
Microscopic and Transcriptomic Analysis of Pollination Processes in Self-Incompatible Taraxacum koksaghyz


Abstract
:1. Introduction
2. Results
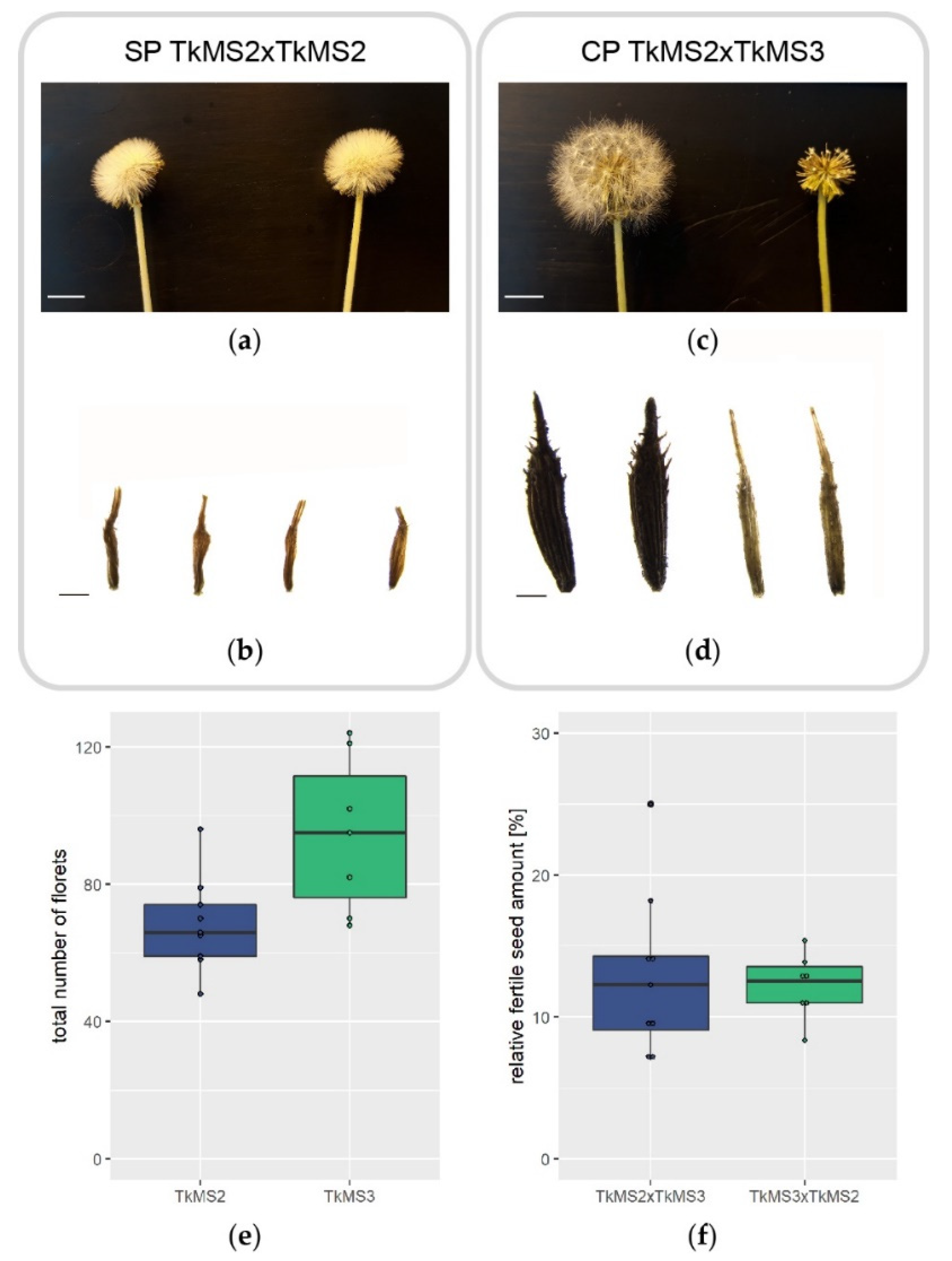
2.1. Seed Development after Self-Pollination and Cross-Pollination of T. koksaghyz
2.2. Evaluation of Pollen Tube Growth by Microscopy
2.3. Transcriptomic Analysis of CP and SP
2.3.1. Filtering and Venn Diagrams
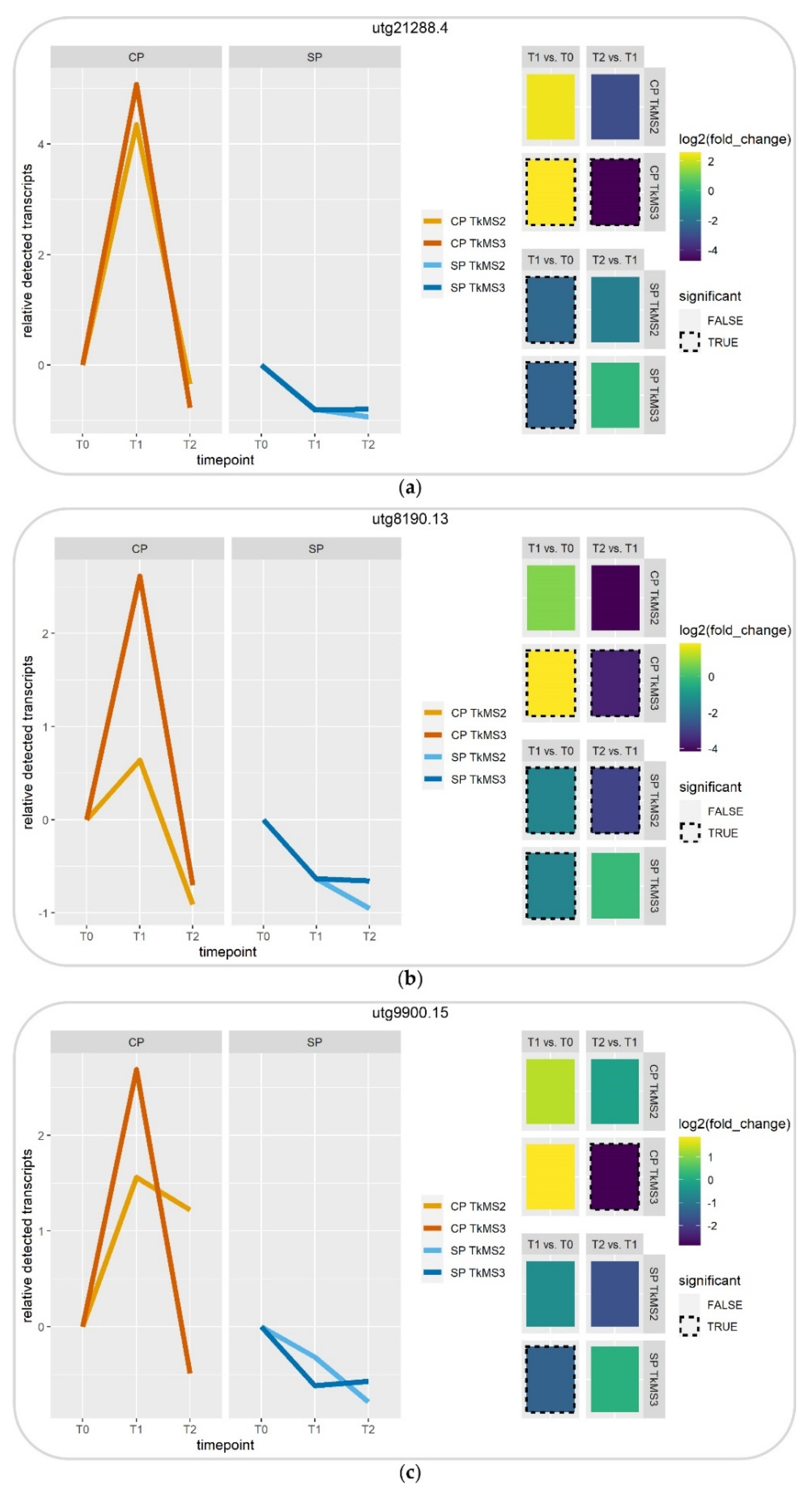
2.3.2. Digital Gene Expression Profile of Selected DEGs
2.3.3. Gene Ontology Analysis of DEGs in the T1 vs. T0 and T2 vs. T1 Comparisons
3. Discussion
4. Materials and Methods
4.1. Plant Material, Cultivation and Pollination
4.2. Confocal Laser Scanning Microscopy (CLSM)
4.3. RNA Extraction, RNA-Seq and Transcriptome Analysis
4.4. Differential Expression Analysis
4.5. GO Enrichment Analysis and Venn Diagrams
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, A.M.; Lexer, C.; Hiscock, S.J. Comparative Analysis of Pistil Transcriptomes Reveals Conserved and Novel Genes Expressed in Dry, Wet, and Semidry Stigmas. Plant Physiol. 2010, 154, 1347–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, M.M.; Good-Avila, S.V. Macrophylogenetic analyses of the gain and loss of self-incompatibility in the Asteraceae. New Phytol. 2007, 173, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J.B.; Poirier, Y. Guayule and Russian dandelion as alternative sources of natural rubber. Crit. Rev. Biotechnol. 2007, 27, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Stolze, A.; Wanke, A.; van Deenen, N.; Geyer, R.; Prüfer, D.; Schulze Gronover, C. Development of rubber-enriched dandelion varieties by metabolic engineering of the inulin pathway. Plant Biotechnol. J. 2017, 15, 740–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Xu, X.; Ruan, J.; Liu, S.; Wu, S.; Shao, X.; Wang, X.; Gan, L.; Qin, B.; Yang, Y.; et al. Genome analysis of Taraxacum kok-saghyz Rodin provides new insights into rubber biosynthesis. Natl. Sci. Rev. 2018, 5, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Niephaus, E.; Müller, B.; van Deenen, N.; Lassowskat, I.; Bonin, M.; Finkemeier, I.; Prüfer, D.; Schulze Gronover, C. Uncovering mechanisms of rubber biosynthesis in Taraxacum koksaghyz—Role of cis-prenyltransferase-like 1 protein. Plant J. 2019, 100, 591–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieghaus, A.; Prüfer, D.; Schulze Gronover, C. Loss of function mutation of the Rapid Alkalinization Factor (RALF1)-like peptide in the dandelion Taraxacum koksaghyz entails a high-biomass taproot phenotype. PLoS ONE 2019, 14, e0217454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzberger, M.; Hahn, T.; Zibek, S.; Schiemann, J.; Thiele, K. Seasonal pattern of biomass and rubber and inulin of wild Russian dandelion (Taraxacum koksaghyz L. Rodin) under experimental field conditions. Eur. J. Agron. 2016, 80, 66–77. [Google Scholar] [CrossRef]
- Eggert, M.; Schiemann, J.; Thiele, K. Yield performance of Russian dandelion transplants (Taraxacum koksaghyz L. Rodin) in flat bed and ridge cultivation with different planting densities. Eur. J. Agron. 2018, 93, 126–134. [Google Scholar] [CrossRef]
- Nowicki, M.; Zhao, Y.; Boggess, S.L.; Fluess, H.; Payá-Milans, M.; Staton, M.E.; Houston, L.C.; Hadziabdic, D.; Trigiano, R.N. Taraxacum kok-saghyz (rubber dandelion) genomic microsatellite loci reveal modest genetic diversity and cross-amplify broadly to related species. Sci. Rep. 2019, 9, 1915. [Google Scholar] [CrossRef]
- Mogie, M.; Ford, H. Sexual and asexual Taraxacum species. Biol. J. Linn. Soc. 1988, 35, 155–168. [Google Scholar] [CrossRef]
- Kirschner, J.; Štěpánek, J.; Černý, T.; De Heer, P.; van Dijk, P.J. Available ex situ germplasm of the potential rubber crop Taraxacum koksaghyz belongs to a poor rubber producer, T. brevicorniculatum (Compositae–Crepidinae). Genet. Resour. Crop Evol. 2013, 60, 455–471. [Google Scholar] [CrossRef]
- Goulet, B.E.; Roda, F.; Hopkins, R. Hybridization in Plants: Old Ideas, New Techniques. Plant Physiol. 2017, 173, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Hinata, K.; Isogai, A.; Isuzugawa, K. Manipulation of sporophytic self-incompatibility in plant breeding. In Genetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants; Springer: Dordrecht, The Netherlands, 1994; pp. 102–115. [Google Scholar]
- Fick, G.N.; Miller, J.F. Sunflower Breeding. Sunflower Technol. Prod. 1997, 35, 395–439. [Google Scholar]
- Elleman, C.J.; Dickinson, H.G. Commonalities between pollen/stigma and host/pathogen interactions: Calcium accumulation during stigmatic penetration by brassica oleracea pollen tubes. Sex. Plant Reprod. 1999, 12, 194–202. [Google Scholar] [CrossRef]
- Kessler, S.A.; Shimosato-Asano, H.; Keinath, N.F.; Wuest, S.E.; Ingram, G.; Panstruga, R.; Grossniklaus, U. Conserved molecular components for pollen tube reception and fungal invasion. Science 2010, 330, 968–971. [Google Scholar] [CrossRef] [PubMed]
- Iwano, M.; Takayama, S. Self/non-self discrimination in angiosperm self-incompatibility. Curr. Opin. Plant Biol. 2012, 15, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.A.; Grossniklaus, U. She’s the boss: Signaling in pollen tube reception. Curr. Opin. Plant Biol. 2011, 14, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscock, S.J. Genetic control of self-incompatibility in Senecio squalidus L. (Asteraceae): A successful colonizing species. Heredity (Edinb) 2000, 85, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascallares, M.; Setzes, N.; Marchetti, F.; López, G.A.; Distéfano, A.M.; Cainzos, M.; Zabaleta, E.; Pagnussat, G.C. A Complex Journey: Cell Wall Remodeling, Interactions, and Integrity during Pollen Tube Growth. Front. Plant Sci. 2020, 11, 599247. [Google Scholar] [CrossRef] [PubMed]
- Chapman, L.A.; Goring, D.R. Pollen-pistil interactions regulating successful fertilization in the Brassicaceae. J. Exp. Bot. 2010, 61, 1987–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, D.J. The extracellular pollen coat in members of the Brassicaceae: Composition, biosynthesis, and functions in pollination. Protoplasma 2006, 228, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Hoedemaekers, K.; Derksen, J.; Hoogstrate, S.W.; Wolters-Arts, M.; Oh, S.A.; Twell, D.; Mariani, C.; Rieu, I. BURSTING POLLEN is required to organize the pollen germination plaque and pollen tube tip in Arabidopsis thaliana. New Phytol. 2015, 206, 255–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tan, X.; Wang, M.; Li, B.; Zhao, Y.; Wu, C.; Rui, Q.; Wang, J.; Liu, Z.; Bao, Y. Exocyst subunit SEC3A marks the germination site and is essential for pollen germination in Arabidopsis thaliana. Sci. Rep. 2017, 7, 40279. [Google Scholar] [CrossRef] [PubMed]
- Hiscock, S.J.; Allen, A.M. Diverse cell signalling pathways regulate pollen-stigma interactions: The search for consensus. New Phytol. 2008, 179, 286–317. [Google Scholar] [CrossRef] [PubMed]
- Parre, E.; Geitmann, A. More than a leak sealant. The mechanical properties of callose in pollen tubes. Plant Physiol. 2005, 137, 274–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winship, L.J.; Obermeyer, G.; Geitmann, A.; Hepler, P.K. Under pressure, cell walls set the pace. Trends Plant Sci. 2010, 15, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Dehors, J.; Mareck, A.; Kiefer-Meyer, M.C.; Menu-Bouaouiche, L.; Lehner, A.; Mollet, J.C. Evolution of cell wall polymers in tip-growing land plant gametophytes: Composition, distribution, functional aspects and their remodeling. Front. Plant Sci. 2019, 10, 441. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Boavida, L.C.; Aggarwal, M.; Wu, H.M.; Feijó, J.A. The pollen tube journey in the pistil and imaging the in vivo process by two-photon microscopy. J. Exp. Bot. 2010, 61, 1907–1915. [Google Scholar] [CrossRef] [Green Version]
- Higashiyama, T.; Yang, W. Gametophytic Pollen Tube Guidance: Attractant Peptides, Gametic Controls, and Receptors. Plant Physiol. 2017, 173, 112–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, H.; Higashiyama, T. Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 2016, 531, 245–248. [Google Scholar] [CrossRef]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen Tube Growth and Guidance Is Regulated by POP2, an Arabidopsis Gene that Controls GABA Levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, P.B.; Liu, X.; Wu, X.; Zhu, S.; Kasahara, R.D. Fertilization in flowering plants: An odyssey of sperm cell delivery. Plant Mol. Biol. 2020, 103, 9–32. [Google Scholar] [CrossRef] [PubMed]
- Völz, R.; Heydlauff, J.; Ripper, D.; VonLyncker, L.; Groß-Hardt, R. Ethylene Signaling Is Required for Synergid Degeneration and the Establishment of a Pollen Tube Block. Dev. Cell 2013, 25, 310–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinkl, G.M.; Zwiebel, B.I.; Grier, D.G.; Preuss, D. Pollen-stigma adhesion in Arabidopsis: A species-specific interaction mediated by lipophilic molecules in the pollen exine. Development 1999, 126, 5431–5440. [Google Scholar] [PubMed]
- Hiscock, S.J.; Hoedemaekers, K.; Friedman, W.E.; Dickinson, H.G. The Stigma Surface and Pollen-Stigma Interactions in Senecio squalidus L. (Asteraceae) following Cross (Compatible) and Self (Incompatible) Pollinations. Int. J. Plant Sci. 2002, 163. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; Bhatla, S.C. Structural analysis of stigma development in relation with pollen-stigma interaction in sunflower. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 420–429. [Google Scholar] [CrossRef]
- Boyle, T.H.; Stimart, D.P. Self-incompatibility and interspecific incompatibility: Relationships in intra- and interspecific crosses of Zinnia elegans Jacq. and Z. angustifolia HBK (Compositae). Theor. Appl. Genet. 1986, 73, 305–315. [Google Scholar] [CrossRef]
- Yuan, B.; Ding, G.; Ma, J.; Wang, L.; Yu, L.; Ruan, X.; Zhang, X.; Zhang, W.; Wang, X.; Xie, Q. Comparison of Morphological Characteristics and Determination of Different Patterns for Rubber Particles in Dandelion and Different Rubber Grass Varieties. Plants 2020, 9, 1561. [Google Scholar] [CrossRef] [PubMed]
- Schuchovski, C.; Meulia, T.; Sant’Anna-Santos, B.F.; Fresnedo-Ramírez, J. Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz. Plants 2020, 9, 1258. [Google Scholar] [CrossRef]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in brassica and arabidopsis stigmas intersect at exo70a1, a putative component of the exocyst complex. Plant Cell 2009, 21, 2655–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscock, S.J. Self-incompatibility in Senecio squalidus L. (Asteraceae). Ann. Bot. 2000, 85, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; Thorogood, C.J.; Hegarty, M.J.; Lexer, C.; Hiscock, S.J. Pollen–pistil interactions and self-incompatibility in the Asteraceae: New insights from studies of Senecio squalidus (Oxford ragwort). Ann. Bot. 2011, 108, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, S.; Kanaoka, M.M.; Hashimoto, K.; Takakura, K.-I.; Nishida, T. Pollen-pistil interactions in reproductive interference: Comparisons of heterospecific pollen tube growth from alien species between two native Taraxacum species. Funct. Ecol. 2014, 28, 450–457. [Google Scholar] [CrossRef]
- Shivanna, K.R.; Tandon, R. Pollen–Pistil Interaction. In Reproductive Ecology of Flowerin Plants: A Manual; Shivanna, K.R., Tandon, R., Eds.; Springer: New Delhi, India, 2014; pp. 97–105. [Google Scholar]
- Chen, M.; Fan, W.; Hao, B.; Zhang, W.; Yan, M.; Zhao, Y.; Liang, Y.; Liu, G.; Lu, Y.; Zhang, G.; et al. EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, D.; Vekemans, X.; Castric, V.; Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New Phytol. 2005, 168, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Barberini, M.L.; Sigaut, L.; Huang, W.; Mangano, S.; Juarez, S.P.D.; Marzol, E.; Estevez, J.; Obertello, M.; Pietrasanta, L.; Tang, W.; et al. Calcium dynamics in tomato pollen tubes using the Yellow Cameleon 3.6 sensor. Plant Reprod. 2018, 31, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, K.; Yin, G.; Liu, X.; Liu, M.; Cao, N.; Duan, Y.; Gao, H.; Wang, W.; Ge, W.; et al. Pollen-Expressed Leucine-Rich Repeat Extensins Are Essential for Pollen Germination and Growth. Plant Physiol. 2018, 176, 1993–2006. [Google Scholar] [CrossRef] [Green Version]
- Sede, A.R.; Borassi, C.; Wengier, D.L.; Mecchia, M.A.; Estevez, J.M.; Muschietti, J.P. Arabidopsis pollen extensins LRX are required for cell wall integrity during pollen tube growth. FEBS Lett. 2018, 592, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Borassi, C.; Sede, A.R.; Mecchia, M.A.; Salgado Salter, J.D.; Marzol, E.; Muschietti, J.P.; Estevez, J.M. An update on cell surface proteins containing extensin-motifs. J. Exp. Bot. 2016, 67, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.A.; Jones, J.D.G. The Role of Leucine-Rich Repeat Proteins in Plant Defences. Adv. Bot. Res. 1997, 24, 89–167. [Google Scholar]
- Hiscock, S.J.; Kües, U.; Dickinson, H.G. Molecular mechanisms of self-incompatibility in flowering plants and fungi. Different means to the same end. Trends Cell Biol. 1996, 6, 421–428. [Google Scholar] [CrossRef]
- Sanabria, N.; Goring, D.; Nürnberger, T.; Dubery, I. Self/nonself perception and recognition mechanisms in plants: A comparison of self-incompatibility and innate immunity. New Phytol. 2008, 178, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Gavazzi, F.; Pigna, G.; Braglia, L.; Gianì, S.; Breviario, D.; Morello, L. Evolutionary characterization and transcript profiling of β-tubulin genes in flax (Linum usitatissimum L.) during plant development. BMC Plant Biol. 2017, 17, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kost, B.; Mathur, J.; Chua, N. Cytoskeleton in plant development. Curr. Opin. Plant Biol. 1999, 2, 462–470. [Google Scholar] [CrossRef]
- Cyr, R.J.; Palevitz, B.A. Organization of cortical microtubules in plant cells. Curr. Opin. Cell Biol. 1995, 7, 65–71. [Google Scholar] [CrossRef]
- McCann, M.C.; Roberts, K. Changes in cell wall architecture during cell elongation. J. Exp. Bot. 1994, 45, 1683–1691. [Google Scholar] [CrossRef]
- Dardelle, F.; Lehner, A.; Ramdani, Y.; Bardor, M.; Lerouge, P.; Driouich, A.; Mollet, J.C. Biochemical and immunocytological characterizations of arabidopsis pollen tube cell wall. Plant Physiol. 2010, 153, 1563–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Kaida, R. Functions of xyloglucan in plant cells. Mol. Plant 2011, 4, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Maris, A.; Suslov, D.; Fry, S.C.; Verbelen, J.P.; Vissenberg, K. Enzymic characterization of two recombinant xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Arabidopsis and their effect on root growth and cell wall extension. J. Exp. Bot. 2009, 60, 3959–3972. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Gao, C.; Yue, Y.; Liu, Z.; Ma, C.; Zhou, G.; Yang, Y.; Duan, Z.; Li, B.; Wen, J.; et al. Time-Course Transcriptome Analysis of Compatible and Incompatible Pollen-Stigma Interactions in Brassica napus L. Front. Plant Sci. 2017, 8, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanares, C.; Barth, S.; Thorogood, D.; Byrne, S.L.; Yates, S.; Czaban, A.; Asp, T.; Yang, B.; Studer, B. A Gene Encoding a DUF247 Domain Protein Cosegregates with the S Self-Incompatibility Locus in Perennial Ryegrass. Mol. Biol. Evol. 2016, 33, 870–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pütter, K.M.; van Deenen, N.; Müller, B.; Fuchs, L.; Vorwerk, K.; Unland, K.; Bröker, J.N.; Scherer, E.; Huber, C.; Eisenreich, W.; et al. The enzymes OSC1 and CYP716A263 produce a high variety of triterpenoids in the latex of Taraxacum koksaghyz. Sci. Rep. 2019, 9, 5942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A.; et al. WEGO 2.0: A web tool for analyzing and plotting GO annotations, 2018 update. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selected DEGs | Source | BLASTX e-Value 1 | Percent Identity | Description | Abbreviation |
|---|---|---|---|---|---|
| utg21288.4 | Venn/Visual analysis | 0.0 | 82.67% | leucine-rich repeat extensin-like protein 4 [Lactuca sativa] | LRX4 |
| utg8190.13 | Venn/Visual analysis | 9 × 10−150 | 99.55% | tubulin beta chain-like [Lactuca sativa] | TUBB |
| utg9900.15 | Visual analysis | 0.0 | 91.77% | probable xyloglucan endotransglucosylase/hydrolase protein 33 [Lactuca sativa] | XTH33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wollenweber, T.E.; van Deenen, N.; Roelfs, K.-U.; Prüfer, D.; Gronover, C.S. Microscopic and Transcriptomic Analysis of Pollination Processes in Self-Incompatible Taraxacum koksaghyz. Plants 2021, 10, 555. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030555
Wollenweber TE, van Deenen N, Roelfs K-U, Prüfer D, Gronover CS. Microscopic and Transcriptomic Analysis of Pollination Processes in Self-Incompatible Taraxacum koksaghyz. Plants. 2021; 10(3):555. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030555
Chicago/Turabian StyleWollenweber, Tassilo Erik, Nicole van Deenen, Kai-Uwe Roelfs, Dirk Prüfer, and Christian Schulze Gronover. 2021. "Microscopic and Transcriptomic Analysis of Pollination Processes in Self-Incompatible Taraxacum koksaghyz" Plants 10, no. 3: 555. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030555