 BL4 | 12.00 g | 26.18 h | 40.77 ef | 46.10 def | 8.00 cde | 20.33 d–j | 24.00 k | 34.00 h–l | 2.58 hi | 3.35 hij | 4.25 hi | 170.70 j |
| S3 BL4 | 19.00ab | 24.78 hi | 32.28 lm | 44.17 d–h | 7.00 de | 16.33 k–m | 24.00 k | 31.67 j–m | 2.77 ghi | 3.72 ghi | 4.12 i | 141.90 mn |
| S0 BL5 | 20.33 a | 42.44 a | 56.67 a | 68.93 a | 8.00 cde | 32.00 a | 78.33 a | 88.00 a | 6.42 a | 9.25 a | 11.80 a | 233.40 a |
| S1 BL5 | 15.00 def | 33.10 cd | 43.46 d | 53.07 b | 8.00 cde | 23.00 cde | 47.00 f | 52.67 d | 5.03 bc | 5.54 def | 6.34 d | 206.20 e |
| S2 BL5 | 18.00 abc | 26.44 h | 35.63 jk | 39.93 ghi | 8.00 cde | 22.67 c–f | 23.00 k | 36.67 h–k | 2.94 gh | 3.43 hij | 4.29 hi | 188.60 h |
| S3 BL5 | 17.67 bc | 24.82 hi | 34.53 k | 40.83 f–i | 8.00 cde | 16.00 lmn | 20.00 l | 29.00 mn | 5.57 hi | 3.77 ghi | 4.25 hi | 143.40 m |
| Sources of Variance | Calcium (mg/g) | Potassium (mg/g) | Magnesium (mg/g) | Sodium (mg/g) | K+/Na+ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Salinity (S) Concentrations (mM/L) | Leaves | Stems | Roots | Leaves | Stems | Roots | Leaves | Stems | Roots | Leaves | Stems | Roots | Leaves | Stems | Roots |
| S0 | 3.00 a | 0.75 a | 0.58 a | 22.50 a | 14.88 a | 15.59 a | 1.16 a | 0.78 a | 0.65 a | 0.73 a | 0.99 c | 4.71 b | 66.76 a | 21.95 a | 5.86 a |
| S1 | 2.36 b | 0.54 b | 0.39 b | 20.39 a | 12.59 a | 10.20 b | 1.00 b | 0.70 ab | 0.30 b | 2.62 b | 10.70 b | 14.09 a | 12.30 b | 2.14 b | 0.71 b |
| S2 | 2.09 c | 0.51 bc | 0.32 b | 17.59 b | 11.22 bc | 5.55 c | 0.90 c | 0.61 bc | 0.13 c | 5.52 a | 15.47 a | 14.64 a | 7.20 b | 0.73 b | 0.38 b |
| S3 | 1.39 d | 0.47 c | 0.30 b | 12.39 c | 10.06 c | 3.81 c | 0.81 d | 0.53 c | 0.09 c | 6.29 a | 16.28 a | 14.90 a | 3.37 b | 0.62 b | 0.25 b |
| Growth stages of BL applications | |||||||||||||||
| BL0 | 1.27 d | 0.50 b | 0.26 a | 11.34 b | 7.29 c | 3.50 c | 0.84 b | 0.53 b | 0.11 c | 5.69 a | 11.65 a | 13.58 a | 8.25 b | 2.69 a | 0.31 b |
| BL1 | 2.17 c | 0.55 ab | 0.38 a | 17.63 a | 11.54 b | 8.05 b | 0.98 a | 0.66 ab | 0.31 b | 2.88 b | 11.13 a | 12.15 b | 20.56 ab | 8.22 a | 1.09 b |
| BL2 | 2.28 bc | 0.58 ab | 0.38 a | 19.29 a | 12.47 ab | 9.26 ab | 0.98 a | 0.69 a | 0.28 b | 3.93 ab | 10.97 a | 11.94 b | 22.68 ab | 4.83 a | 1.27 b |
| BL3 | 2.26 bc | 0.57 ab | 0.40 a | 19.25 a | 13.21 ab | 9.24 ab | 0.98 a | 0.69 a | 0.29 b | 4.19 ab | 11.80 a | 11.88 b | 24.58 ab | 7.23 a | 1.84 b |
| BL4 | 2.53 ab | 0.57 ab | 0.43 a | 20.75 a | 14.06 ab | 10.44 ab | 1.00 a | 0.72 a | 0.36 ab | 3.18 b | 10.54 a | 11.64 b | 28.55 a | 5.83 a | 1.78 b |
| BL5 | 2.73 a | 0.63 a | 0.53 a | 21.05 a | 14.56 a | 12.22 a | 1.03 a | 0.65 ab | 0.42 a | 2.86 b | 9.07 b | 11.30 b | 29.81 a | 9.37 a | 4.55 a |
| Sources of Variance | Leaf Dry Weight (g/Plant) | Stem Dry Weight (g/Plant) | Harvest Index (%) | Shoot to Root Ratio | Protein Concentration (%) |
|---|---|---|---|---|---|
| Salinity (S) Concentrations (mM/L) | |||||
| S0 | 8.59 a | 5.81 a | 32.86 a | 15.99 a | 39.41 a |
| S1 | 7.58 b | 5.15 b | 29.78 ab | 12.90 b | 31.15 b |
| S2 | 3.26 c | 2.37 c | 28.48 b | 10.79 c | 27.53 c |
| S3 | 2.20 d | 1.28 d | 19.06 c | 9.22 d | 20.30 d |
| Growth Stages of BL Applications | |||||
| BL0 | 2.86 e | 2.20 c | 18.84 b | 8.58 d | 15.17 c |
| BL1 | 4.80 d | 3.68 b | 27.78 a | 11.58 c | 30.35 b |
| BL2 | 5.36 cd | 3.95 ab | 29.55 a | 12.12 bc | 29.21 b |
| BL3 | 6.98 a | 4.36 a | 30.61 a | 12.68 b | 32.12 b |
| BL4 | 6.30 ab | 3.73 b | 29.95 a | 13.04 b | 32.68 b |
| BL5 | 6.13 bc | 3.98 ab | 28.54 a | 15.36 a | 38.06 a |
| S × BL | |||||
| S0 BL0 | 4.73 f | 3.85 d | 20.06 b–e | 10.93 f–i | 28.70 a |
| S1 BL0 | 3.40 fgh | 2.53 ef | 23.35 cde | 9.78 hij | 19.62 a |
| S2 BL0 | 2.09 hi | 1.75 f–j | 25.96 b–e | 8.14 j | 12.37 a |
| S3 BL0 | 1.21 i | 0.66 j | 0.00 f | 5.46 k | 0.00 a |
| S0 BL1 | 7.07 e | 5.68 abc | 30.02 a–e | 14.19 cd | 37.08 a |
| S1 BL1 | 6.54 e | 5.18 c | 31.20 a–d | 11.96 e–h | 33.88 a |
| S2 BL1 | 3.25 fgh | 2.81 ef | 29.54 a–e | 11.02 f–h | 28.17 a |
| S3 BL1 | 2.35 ghi | 1.03 h–j | 20.36 e | 9.15 ij | 22.27 a |
| S0 BL2 | 7.98 cde | 6.44 ab | 33.09 abc | 15.70 bc | 39.40 a |
| S1 BL2 | 7.49 de | 5.62 abc | 32.83 abc | 12.61 d–g | 27.60 a |
| S2 BL2 | 3.21 fgh | 2.07 e–h | 28.34 b–e | 10.93 f–i | 27.03 a |
| S3 BL2 | 2.77 ghi | 1.72 f–j | 23.93 cde | 9.25 ij | 22.84 a |
| S0 BL3 | 11.49 a | 6.57 a | 39.45 a | 17.01 b | 40.35 a |
| S1 BL3 | 9.72 b | 5.83 abc | 30.17 a–e | 13.24 de | 31.59 a |
| S2 BL3 | 3.97 fg | 3.07 de | 30.65 a–d | 10.48 ghi | 32.35 a |
| S3 BL3 | 2.73 ghi | 1.98 e–i | 22.17 de | 9.99 hij | 24.17 a |
| S0 BL4 | 10.42 ab | 6.19 abc | 35.13 ab | 16.92 b | 40.51 a |
| S1 BL4 | 8.94 bcd | 5.41 bc | 32.06 a–d | 14.06 cd | 34.26 a |
| S2 BL4 | 3.66 fgh | 2.38 efg | 28.52 b–e | 11.31 e–i | 30.26 a |
| S3 BL4 | 2.16 ghi | 0.93 ij | 24.10 cde | 9.87 hij | 25.69 a |
| S0 BL5 | 9.85 ab | 6.10 abc | 33.40 abc | 21.21 a | 50.43 a |
| S1 BL5 | 9.37 bc | 6.33 ab | 29.04 b–e | 15.74 bc | 39.96a |
| S2 BL5 | 3.37 fgh | 2.15 efg | 27.89 b–e | 12.89 def | 35.02 a |
| S3 BL5 | 1.94 hi | 1.34 g–j | 23.82 cde | 11.60 e–h | 26.84 a |
| Sources of Variance | Number of Branches/Plant | Days to 50% Flowering | Days to 50% Podding | Number of Pods/Plant | Number of Seeds/Plant |
|---|---|---|---|---|---|
| Salinity (S) Concentrations (mM/L) | |||||
| S0 | 25.17 a | 19.94 d | 35.00 c | 35.39 a | 88.22 a |
| S1 | 20.06 b | 20.78 c | 34.44 c | 24.39 b | 60.44 b |
| S2 | 16.33 c | 28.00 b | 42.00 b | 18.50 c | 46.17 c |
| S3 | 10.00 d | 34.67a | 48.67 a | 6.00 d | 13.78 d |
| Growth Stages of BL Applications | |||||
| BL0 | 13.25 d | 27.33 a | 43.25 a | 15.25 b | 35.92 b |
| BL1 | 16.08 c | 25.25 b | 38.83 c | 21.58 a | 53.67 a |
| BL2 | 18.42 b | 25.33 b | 39.00 bc | 21.83 a | 54.33 a |
| BL3 | 19.25 ab | 25.75 b | 39.75 bc | 22.58 a | 56.25 a |
| BL4 | 20.00 ab | 25.42 b | 39.33 bc | 21.92 a | 54.67 a |
| BL5 | 20.33 a | 26.00 b | 40.00 b | 23.25 a | 58.08 a |
| S × BL | |||||
| S0 BL0 | 19.67 a | 23.00 c | 44.67 b | 25.53 a | 63.00 a |
| S1 BL0 | 16.00 a | 23.67 c | 37.67 d | 20.67a | 50.00 a |
| S2 BL0 | 13 33 a | 27.67 b | 41.67 c | 12.33 a | 30.67 a |
| S3 BL0 | 4.00 a | 35.00 a | 49.00 a | 2.67 a | 0.00 a |
| S0 BL1 | 23.00 a | 19.00 e | 32.67fg | 36.33 a | 90.67 a |
| S1 BL1 | 16.67 a | 19.00 e | 31.67 g | 24.67 a | 61.33 a |
| S2 BL1 | 15.33 a | 28.67 b | 42.67 c | 20.33 a | 50.67 a |
| S3 BL1 | 9.33 a | 34.33 a | 48.33 a | 5.00 a | 12.00 a |
| S0 BL2 | 25.00 a | 19.00 e | 32.67 fg | 36.67 a | 91.33 a |
| S1 BL2 | 20.67 a | 19.33 e | 32.33 fg | 25.33 a | 63.00 a |
| S2 BL2 | 16.67 a | 28.33 b | 42.33 c | 19.33 a | 48.33 a |
| S3 BL2 | 11.33 a | 34.67 a | 48.67 a | 6.00 a | 14.67 a |
| S0 BL3 | 27.33 a | 20.00 de | 33.67 efg | 38.33 a | 95.67 a |
| S1 BL3 | 22.33 a | 20.67 de | 35.00 e | 26.33 a | 65.67 a |
| S2 BL3 | 16.33 a | 27.33 b | 41.33 c | 18.33a | 45.67 a |
| S3 BL3 | 11.00 a | 35.00 a | 49.00 a | 7.33 a | 18.00 a |
| S0 BL4 | 28.00 a | 19.00 e | 32.67 fg | 34.67 a | 86.67 a |
| S1 BL4 | 22.33 a | 20.33 de | 34.33 ef | 24.67 a | 61.33 a |
| S2 BL4 | 17.67 a | 28.33 b | 42.33 c | 20.00 a | 50.00 a |
| S3 BL4 | 12.00 a | 34.00 a | 48.00 a | 8.33 a | 20.67 a |
| S0 BL5 | 28.00 a | 19.67 de | 33.67 efg | 41.00 a | 102.00 a |
| S1 BL5 | 22.33 a | 21.67 cd | 35.67 e | 24.67 a | 61.33 a |
| S2 BL5 | 18.67 a | 27.67 b | 41.67 c | 20.67 a | 51.67 a |
| S3 BL5 | 12.33 a | 35.00 a | 49.00 a | 6.67 a | 17.33 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otie, V.; Udo, I.; Shao, Y.; Itam, M.O.; Okamoto, H.; An, P.; Eneji, E.A. Salinity Effects on Morpho-Physiological and Yield Traits of Soybean (Glycine max L.) as Mediated by Foliar Spray with Brassinolide. Plants 2021, 10, 541. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030541
Otie V, Udo I, Shao Y, Itam MO, Okamoto H, An P, Eneji EA. Salinity Effects on Morpho-Physiological and Yield Traits of Soybean (Glycine max L.) as Mediated by Foliar Spray with Brassinolide. Plants. 2021; 10(3):541. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030541
Chicago/Turabian StyleOtie, Victoria, Idorenyin Udo, Yang Shao, Michael O. Itam, Hideki Okamoto, Ping An, and Egrinya A. Eneji. 2021. "Salinity Effects on Morpho-Physiological and Yield Traits of Soybean (Glycine max L.) as Mediated by Foliar Spray with Brassinolide" Plants 10, no. 3: 541. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030541