Bioherbicidal Potential of the Essential Oils from Mediterranean Lamiaceae for Weed Control in Organic Farming


1
Department of Agricultural and Environmental Science, Università degli Studi di Bari Aldo Moro, Via Amendola 165/a, 70125 Bari, Italy
2
CIHEAM Centre International des Hautes Études en Agriculture Méditerranéenne, IAM Mediterranean Agronomic Institute of Bari, Via Ceglie 9, 70010 Valenzano, Italy
*
Author to whom correspondence should be addressed.
Plants 2021, 10(4), 818; https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040818
Submission received: 10 March 2021
/
Revised: 7 April 2021
/
Accepted: 9 April 2021
/
Published: 20 April 2021
(This article belongs to the Special Issue Sustainable Weed Management)
Abstract
:In all farming systems, weeds are the most expensive pest to manage, accounting for 30% of potential losses. In organic farming, the problem may be further amplified by restrictions on herbicides, thus making weeds the main problem faced by organic farmers in the field. In this sense, much research is focusing on the allelopathic potential of plants as an ecological weed control tool. Many plant species can release allelopathic compounds with high phytotoxicity that can be used in weed control. Species belonging to the Lamiaceae family have been studied widely for this purpose, and their essential oils (EOs) appear to be promising bioherbicides. However, there are still many challenges for their development. Considering these aspects, a review of the bioherbicidal effect of EOs from Mediterranean Lamiaceae could help identify the most effective ones and the challenges for their actual development.
1. Introduction
The emerging worldwide need to find alternatives to synthetic herbicides for sustainable weed control has prompted considerable interest in exploiting the natural herbicidal potential in plants [1]. Bioherbicide sources are sought out by both conventional and organic farming systems: the former wish to identify new sites of action to cope with weed resistance, the latter seek potent alternatives to synthetic herbicides that can be integrated in an overall management approach [2]. In this context, weed control research has recently focused on extracts from allelopathic species. These are species that can release secondary metabolites able to interfere with the growth and functions of surrounding plants [3].
A well-established group of allelopathic plants is that of the Lamiaceae family. They are known to contain high concentrations of volatile allelochemicals, which are responsible for their aroma, and are reported to give the species a competitive advantage in their natural habitats [4]. In this context, extracts of different Lamiaceae species were studied extensively and found to inhibit the germination and growth of many weed species [5,6,7,8,9,10,11,12]. Essential oils (EOs) from species such as oregano, thyme, rosemary, sage and mint are reported to be particularly strong bioherbicide candidates.
The phytotoxic effect of these species extracts, notably EOs, has mainly been linked to the presence of volatile bio-active compounds such as α-pinene, limonene, 1,8-cineole, carvacrol, camphor and thymol, which have been shown to have varying individual phytotoxicity levels [4,11,12,13,14,15]. Some phenolic compounds present in the EOs were also reported to be involved in allelopathic interactions and were even used to develop commercial bioherbicides. The mechanisms by which these allelochemicals can affect weeds was not discussed in detail. Only a few individual compounds were studied [16,17,18], in addition to the mechanism behind some naturally occurring allelopathic interactions [16,19,20,21].
Although there are numerous studies reporting on the successful use of EOs in weed control, to date there are still many constraints limiting their practical application in commercial bioherbicides. For instance, the role of the EO composition is still not clearly described. The mechanisms of action and the observed selectivity are also very poorly understood, limiting their rational implementation. Moreover, studies concerning the possible side effects of these EOs on beneficial soil microorganisms are still lacking.
This review will address all the above-mentioned issues pertaining to the use of EOs from Lamiaceae species in weed control in order to further highlight their potential uses and perspectives for future studies. It will also review the literature on certain species most frequently studied.
2. Weed Management in Organic Agriculture
2.1. Objectives and Methods of Weed Management in Organic Agriculture
To understand the aims of weed control in organic farming one must understand the overall objective of this production system. How to maximize yields and economic gain are major concerns for organic farmers, like others. However, in this system the emphasis is on the long-term outcome and overall health of the soil, plants, animals, and humans rather than just immediate maximum profitability [22]. In this context, many operational techniques have been defined to meet what could be regarded as the main goals of organic farming. Kirschenmann [23] presented four techniques related to the different aspects of management: nutrients, insects, plant disease and weeds. As for the latter, the overall goal was “to achieve weed control using crop rotation systems to deprive weeds of favorable growing conditions”. Liebman et al. [24] defined more detailed, equally important objectives. These can be summarized as follows:
- reducing weed density to a tolerable level, instead of targeting 100% control or total suppression;
- reducing the damage that a given density of weeds can cause, by increasing the competitive ability of crops and minimizing that of weeds through different preventive and cultural tools (competitive varieties, fertilization, irrigation and false seed beds);
- shifting the composition of weed communities to less aggressive, more easily managed species.
These goals may be achieved through a knowledge-intensive process. A good understanding of weed ecology, of the site and of the crop-weed interactions is required.
Kirschenmann [23] claims in this context that the organic system seeks to farm like nature, which implies knowing and understanding the natural processes and incorporating those principles on the farm.
Managing weeds in an organic system is more complicated than in a conventional system, mainly because of restrictions on the use of herbicides [25]. The latter are easy to apply and aggressively marketed, although in recent years there has been a tendency to restrict the use of chemicals in agriculture to preserve human health and the environment. [24]. Therefore, a combination of tools and practices that take into account the natural system’s cycles and interactions are increasingly being adopted to manage weeds. The management system is consequently an integrated approach, one that adopts different preventive, cultural, and direct control methods to achieve the goals detailed in the paragraph above [25,26].
- Preventive methods
Prevention aims at reducing the density of the actual weed vegetation by exhausting the potential weed vegetation (e.g., weed seedbank in the soil). This means reducing in-crop weed emergence and weed seed dispersal. Operational techniques include crop rotation, tillage systems, the false seedbed technique, cover crops, mulching and soil solarization.
- Cultural methods
Cultural methods are commonly used to reduce the need for direct weed control (e.g., herbicides) and increase its effectiveness. This is achieved by choosing cultural techniques that favor the competitive ability of crops against weeds. Cultural weed management techniques include crop genotype choice, planting pattern, polyculture production systems, fertilization, and irrigation strategies.
- Direct methods
Direct methods aim at intervening directly during the crop cycle to eliminate the weeds, mainly using physical or chemical tools.
Physical tools include mechanical weeding or cultivation, which is based on a variety of equipment. Recent technical innovations focusing on intra-row weed control in arable and vegetable crops have proven to be effective [26]. Robotic control is another technological innovation increasingly adopted by organic farmers. Post-emergence flame weeding, which can be used after planting or crop emergence, is another physical technique. Flaming can be used to eliminate weeds within the row where cultivation is difficult or can considerably damage the crop. Lastly, manual weeding is also widely used in organic management, notably when other measures are not feasible, such as within rows or when the crop is susceptible to damage by cultivation.
Bioherbicides are the main chemical tools. The latter are compounds and secondary metabolites derived from microbes, phytotoxic plant extracts or single compounds [2].
2.2. Challenges of the Current Weed Management Methods in Organic Agriculture
Effective weed management in the organic system necessarily involves integration of the highest number of available tools and approaches [27,28]. Preventive methods based on ecological principles and building biodiversity such as rotation or cover cropping are of particular interest to this system. However, direct methods are still contributing the most to weed control in many organic farms [27,29,30]. Therefore, serious weed competition problems may arise when few direct control methods are available or applicable. Moreover, those currently available present serious limitations (Table 1).
By studying existing limitations in direct weed control methods, it is possible to define research needs and opportunities. This is particularly true of bioherbicides, especially considering the limitations and the numerous potential sources of active compounds in nature. This research field is increasingly important due to increasing consumer awareness and environmental problems related to synthetic herbicides (residues, weed resistance). Usually, the EOs have various modes of action, and therefore it is more complicated for weeds to develop easily resistance against them [10,31]. This aspect increased the attention to their bioherbicidal potential, widely investigated in the hope of finding effective, viable products that can meet registration requirements.
3. The Use of Plant-Based Bioherbicides
3.1. Bioherbicidal Potential in Plants
The interest in exploiting the natural herbicidal potential of plants stems from a worldwide need to find new sustainable weed control strategies [1].
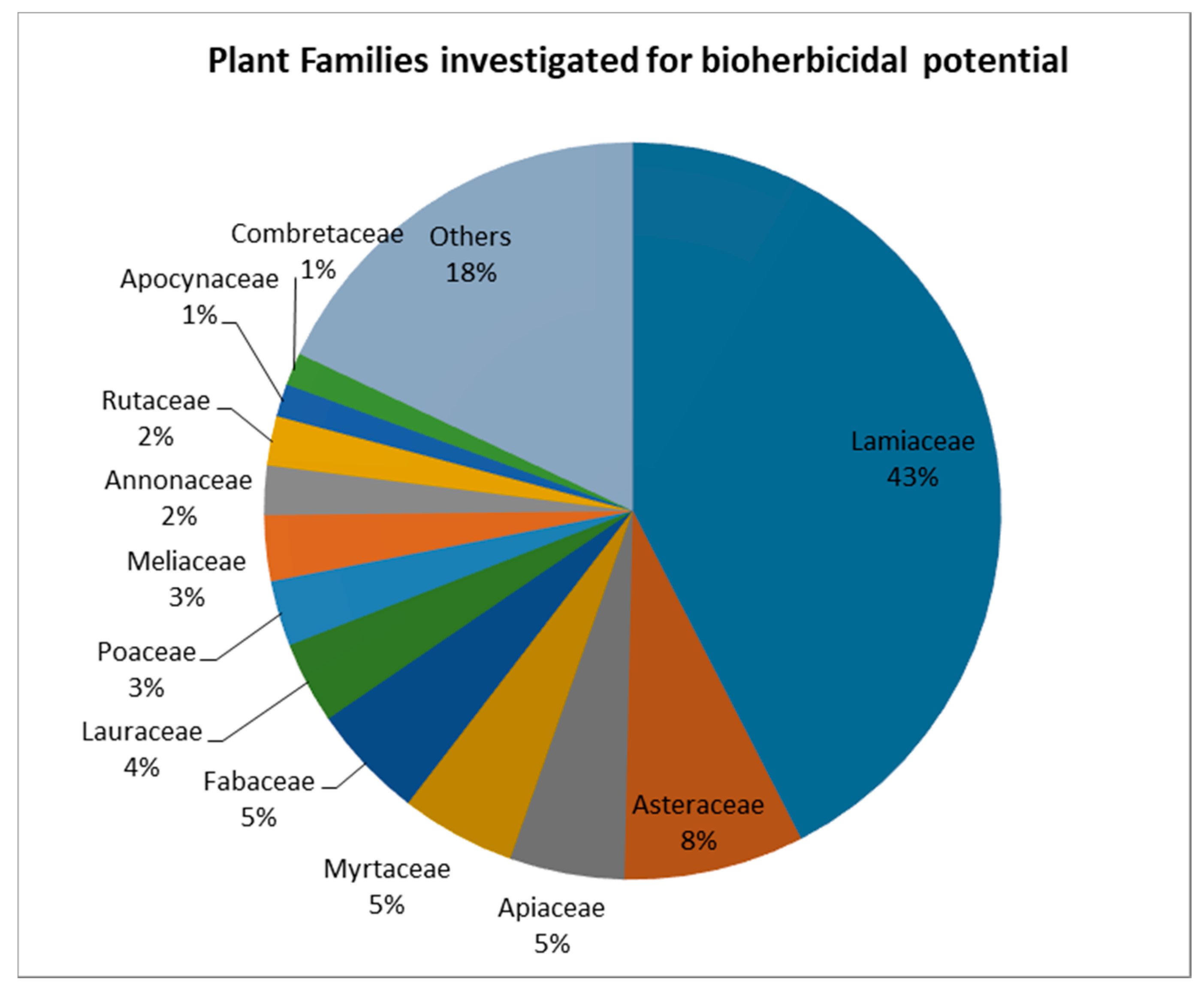
As plants are the richest source of active organic compounds on Earth, the bioherbicidal potential of a long list of plant species has been explored [32]. A Scopus literature search using the keywords « Bioherbicides AND plant extracts » and « Bioherbicides AND Essential oils » found 130 articles (excluding review articles) on the bioherbicidal potential of species from 38 different families (Figure 1).
Plant species can be considered for investigations due to their known composition in terms of biologically active compounds, or an observed allelopathic effect in their natural environment. Allelopathy is a characteristic of many plant species, and can be defined as a form of interaction between plants through chemical inhibitors released from living or decaying tissues [3]. Evidence that some plants are able to inhibit the growth of other plants in their surroundings has long been known and reported, and studies have linked these interactions to the presence of compounds named “allelochemicals” [4,21,33]. This can justify the high interest in the Lamiaceae family species—accounting for 43% of the total studied species from the papers covered by the search—which are known to possess high concentrations of volatile allelochemicals. In this context, much effort has been made to extract allelochemicals from plants and test their bioherbicidal activity in bioassays; many were successful (Table 2). Other studies evaluated the activity of single compounds isolated from plants, such as flavonoids, alkaloids and terpenoids. These different classes of secondary metabolites have different importance in term of bioherbicidal activity. In extracts from the same species, they can occur in different proportions, depending on the type of extract and the extraction method [34].
3.2. Types of Active Compounds and Plant Extracts Tested as Bioherbicides
3.2.1. Active Compounds with Bioherbicidal Potential
The term active compounds usually refers to secondary metabolites occurring in plants, known for having diverse biological activities. These are the compounds with no relevance to vital functions (like respiration, photosynthesis and reproduction), but involved in interactions between plants and their surrounding environment, notably as part of their mechanism of defense against stress [40,41]. Secondary metabolites in plants have been classified differently by different authors; a recent review by Yasri et al., [42] defined four main groups: terpenoids, phenolics, sulphur-containing secondary metabolites and nitrogen-containing secondary metabolites. Not all of these groups of secondary metabolites were found to be implicated in allelopathic interactions or showed a bioherbicidal potential. Although some authors included amino acids and proteins among the phytotoxic compounds, terpenoids and phenolics were the ones most frequently studied [4]. Only these two groups will therefore be considered in detail in this paragraph.
- (a)—Terpenoids
The terpenoid group is present in the majority of secondary metabolite classifications and is reported to be very important in allelopathic interactions [4,21,43]. The compounds of this group, sometimes referred to as volatile allelochemicals, can be divided into monoterpenes, sesquiterpenes, diterpene, triterpenes and polyterpenes [42]. Monoterpenes are the major constituents of essential oils and have been shown to inhibit seed germination and seedling growth [14]. They are the most frequently described secondary metabolites for bioherbicidal activity [4,14,42,43,44,45,46]. Some monoterpene-based commercial herbicides have been developed such as cinmethylin, which is a derivative of 1,4-cineole [14]. Compounds having different chemical functions (Table 3) belonging to this sub-group were found to have varied inhibition effects.
Most authors considered that ketone-containing compounds such as camphor and pulegone are the most toxic, followed by alcohol compounds such as cineol and citronellol, and by ether, diene and monoene compounds such as α-pinene, which are the least toxic [47]. This was confirmed by many other authors [14,47]. Considering that plant species and chemotypes have different monoterpene composition, the phytotoxicity of extracts can vary between plant materials with different percentages of effective compounds (e.g camphor). However, there is no clear evidence reported in the literature as to how the active compounds of a plant extract define its activity level. In other words, it is not clear whether the observed toxic effect of plant extracts is due to the potent phytotoxicity of a single compound or to the synergic action of many constituents.
- (b)—Phenolics
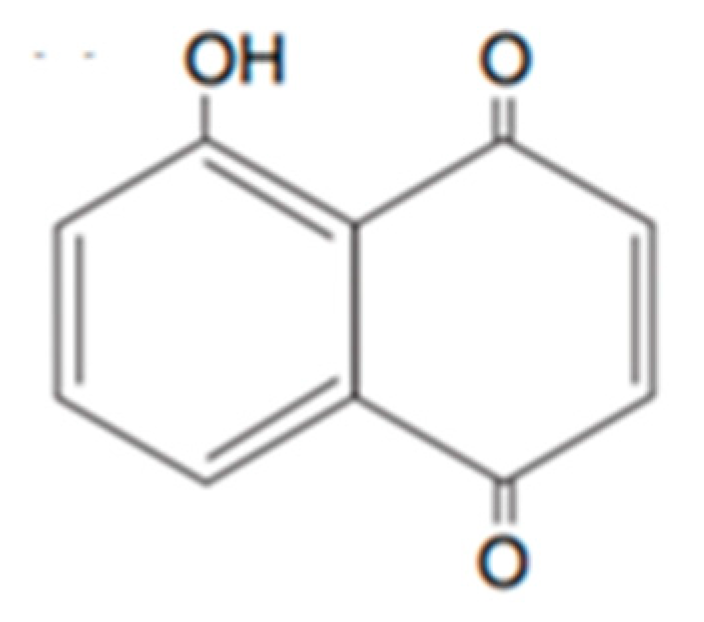
Plant phenolics include phenolic acids, flavonoids and tannins. They are synthetized by plants as a response to ecological and physiological conditions, mainly when they are under biotic or abiotic stress [54]. Like the terpenoids, an important focus exists on the identification of phenols with bioherbicidal activity. This was attributed to the easiness of their extraction and their water solubility [47]. They are usually the main components in aqueous and organic solvents extracts, and their polarity determines the type and amount of phenols extracted. An example of a well-studied phenolic for this effect is juglone (Figure 2) produced by walnuts [55].
3.2.2. Types of Plant Extracts Tested for their Bioherbicidal Activity
The extraction method is a determining factor in the recovery of active compounds from plants, considering that secondary metabolites of different groups have varying chemical properties (volatility, polarity etc.). For instance, anthocyanins, tannins, saponins and terpenoids can be recovered using water, whereas polyphenols, flavonoids, flavones and alkaloids require organic solvents [34]. Like the terpenoids, much research focuses on identifying phenols with bioherbicidal potential. This is because they are easily extracted and are soluble in water [43]. They are usually the main components in aqueous and organic solvent extracts, the polarity of which determines the type and amount of phenols extracted. An example of a well-studied phenolic with this effect is juglone (Figure 2), produced by walnuts [55]. These different extracts often show different levels of toxicity. In a study conducted to test Calamintha nepeta L. (Savi) as a source of phytotoxic compounds, solvents of varying polarity (n-hexane, chloroform, ethyl acetate and n-butanol) were used to fractionate the leaves’ methanol extract. The study defined the following hierarchical phytotoxicity: ethyl acetate > n-hexane > chloroform > n-butanol [56].
In general, three main groups of extracts can be found in the literature: essential oils (EOs), aqueous extract and organic solvent extracts.
- Essential oils: Sometimes called volatile oils, these are natural substances that can be extracted from aromatic plants by distillation or by appropriate mechanical process without heating. EOs mainly contain compounds that can be volatilized during extraction, making this an effective means of extracting plant terpenoids in the purest form [18,57]. These, the most frequently tested extracts from aromatic plants, can cause higher phytoxicity compared to aqueous or organic solvent extracts [38,56,58].
- Aqueous extracts: These are obtained simply by soaking in water ground dry material from plants, from which water-soluble compounds are extracted. Several phenols are water soluble and can successfully be extracted using this method. Aqueous extracts have been used to investigate the bioherbicidal potential of many plants and have been found to produce significant effects mainly at the highest tested concentrations [1,59,60].
- Organic solvent extracts. This group consists mainly of phenols. As the type of solvent (mainly differing in polarity) affects the amount and type of phenols extracted, authors have used various solvents. Methanol, ethanol, acetone, ethyl acetate, n-hexane and chloroform are among those used most frequently [54]. The choice of solvent depends on the types of phenol present in the tested plant, and many authors have tested different ones simultaneously in order to compare the composition and phytotoxicity of the resulting extract [56,61].
3.3. Modes of Action of Plant Allelochemicals
After investigating the type of active compounds in the plants’ extracts, research has also explored the mechanisms of toxicity to weeds. The most frequently described effects are from single allelochemicals rather than whole plant extracts; the modes of action of terpenoids and phenolic acids, which are reported to be the most relevant secondary metabolites in allelopathic interactions, have been studied by many authors. However, studies on this topic are still lacking and the mechanism of only a few phytotoxic compounds has been described. This paragraph therefore focuses mainly on toxicity mechanisms reported for single allelochemicals, as well as allelopathic mechanisms observed in nature. Note that to determine the stage in which plants are most sensitive to allelochemicals, the latter were often tested in two different periods (pre-germination and post-emergence), and different features and mechanisms were analyzed accordingly.
3.3.1. Effect on Cells Division, Elongation and Structure
The size and weight of weed seedlings are the features most often measured to assess their reaction to the application of allelopathic compounds. The application of plant extracts usually results in a significant decrease of these parameters compared to the control. The substances undoubtedly affect the responsible physiological processes: cell division and elongation [21]. In this sense some studies have reported that some allelochemicals affect mitosis: the process was either slowed down [19], interrupted in the anaphases or hindered altogether [16,19,20,62,63]. All the cited studies measured the number of cells and their ultrastructure at specific times as indicators. Muller [16] also reported that volatile terpenes extracted from Salvia leucophylla Greene (mainly cineol and camphor), prevented the elongation of root and hypocotyl cells. Cineole is in fact the most widely described of all monoterpenes [64]. It is generally reported to strongly inhibit all stages of mitosis. The suggested mechanism can therefore result in considerable damage to weeds by reducing their growth or retarding it, which can give the crop a competitive advantage.
3.3.2. Effect on the Cells Membrane Integrity and Permeability
Cell membrane integrity is critical for cell functions and survival. Any alteration may compromise its role as a barrier, affecting permeability to nutrients or toxins or inducing the leakage of solutes [65,66]. A number of allelochemicals seem to alter plant cell membranes. Due to lipophilic nature of the cell membranes, monoterpenes can cause their destruction by increasing permeability or inhibiting enzymes [18]. Moreover, some monoterpenes are reported to induce oxidative stress; α-pinene, for example, caused lipid peroxidation when applied to young seedlings of Cassia occidentalis L., resulting in an increase in solute leakage [48]. Furthermore, some compounds produced changes to the permeability of membranes; Varona et al. [67] found that linalool caused an increase in permeability, whereas Muller et al. [16] found that permeability decreased after applying cineole and dipentene from S. leucophylla. This suggests that allelochemicals can result in important damage to weeds by acting at the membrane level.
3.3.3. Effect on Photosynthesis
There is evidence of a relationship between the visible effects on weeds and photosynthetic functions. Early studies found a correlation between the reduction in growth caused by a plant-extracted phenolic substance, “scopoletin”, and net photosynthesis in Amarantus retroflexus L. [68]. Many other studies found that a number of phenolic acids affect photosynthesis, and this was linked to changes to stomatal conductance or to plant chlorophyll contents [68,69]. Furthermore, many monoterpenes were also found to inhibit photosynthesis and chlorophyll synthesis [70]. Citronellol and 1,8-cineole, for example, showed a similar effect on the invasive weed species Ageratum conyzoides L.: its chlorophyll content decreased by 60% and 66%, respectively [18,50]. Eugenol, another monoterpene, has a similar effect: it induced photosynthetic inhibition by reducing chlorophyll content in C. occidentalis and Bidens pilosa L. [71]. These examples suggest that photosynthesis-related processes could be behind the observed damage. However, only a few of the allelochemicals were tested, and the actual cause-effect between the described processes is not yet well understood.
3.3.4. Effect on Nutrients Availability and Uptake
Because of the observed effects on the root appearance, some research has focused on whether allelochemicals inhibit nutrient uptake [21]. The uptake of phosphorous, potassium, calcium and zinc, for example, was affected either by the direct application of some phenolic acids or by growing plants in association with allelopathic species [72,73,74,75,76,77]. Moreover, some early studies found that toxic excretions from plants reduce the availability of nutrients by affecting nutrient cycling mechanisms; mineralization, for example, was suppressed by the root excretion of some natural forest vegetation due to its toxicity to the nitrification process [78]. This suggests that phytotoxic compounds from plants may affect soil microbial activity, which plays an essential role in making important nutrients like nitrogen available to plants.
All the presented modes of action suggest that allelochemicals have a strong potential as weed control tools. However, they also highlight the many challenges to their practical application. For instance, no clear selectivity can be concluded from the reported mechanisms, which means that crops may also be susceptible. Moreover, the impact on crop and soil health is also of concern if the allelochemicals have a detrimental effect on beneficial soil microbes. This, in addition to other possible challenges, will be detailed in the next paragraph.
3.4. Challenges and Perspectives to the Use of Plant Extracts as Bioherbicides
3.4.1. Challenges to the Use of Plant Extracts as Bioherbicides
- (a)—Unclear selectivity
Although allelochemicals may affect specific functions like photosynthesis or respiration, they lack site specificity, which excludes their use as selective bioherbicides. This also means they could be phytotoxic to crops and must be managed carefully when applied. However, many studies that tested plant extracts on different weed species revealed varying degrees of sensitivity. In most cases monocots were more resistant than dicots [7,8,79]. Moreover, it was frequently reported that many crops were less affected than weeds; for instance, when applying the EO of Satureja hortensis L. and Laurus nobilis L. at low concentrations, A. retroflexus germination decreased significantly whereas tomatoes were unaffected. However, at the highest tested concentration, tomato germination also decreased, albeit at a lower rate than A. retroflexus [8]. Similar results were obtained when applying Origanum onites L. and Rosmarinus officinalis L. on Avena sterilis L., Sinapis arvensis L. and a number of wheat cultivars, where the latter were less affected [7]. This suggests that careful dosage may resolve phytotoxicity to crops. Nevertheless, studies were not able to explain this variation in sensitivity, which makes it difficult to predict and exploit. Further research is required to better understand the mechanism of action of different allelochemicals and the synergies by which they operate in plant extracts.
- (b)—Toxicity to soil microorganisms
Organic farming relies on soil health and natural soil processes to satisfy crop nutrient needs and ensure long term fertility. In fact, one of the serious drawbacks of synthetic chemicals is their impact on soil biodiversity and their harmful effect on beneficial organisms. Plant extracts with similar effects cannot be recommended regardless of their possible effectiveness on weeds. Only a few studies have addressed this important aspect. As mentioned in the “effect on nutrients uptake and availability” paragraph, some allelochemicals may be detrimental to nitrification bacteria [78]. Moreover, many plant extracts, notably those from the Lamiaceae family, have been shown to possess antimicrobial properties [80,81,82]. Doubts may thus arise about their possible harmful effects on soil microbes. However, other studies have reported a positive impact in this respect; volatile substances from alfalfa (Medicago sativa L.), for example, induced a rapid increase in microbial respiration and fungi mycelium growth when added to the soil. Results thus suggest a possible beneficial effect on the initial colonization stage of plant residue decomposers [83]. The different findings may be ascribed to variations in the concentration of compounds in contact with microorganisms.
In summary, the soil microbial community seems to be affected by allelochemicals (either negatively or positively). Hence, when assessing the use of plant extracts as agrochemicals, care should also be taken to detect any possible negative repercussions on soil life, a crucial component of any sustainable management strategy.
- (c)—Degradation of plant extracts in the environment
While the incorporation of allelopathic plant species biomass into the soils is constrained by the difficulty in accumulating active concentrations [21], the direct use of concentrated extracts is mainly limited by susceptibility to environmental elements. Once released in the environment, the extracts are subject to decomposition either by microorganisms or by chemical reactions. Blum [84] reported in the book chapter «Fate of phenolic allelochemicals in soils − the role of soil and rhizosphere microorganisms» that because microorganisms use phenolic acids as a source of carbon or energy, they are thus more subject to microbial transformation and utilization than to other processes (ionization, oxidation, sorption onto soil particles, fixation into the recalcitrant organic matter (e.g., polymerization)). In this chapter the author reports results from many studies suggesting that this degradation is very likely and that phenolic acids are unlikely to produce any phytotoxic effects. Moreover, Marmulla and Harder [85] report that monoterpenes such as d-limonene, α-pinene, γ-terpinene and terpinolene are readily biodegradable. They also found that different monoterpenes show different susceptibility to degradation. In addition, many allelochemicals are highly susceptible to spontaneous decomposition; abiotic photochemical processes in the atmosphere can result in lifetimes of minutes to hours, as cited by the same authors [21]. However, very little is known about their abiotic degradation in soil [81]. These aspects suggest that allelochemicals may lack the necessary persistence to be effective bioherbicides. This may be remedied by selecting critical stages of weed growth. Even a brief period of phytotoxicity could affect the competitive ability of weeds with respect to crops [81]. Another approach recently under study is the use of innovative formulations that could regulate the rate of release without compromising the desired concentration levels. For instance, experiments with rosemary EO encapsulated in a starch matrix were successful [86].
3.4.2. Perspectives for the Use of Plant Extracts as Bioherbicides
Despite the many constraints, the use of plant extracts for weed control is still considered a field with great potential. However, to address limitations, research should focus on better understanding the phenomena in terms of:
- Linking the observed effects of extracts to the action of specific compounds and their synergies;
- Defining the mechanisms behind the phytotoxicity to enhance it and understand the selectivity;
- Defining the most sensitive stages of weed development to increase effectiveness and tackle the problem of the limited duration of the effect;
- Defining innovative formulations that take into consideration the interactions of the extracts with field conditions (soil texture, microorganisms and abiotic factors such as light and temperature);
- Defining innovative techniques for the cultivation and extraction of essential oils to guarantee the commercial feasibility of a mass production large quantity of EOs;
- Defining formulations that allow for containing the concentrations of EOs within technical limits for an easy application on an agricultural scale.
4. Examples of Lamiaceae Species with Bioherbicidal Potential
4.1. Oregano
In the literature, oregano is used to refer to a number of species in different genera and families, the leaves and flowers of which have a common characteristic odor and flavor [87,88]. The major oregano species belonging to the Lamiaceae family, Poliomintha longiflora L., Origanum vulgare L. and O. onites, are mainly found in the Mediterranean basin [87,88]. O. vulgare, and O. vulgare subsp. hirtum (Link) Iestwart plants in particular are extremely rich in essential oils (up to 8% dry weight) [87]. The famous odor and flavor of these species are mainly linked to their carvacrol content, which in addition to thymol, p-cymene and γ-terpinene, is known to be the major component of oregano essential oil [31,89,90]. As for the bioherbicidal effect, pure carvacrol and the EO from some species (mainly O. vulgare and O. onites) have been widely investigated, with interesting findings. In a study by De Mastro et al. [31] on A. retroflexus and Lolium perenne L., carvacrol at the concentration of 0.3 µL/mL completely inhibited the germination of both species. The same study assessed the application of dry biomass of an oregano hybrid (O. vulgare ssp. virilidum x O. vulgare L. ssp. hirtum) in a pot trial, and promising results were obtained using 20 g per kg of soil. In another investigation by Atak et al. [7], O. onites EO was tested against A. sterilis and S. arvensis L., and severe inhibition was observed on both species starting from 0.2 µL/mL. In this experiment the EO was also tested on a number of wheat cultivars, which were found to be less sensitive: this led the authors to suggest a possible dosed application of the EO as a bioherbicide in wheat fields. Ibáñez and Blázquez [91] also tested an EO dominated by carvacrol (60.42%) against Portulaca oleracea L., Lolium multiflorum L. and Echinochloa crus-galli L. They found that germination was completely inhibited in all the species starting from the lowest tested dose: 0.125 µL/mL. Hanana et al. [92] found the same high effectiveness at low doses on some important weed species (S. arvensis L., Phalaris paradoxa L. and Lolium rigidum Gaud) using a carvacrol- and δ-terpinene- rich EO from O. vulgare. Its high yield and strong anti-germination and phytotoxic effect make oregano EO a promising bioherbicide candidate.
4.2. Rosemary
Rosemary (R. officinalis), an evergreen shrub that can grow up to 2 m high, has aromatic leaves and flowers rich in essential oils [87,93]. Native to the Mediterranean region, it is characterized by high tolerance to heat, drought, and poor, dry, sandy and rocky soil types [87]. It is grown in different parts of the world, such as Europe, Africa and Asia, and is used in various culinary, pharmaceutical and cosmetics industries [94]. There are three species in the Rosmarinus genus, R. officinalis, R. eryocalix and R. tomentosus, but R. officinalis is the most widely distributed and important for its valuable EO, which can be extracted in amounts ranging from 0.9 to 2.5%, depending on many factors [93,95,96]. Rosemary EO is appreciated in cosmetics for its strong camphorous aroma, and in medicine for its content in biological compounds of high value [87,97].
The compounds generally found to be dominant in R. officinalis EO are 1,8-cineol, camphor, α-pinene, borneol, p-cymene and verbenone, as reported in Hernández et al. [96]. The proportions of these, however, vary considerably among chemotypes [98,99]. R. officinalis is one of the Lamiaceae species that has received considerable attention in plant-based bioherbicide research. Many recent studies have investigated the phytotoxicity of its extracts to weeds, and in this respect significant results were obtained on a number of important weed species, such as A. retroflexus L., Bromus tectorum L., Cynodon dactylon L., Digitaria sanguinalis L. and L. perenne [5,8,99]. All the studies reported a concentration-dependent effect: some even found that at very low concentrations, such as 100 or 200 μL/L, the extracts had a stimulatory effect rather than a phytotoxic one [8]. Nevertheless, for some species, significant decreases in germination were found at concentrations as low as 400 μL/L [8,99]. More resistant species (mostly monocots), however, were only sensitive to higher tested doses [5,7,8,99]. A range of concentrations were therefore always tested. As EOs are very susceptible to environmental conditions, recent studies are now investigating innovative formulations to support their practical implementation as bioherbicides. For instance, nanoformulation and encapsulation in starch were successfully tested as germination and early growth inhibitors [100].
4.3. Thyme
The genus Thymus L. from the Lamiaceae family consists of over 200 species of herbaceous perennials and small shrubs [101,102]. Many of these species are widespread in the world, but the center of the genus is considered to be in the Mediterranean region [101,103]. Thymus is one of the most studied genera for bioactive activity due to its wide use in folk medicine [102]. This bioactive activity can be linked to its high phenolic monoterpene content (e.g., carvacrol and thymol are the major compounds in the species EO), in addition to α-terpinene, α-cymene and borneol [6,92,101]. One of the emerging bioactivity investigations of Thymus spp. EO is in weed control research. Similar to oregano, low doses of EO from thyme were reported to have potent phytotoxic effects on a number of problematic weed species. For instance, Hanana et al. [92] tested the EO from Thymus capitatus against S. arvensis, P. paradoxa and L. rigidum, and germination inhibition was significant at concentrations as low as 0.25 µL/mL. A recent study by Sarić-Krsmanović et al. [104] also tested the bioherbicidal effect of T. vulgaris EO, which is dominated by carvacrol (17.0%), thymol (11.6%) and p-cymene (11.6%), against Abutilon theophrasti Medik. Complete germination inhibition was obtained at a concentration of 1% EO. After applying Thymus spp. EO, other weed species such as A. retroflexus, Avena fatua L., Datura stramonium L., Lepidium sativum L. and Agrostemma githago L. were also significantly affected, either at germination or at the seedling growth stage [6,105]. These findings indicate that different species of Thymus (T. fallax Fisch. et Mey, T. vulgaris L., T. capitatus (L.) Hoffmanns. & Link, T. daenensis Celak.), even if somewhat varied in chemical composition, have great potential as bioherbicides. Kashkooli and Saharkhiz [6] tested different ecotypes of the same species (T. daenensis Celak.), and despite the important chemical variations in the EOs, no significant differences were observed in their effect on weeds.
4.4. Mint
Mentha L. is another important genus of the Lamiaceae family that is widely distributed and cultivated in most parts of the world thanks to its adaptation to diverse environments. About 42 species and 15 hybrids fall under this genus: they are commonly characterized by odorous secondary metabolites that make its EO famous [106,107]. In fact, most Mentha species are industrially cultivated for EO production. Members of the genus Mentha show a great variability in chemical composition, both intra- and inter-species, resulting in different chemotypes. Nevertheless, most of the species are either C3-oxygenated p-menthane types (e.g., pulegone, menthone, menthol) or C6-oxygenated p-menthane (e.g., carvone) types [107,108]. The EOs of species from both types figure in bioherbicide research. For instance, the EOs of M. dumetorum Schult, Mentha × piperita L. cv. Mitcham (Peppermint), M. pulegium L. and M. spicata L. were successfully tested on different weed species as germination and growth inhibitors. Different studies tested them on a variety of weed species with varying results. Onaran et al. [105] found that M. dumetorum EO suppressed the germination of A. theophrasti better than other tested EOs such as O. vulgare and T. fallax. Another test used the EO from a specific Peppermint cultivar (Mentha × piperita L. cv. Mitcham) with 35% menthol on tomato (Lycopersicon esculentum Mill.) and radish (Raphanus sativus L.), in addition to three weeds: Convolvulus arvensis L., P. oleracea and Echinochloa colonum L. The EO caused varying degrees of inhibition, depending on the species; tomato was particularly sensitive, and germination was completely suppressed at 900µL/L, whereas field bindweed and purslane were still able to germinate even at 1500µL/L [109]. Similarly, a study by Argyropoulos et al. [110] assessed the effects of a Mentha species EO (M. spicata) on two horticultural crops (cotton and tomato), besides weeds (A. retroflexus, E. crus-galli, Oryza sativa L., P. oleracea and Setaria verticillata). The EO containing 82% trans piperitone oxide severely inhibited all the tested species, but with a greater effect on cotton. The EOs of Mentha, like that of the other species discussed above, mainly have dose- and species-dependent effects. The reason for this selectivity and the mechanisms involved are still unclear.
4.5. Other Species
With fewer occurrences in the literature than those detailed above, other species in the Lamiaceae family were used to produce bioherbicidal extracts, mainly EOs. Among these number the Salvia, Satureja, Nepeta and Lavandula species [8,79,111,112]. Findings reveal differences among tested species and in resulting toxicity levels, but all conclude that the tested monoterpene-rich EOs are promising.
5. Conclusions
In summary, many Lamiaceae species are valuable for the bioactive compounds they contain. They are widely distributed throughout the Mediterranean area and are currently cultivated in most parts of the world. Many yield significant amounts of EOs (up to 8%) rich in terpenes, which are considered important in allelopathic interactions, and may thus have potential as bioherbicides. To this end, a considerable amount of research is still needed on technical aspects, such as exploring the mechanism of action, understanding selectivity, investigating side effects on beneficial plants, and exploring innovative formulations for effective application. Furthermore, studies are required to assess cost-benefits and define the target value for the class crops, as well as the environmental impact of production.
Author Contributions
Conceptualization, G.D.M.; methodology, J.E.M.; investigation, J.E.M.; writing—original draft preparation, J.E.M.; review and editing, J.E.M., C.R., G.D.M.; project administration, G.D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the bioherbicide potential of Eucalyptus globulus Labill: Biochemistry and effects of its aqueous extract. PLoS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [Green Version]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.-P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop. Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Allelopathy. In Fundamentals of Weed Science, 5th ed.; Zimdahl, R.L., Ed.; Academic Press: Fort Collins, CO, USA, 2018; pp. 253–270. [Google Scholar]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of Essential Oils as Bioherbicides. J. Essent. Oil Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Chen, F.; Peng, S.; Chen, B.-M.; Ni, G.; Liao, H. Allelopathic potential and volatile compounds of Rosmarinus officinalis L. against weeds. Allelopath. J. 2013, 32, 57–66. [Google Scholar]
- Kashkooli, A.B.; Saharkhiz, M.J. Essential Oil Compositions and Natural Herbicide Activity of Four Denaei Thyme (Thymus daenensis Celak.) Ecotypes. J. Essent. Oil Bear. Plants 2014, 17, 859–874. [Google Scholar] [CrossRef]
- Atak, M.; Mavi, K.; Uremis, I. Bioherbicidal effects of oregano and rosemary essential oils on germination and seedling growth of bread wheat cultivars and weeds. Rom. Biotechnol. Lett. 2016, 21, 11149–11158. [Google Scholar]
- Hazrati, H.; Saharkhiz, M.J.; Moein, M.; Khoshghalb, H. Phytotoxic effects of several essential oils on two weed species and Tomato. Biocatal. Agric. Biotechnol. 2018, 13, 204–212. [Google Scholar] [CrossRef]
- Matković, A.; Marković, T.; Vrbničanin, S.; Sarić-Krsmanović, M.; Božić, D. Chemical composition and in vitro herbicidal activity of five essential oils on Johnson grass (Sorghum halepense [L.] Pers.). Lek. Sirovine 2018, 38, 44–50. [Google Scholar] [CrossRef]
- Jouini, A.; Verdeguer, M.; Pinton, S.; Araniti, F.; Palazzolo, E.; Badalucco, L.; Laudicina, V. Potential Effects of Essential Oils Extracted from Mediterranean Aromatic Plants on Target Weeds and Soil Microorganisms. Plants 2020, 9, 1289. [Google Scholar] [CrossRef]
- Abd-Elgawad, A.M.; El Gendy, A.E.-N.G.; Assaeed, A.M.; Al-Rowaily, S.L.; Alharthi, A.S.; Mohamed, T.A.; Nassar, M.I.; Dewir, Y.H.; ElShamy, A.I. Phytotoxic Effects of Plant Essential Oils: A Systematic Review and Structure-Activity Relationship Based on Chemometric Analyses. Plants 2020, 10, 36. [Google Scholar] [CrossRef]
- Maccioni, A.; Santo, A.; Falconieri, D.; Piras, A.; Farris, E.; Maxia, A.; Bacchetta, G. Phytotoxic effects of Salvia rosmarinus essential oil on Acacia saligna seedling growth. Flora-Morphol. Distrib. Funct. Ecol. Plants 2020, 269, 151639. [Google Scholar] [CrossRef]
- Muller, W.H.; Lorber, P.; Haley, B.; Johnson, K. Volatile Growth Inhibitors Produced by Salvia leucophylla: Effect on Oxygen Uptake by Mitochondrial Suspensions. Bull. Torrey Bot. Club 1969, 96, 89–96. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Spencer, G.F. Volatile Monoterpenes as Potential Parent Structures for New Herbicides. Weed Sci. 1993, 41, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Alipour, M.; Saharkhiz, M.J. Phytotoxic activity and variation in essential oil content and composition of Rosemary (Rosmarinus officinalis L.) during different phenological growth stages. Biocatal. Agric. Biotechnol. 2016, 7, 271–278. [Google Scholar] [CrossRef]
- Muller, W.H. Volatile Materials Produced by Salvia leucophylla: Effects on Seedling Growth and Soil Bacteria. Int. J. Plant Sci. 1965, 126, 195–200. [Google Scholar] [CrossRef]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Varela, R.M.; Simonet, A.M.; Carrera, C.; Molinillo, J.M. Allelopathy as a new strategy for sustainable ecosystems development. Biol. Sci. Space 2003, 17, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Grana, E.; Sanchez-Moreiras, A.M.; Reigosa, M.J. Mode of Action of Monoterpenes in Plant-Plant Interactions. Curr. Bioact. Compd. 2012, 8, 80–89. [Google Scholar] [CrossRef]
- Cornman, I. The responses of onion and lily mitosis to coumarin and parasorbic acid. J. Exp. Biol. 1947, 23, 292–297. [Google Scholar] [PubMed]
- Jensen, T.E.; Welbourne, F. The Cytological Effects of Growth Inhibitors on Excised Roots of Vicia Faba and Pisum Sativum. 1962. Available online: https://sdaos.org/wp-content/uploads/pdfs/Vol%2041%201962/62p131.pdf (accessed on 22 July 2019).
- Rice, E.L. Allelopathy, 2nd ed.; Rice, E.L., Ed.; Academic Press: San Diego, CA, USA, 1984; pp. 8–344. [Google Scholar]
- Drinkwater, L. Ecological knowledge: Foundation for sustainable organic agriculture. In Organic Farming: The Ecological System; Francis, C., Ed.; American society of agronomy: New York, NY, USA, 2009; pp. 19–47. [Google Scholar]
- Kirschenmann, F.L. Cultivating an Ecological Conscience: Essays from a Farmer Philosopher; Falk, C., Ed.; University Press of Kentucky: Lexington, KY, USA, 2010; pp. 48–61. [Google Scholar] [CrossRef]
- Liebman, M.; Mohler, C.L.; Staver, C.P. Weed management: A need for ecological. In Ecological Management of Agricultural Weeds; Liebman, M., Mohler, C.L., Staver, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 1–20. [Google Scholar]
- Bastiaans, L.; Paolini, R.; Baumann, D.T. Focus on ecological weed management: What is hindering adoption? Weed Res. 2008, 48, 481–491. [Google Scholar] [CrossRef]
- Barberi, P. Weed management in organic agriculture: Are we addressing the right issues? Weed Res. 2002, 42, 177–193. [Google Scholar] [CrossRef]
- Leslie, A.; Chen, G.; Hooks, C.R. Organic Integrated Weed Management in Vegetables. 2016. Available online: https://extension.umd.edu/sites/extension.umd.edu/files/_images/programs/mdvegetables/OrganicWeedMgntVeg21May2016(1).pdf (accessed on 25 July 2019).
- Bàrberi, P. Integrated weed management in organic cropping systems. In Improving Organic Crop Cultivation; Köpke, U., Ed.; Burleigh Dodds: Bonn, Germany, 2018; pp. 323–341. [Google Scholar]
- Smith, R.; Lanini, W.T.; Gaskell, M.; Mitchell, J.; Koike, S.T.; Fouche, C. Weed Management for Organic Crops. 2000, Volume 7250, p. 2. Available online: https://anrcatalog.ucanr.edu/pdf/7250.pdf (accessed on 3 September 2019).
- Bond, W.; Grundy, A.C. Non-chemical weed management in organic farming systems. Weed Res. 2001, 41, 383–405. [Google Scholar] [CrossRef]
- Da Mastro, G.; Fracchiolla, M.; Verdini, L.; Montemurro, P. Oregano and its Potential Use as Bioherbicide. Acta Hortic. 2006, 723, 335–346. [Google Scholar] [CrossRef]
- Flamini, G. Natural Herbicides as a Safer and More Environmentally Friendly Approach to Weed Control: A Review of the Literature Since 2000. In Studies in Natural Products Chemistry; Elsevier BV: Amsterdam, The Netherlands, 2012; Volume 37, pp. 353–396. [Google Scholar]
- Molisch, H. Der Einfluss einer Pflanze auf die Andere, Allelopathie. Nat. Cell Biol. 1938, 141, 493. [Google Scholar] [CrossRef]
- Azmir, J.; Zaidul, I.; Rahman, M.; Sharif, K.; Mohamed, A.; Sahena, F.; Jahurul, M.; Ghafoor, K.; Norulaini, N.; Omar, A. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- El-Gawad, A.A.; ElShamy, A.; El Gendy, A.E.-N.; Gaara, A.; Assaeed, A. Volatiles Profiling, Allelopathic Activity, and Antioxidant Potentiality of Xanthium Strumarium Leaves Essential Oil from Egypt: Evidence from Chemometrics Analysis. Molecules 2019, 24, 584. [Google Scholar] [CrossRef] [Green Version]
- Benchaa, S.; Hazzit, M.; Zermane, N.; Abdelkrim, H. Chemical composition and herbicidal activity of essential oils from two Labiatae species from Algeria. J. Essent. Oil Res. 2019, 31, 335–346. [Google Scholar] [CrossRef]
- Pardo-Muras, M.; Puig, C.G.; López-Nogueira, A.; Cavaleiro, C.; Pedrol, N. On the bioherbicide potential of Ulex europaeus and Cytisus scoparius: Profiles of volatile organic compounds and their phytotoxic effects. PLoS ONE 2018, 13, e0205997. [Google Scholar] [CrossRef]
- Kayanifard, M.; Mohsenzadeh, S. Allelopathic Analysis of Four Ecotypes of Ajowan. Iran. J. Sci. Technol. Trans. A Sci. 2017, 41, 971–978. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Setia, N.; Kohli, R.K. Herbicidal activity of volatile oils from Eucalyptus citriodora against Parthenium hysterophorus. Ann. Appl. Biol. 2005, 146, 89–94. [Google Scholar] [CrossRef]
- Bernhoft, A. A Brief Review on Bioactive Compounds in Plants. In Bioactive Compounds in Plants—Benefits and Risks for Man and Animals; Bernhoft, A., Ed.; The Norwegian Academy of Science and Letters: Oslo, Norway, 2010; pp. 11–17. [Google Scholar]
- Yasri, A.; Naboulsi, I.; Aboulmouhajir, A.; Kouisni, L.; Bekkaoui, F. Plants extracts and secondary metabolites, their extraction methods and use in agriculture for controlling crop stresses and improving productivity: A review. Acad. J. Med. Plants 2018, 6, 223–240. [Google Scholar]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef]
- Echeng, F.; Echeng, Z. Research Progress on the use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Asplund, R. Monoterpenes: Relationship between structure and inhibition of germination. Phytochemistry 1968, 7, 1995–1997. [Google Scholar] [CrossRef]
- Reynolds, T. Comparative Effects of Alicyclic Compounds and Quinones on Inhibition of Lettuce Fruit Germination. Ann. Bot. 1987, 60, 215–223. [Google Scholar] [CrossRef]
- Dudai, N.; Ben-Ami, M.; Chaimovich, R.; Chaimovitsh, D. Essential oils as allelopathic agents: Bioconversion of monoterpenes by germinating wheat seeds. Acta Hortic. 2004, 629, 505–508. [Google Scholar] [CrossRef]
- Elakovich, S.D. Terpenoids as Models for New Agrochemicals. In Biologically Active Natural Products; Cutler, H.G., Ed.; ACS Symposium Series; American Chemical Society: Hattiesburg, MS, USA, 1988; Volume 380, pp. 250–261. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Arora, K.; Kohli, R.K. α-Pinene Inhibits Growth and Induces Oxidative Stress in Roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Vaid, S.; Batish, D.R.; Singh, H.P.; Kohli, R.K. Phytotoxicity of limonene against Amaranthus viridis L. Bioscan 2011, 6, 163–165. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic effect of two volatile monoterpenes against bill goat weed (Ageratum conyzoides L.). Crop. Prot. 2002, 21, 347–350. [Google Scholar] [CrossRef]
- Singh, D.H.; Batish, D.R.; Kaur, S.; Vaid, S.; Kohli, R. Weed suppressing ability of some monoterpenes. J. Plant Dis. Prot. 2004, 19, 821–828. [Google Scholar]
- Vokou, D.; Douvli, P.; Blionis, G.J.; Halley, J.M. Effects of monoterpenoids, acting alone or in pairs, on seed germination and subsequent seedling growth. J. Chem. Ecol. 2003, 29, 2281–2301. [Google Scholar] [CrossRef] [PubMed]
- De Martino, L.; Mancini, E.; De Almeida, L.F.R.; De Feo, V. The Antigerminative Activity of Twenty-Seven Monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Stewart, A.J.; Stewart, R.F. Phenols. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 2682–2689. [Google Scholar]
- Araniti, F.; Lupini, A.; Mercati, F.; Statti, G.A.; Abenavoli, M.R. Calamintha nepeta L. (Savi) as source of phytotoxic compounds: Bio-guided fractionation in identifying biological active molecules. Acta Physiol. Plant 2013, 35, 1979–1988. [Google Scholar] [CrossRef]
- Eslahi, H.; Fahimi, N.; Sardarian, A. Chemical composition of essential oils: Chemistry, safety and applications. In Essential Oils in Food Processing: Chemistry, Safety and Applications; Hashemi, S.M.B., Khaneghah, A.M., Sant’Ana, A.S., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 119–171. [Google Scholar]
- Kordali, S.; Tazegul, A.; Cakir, A. Phytotoxic effects of Nepeta meyeri Benth. Extracts and essential oil on seed germinations and seedling growths of four weed species. Rec. Nat. Prod. 2015, 9, 404–418. [Google Scholar]
- Islam, A.M.; Hasan, M.; Musha, M.H.; Uddin, K.; Juraimi, A.S.; Anwar, P. Exploring 55 tropical medicinal plant species available in Bangladesh for their possible allelopathic potentiality. Ann. Agric. Sci. 2018, 63, 99–107. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Pandino, G.; Onofri, A.; Mauromicale, G. Allelopathic effects of Cynara cardunculus L. leaf aqueous extracts on seed germination of some Mediterranean weed species. Ital. J. Agron. 2018, 11, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Tigre, R.; Silva, N.; Santos, M.; Honda, N.; Falcão, E.; Pereira, E. Allelopathic and bioherbicidal potential of Cladonia verticillaris on the germination and growth of Lactuca sativa. Ecotoxicol. Environ. Saf. 2012, 84, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Issa, M.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Yadav, S.S.; Kumari, A. Appraisal of phytotoxic, cytotoxic and genotoxic potential of essential oil of a medicinal plant Vitex negundo. Ind. Crops Prod. 2020, 145, 112083. [Google Scholar] [CrossRef]
- Dutra, Q.P.; Christ, J.A.; Carrijo, T.T.; Alves, T.D.A.; Alves, T.D.A.; Mendes, L.A.; Praça-Fontes, M.M. Phytocytotoxicity of volatile constituents of essential oils from Sparattanthelium Mart. species (Hernandiaceae). Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Oliva, A. Mode of Action of Phytotoxic Terpenoids. In Allelopathy: Chemistry and Mode of Action of Allelochemicals; Macias, F.A., Galindo, J.C.G., Molinillo, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 201–216. ISBN 0-8493-1964-1. [Google Scholar]
- McNeil, P.L.; Steinhardt, R.A. Loss, Restoration, and Maintenance of Plasma Membrane Integrity. J. Cell Biol. 1997, 137, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieracci, J.P.; Armando, J.W.; Westoby, M.; Thommes, J. Industry review of cell separation and product harvesting methods. In Biopharmaceutical Processing; Jagschies, G., Lindskog, E., Łącki, K., Galliher, P., Eds.; Elsevier: Cambridge, CA, USA, 2018; pp. 165–206. [Google Scholar]
- Varona, S.; Martín, Á.; Cocero, M.J. Formulation of a natural biocide based on lavandin essential oil by emulsification using modified starches. Chem. Eng. Process. Process. Intensif. 2009, 48, 1121–1128. [Google Scholar] [CrossRef]
- Einhellig, F.A.; Rice, E.L.; Risser, P.G.; Wender, S.H. Effects of Scopoletin on Growth, CO2 Exchange Rates, and Concentration of Scopoletin, Scopolin, and Chlorogenic Acids in Tobacco, Sunflower, and Pigweed. Bull. Torrey Bot. Club 1970, 97, 22–23. [Google Scholar] [CrossRef] [Green Version]
- Einhellig, F.A.; Rasmussen, J.A. Effects of three phenolic acids on chlorophyll content and growth of soybean and grain sorghum seedlings. J. Chem. Ecol. 1979, 5, 815–824. [Google Scholar] [CrossRef]
- Pouresmaeil, M.; Nojadeh, M.S.; Movafeghi, A.; Maggi, F. Exploring the bio-control efficacy of Artemisia fragrans essential oil on the perennial weed Convolvulus arvensis: Inhibitory effects on the photosynthetic machinery and induction of oxidative stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar] [CrossRef]
- Vaid, S.; Batish, D.; Singh, D.H.; Kohli, R. Phytotoxic effect of eugenol towards two weedy species. Bioscan 2010, 5, 339–341. [Google Scholar]
- Chambers, E.E.; Holm, L.G. Phosphorus Uptake as Influenced by Associated Plants. Weeds 1965, 13, 312–314. [Google Scholar] [CrossRef]
- Iii, C.E.O.; Rice, E.L. Relative Effects of Known Plant Inhibitors on Species from First Two Stages of Old-Field Succession. Southwest. Nat. 1970, 15, 165. [Google Scholar] [CrossRef]
- Glass, A.D.M. Influence of Phenolic Acids on Ion Uptake: I. Inhibition of phosphate uptake. Plant Physiol. 1973, 51, 1037–1041. [Google Scholar] [CrossRef]
- Glass, A.D.M. Influence of Phenolic Acids upon Ion Uptake: III. Inhibition of potassium absorption. J. Exp. Bot. 1974, 25, 1104–1113. [Google Scholar] [CrossRef]
- Harper, J.R.; Balke, N.E. Characterization of the Inhibition of K+ Absorption in Oat Roots by Salicylic Acid. Plant Physiol. 1981, 68, 1349–1353. [Google Scholar] [CrossRef] [Green Version]
- Glass, A.D.M. The allelopathic potential of phenolic acids associated with the rhizosphere of Pteridium aquilinum. Can. J. Bot. 1976, 54, 2440–2444. [Google Scholar] [CrossRef]
- Greenland, D.J. Nitrate fluctuations in tropical soils. J. Agric. Sci. 1958, 50, 82–92. [Google Scholar] [CrossRef]
- Cavalieri, A.; Caporali, F. Effects of essential oils of cinnamon, lavender and peppermint on germination of Mediterranean weeds. Allelopath. J. 2010, 25, 1–5. [Google Scholar]
- Al-Mariri, A.; Safi, M. In Vitro Antibacterial Activity of Several Plant Extracts and Oils against Some Gram-Negative Bacteria. Iran. J. Med Sci. 2014, 39, 36–43. [Google Scholar]
- Elisha, I.L.; Botha, F.S.; McGaw, L.J.; Eloff, J.N. The antibacterial activity of extracts of nine plant species with good activity against Escherichia coli against five other bacteria and cytotoxicity of extracts. BMC Complement. Altern. Med. 2017, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, A.A.; Al-Askar, A.A.; Almaary, K.S.; Dawoud, T.M.; Sholkamy, E.N.; Bakri, M.M. Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi, J. Biol. Sci. 2018, 25, 361–366. [Google Scholar] [CrossRef]
- Menzies, J.D.; Gilbert, R.G. Responses of the Soil Microflora to Volatile Components in Plant Residues. Soil Sci. Soc. Am. J. 1967, 31, 495–496. [Google Scholar] [CrossRef]
- Blum, U. Fate of phenolic allelochemicals in soils—The role of soil and rhizosphere microorganisms. In Allelopathy: Chemistry and Mode of Action of Allelochemicals; Macías, F.A., Galindo, J.C.G., Molinillo, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 57–76. [Google Scholar]
- Emarmulla, R.; Eharder, J. Microbial monoterpene transformations—A review. Front. Microbiol. 2014, 5, 346. [Google Scholar] [CrossRef] [Green Version]
- Alipour, M.; Saharkhiz, M.J.; Niakousari, M.; Damyeh, M.S. Phytotoxicity of encapsulated essential oil of rosemary on germination and morphophysiological features of amaranth and radish seedlings. Sci. Hortic. 2019, 243, 131–139. [Google Scholar] [CrossRef]
- Kokkini, S.; Karousou, R.; Hanlidou, E. HERBS|Herbs of the Labiatae. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 3082–3090. [Google Scholar]
- Rodriguez-Garcia, I.; Silva-Espinoza, B.; Ortega-Ramirez, L.; Leyva, J.; Siddiqui, M.; Cruz-Valenzuela, M.; Gonzalez-Aguilar, G.; Ayala-Zavala, J.F. Oregano Essential Oil as an Antimicrobial and Antioxidant Additive in Food Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 1717–1727. [Google Scholar] [CrossRef]
- Figiel, A.; Szumny, A.; Gutiérrez-Ortíz, A.; Carbonell-Barrachina, Á.A. Composition of oregano essential oil (Origanum vulgare) as affected by drying method. J. Food Eng. 2010, 98, 240–247. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare ) extracts and essential oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Herbicidal value of essential oils from oregano-like flavour species. Food Agric. Immunol. 2017, 28, 1168–1180. [Google Scholar] [CrossRef] [Green Version]
- Hanana, M.; Mansour, M.B.; Algabr, M.; Amri, I.; Gargouri, S.; Romane, A.; Jamoussi, B.; Hamrouni, L. Potential use of es-sential oils from four Tunisian species of Lamiaceae: Biological alternative for fungal and weed control. Rec. Nat. Prod. 2017, 11, 258–269. [Google Scholar]
- Sasikumar, B. Rosemary. In Handbook of Herbs and Spices; Peter, K.V., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 243–255. [Google Scholar]
- Ribeiro-Santos, R.; Carvalho-Costa, D.; Cavaleiro, C.; Costa, H.S.; Albuquerque, T.G.; Castilho, M.C.; Ramos, F.; Melo, N.R.; Sanches-Silva, A. A novel insight on an ancient aromatic plant: The rosemary (Rosmarinus officinalis L.). Trends Food Sci. Technol. 2015, 45, 355–368. [Google Scholar] [CrossRef]
- Ghanmi, M.; Satrani, B.; Aberchane, M.; Ismaili, M.R.; Aafi, A.; Abid, A.E. Nature valorisation et caractérisation des PAM des strates arbustives et herbacées. In Plantes Aromatiques et Médicinales au Maroc: Les Milles et une Vertus; Bayed, G.E., Ed.; Centre de Recherche Forestière: Rabat, Morocco, 2011; p. 28. [Google Scholar]
- Hernández, M.D.; Sotomayor, J.A.; Hernandez, A.S.M.; Jordán, M.J. Rosemary (Rosmarinus officinalis L.) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 677–688. [Google Scholar]
- Andrade, J.M.; Faustino, C.; Garcia, C.; Ladeiras, D.; Reis, C.P.; Rijo, P. Rosmarinus officinalis L.: An update review of its phytochemistry and biological activity. Futur. Sci. OA 2018, 4, FSO283. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Cervelli, C.; Ruffoni, B.; Shachter, A.; Dudai, N. Volatile diversity in wild populations of rosemary ( Rosmarinus officinali s L.) from the Tyrrhenian Sea vicinity cultivated under homogeneous environmental conditions. Ind. Crops Prod. 2016, 84, 381–390. [Google Scholar] [CrossRef]
- El Mahdi, J.; Tarraf, W.; Ruta, C.; Piscitelli, L.; Aly, A.; De Mastro, G. Bio-Herbicidal Potential of The Essential Oils from Different Rosmarinus officinalis L. Chemotypes in Laboratory Assays. Agronomy 2020, 10, 775. [Google Scholar] [CrossRef]
- de Barros Fernandes, R.V.; Borges, S.V.; Botrel, D.A. Gum arabic/starch/maltodextrin/inulin as wall materials on the micro-encapsulation of rosemary essential oil. Carbohydr. Polym. 2014, 101, 524–532. [Google Scholar] [CrossRef]
- Ghasemi Pirbalouti, A.; Bistghani, Z.; Malekpoor, F. An overview on genus Thymus. J. Herb. Drugs. 2015, 6, 93–100. [Google Scholar]
- Leal, F.; Taghouti, M.; Nunes, F.; Silva, A.C.C.A.; Matos, M. Thymus Plants: A Review—Micropropagation, Molecular and Antifungal Activity. In Active Ingredients from Aromatic and Medicinal Plants; El-Shemy, H., Ed.; IntechOpen: London, UK, 2017; pp. 107–126. [Google Scholar]
- Morales, R. The history, botany and taxonomy of the genus Thymus. In Thyme, the Genus Thymus; Stahl-Biskup, E., Sáez, F., Eds.; Taylor & Francis: London, UK, 2002; pp. 1–43. ISBN 0415284880. [Google Scholar]
- Sarić-Krsmanović, M.; Gajić Umiljendić, J.; Radivojević, L.; Šantrić, L.; Potočnik, I.; Đurović-Pejčev, R. Bioherbicidal effects of five essential oils on germination and early seedling growth of velvetleaf (Abutilon theophrasti Medik.). J. Environ. Sci. Health B 2019, 54, 247–251. [Google Scholar] [CrossRef]
- Onaran, A.; Yilar, M.; Belguzar, S.; Bayan, Y.; Aksit, H. Antifungal and Bioherbicidal Properties of Essential Oils of Thymus fallax Fish & Mey., Origanum vulgare L. and Mentha dumetorum Schult. Asian J. Chem. 2014, 26, 5159–5164. [Google Scholar] [CrossRef]
- Fatih, B.; Madani, K.; Chibane, M.; Duez, P.; Brahmi, F.; Khodir, M.; Mohamed, C.; Pierre, D. Chemical Composition and Biological Activities of Mentha Species. In Aromatic and Medicinal Plants—Back to Nature; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2017; Chapter 3. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Armstrong, L.; Rescigno, A.; Yeskaliyeva, B.; Seitimova, G.; Beyatli, A.; Sharmeen, J.; Mahomoodally, M.F.; Sharopov, F.; Durazzo, A.; et al. Lamium Plants-A Comprehensive Review on Health Benefits and Biological Activities. Molecules 2019, 24, 1913. [Google Scholar] [CrossRef] [Green Version]
- Croteau, R. Metabolism of Monoterpenes in Mint (Mentha) Species. Planta Med. 1991, 57, S10–S14. [Google Scholar] [CrossRef] [PubMed]
- Mahdavikia, F.; Saharkhiz, M.J. Phytotoxic activity of essential oil and water extract of peppermint (Mentha×piperita L. CV. Mitcham). J. Appl. Res. Med. Aromat. Plants 2015, 2, 146–153. [Google Scholar] [CrossRef]
- Argyropoulos, E.; Eleftherohorinos, I.; Vokou, D. In vitro evaluation of essential oils from Mediterranean aromatic plants of the Lamiaceae for weed control in tomato and cotton crops. Allelopath. J. 2008, 22, 69–78. [Google Scholar]
- Mutlu, S.; Atici, O.; Esim, N. Bioherbicidal effects of essential oils of Nepeta meyeri Benth. on weed spp. Allelopath. J. 2010, 26, 291–300. [Google Scholar]
- Taban, A.; Saharkhiz, M.J.; Hadian, J. Allelopathic potential of essential oils from fourSaturejaspp. Biol. Agric. Hortic. 2013, 29, 244–257. [Google Scholar] [CrossRef]
Figure 1.
Plant families studied for their bioherbicidal potential. (Source: elaborated from a search on Scopus, 2019).
Figure 1.
Plant families studied for their bioherbicidal potential. (Source: elaborated from a search on Scopus, 2019).

Figure 2.
Representative structure of juglone.


{kind=link}
{kind=link}
Table 1.
Limitations to current direct weed control methods in organic farming.
| Direct Method | Limitations | Reference |
|---|---|---|
| Mechanical weeding (Tillage/ Cultivation) | Weather and soil moisture conditions Excessive soil disturbance Difficult to control perennial weeds Damages to crops root system No/reduced tillage systems Difficult to control within rows Stimulation of latent weed seed germination Most energy-consuming task (Fossil fuel) | [27,28,30] |
| Flame weeding | Possible damage to the crop Effectiveness depends on weed tolerance to heat and weather conditions High machine cost initially | [27,30] |
| Manual weeding | Large surfaces High cost | [27] |
| Bioherbicides | Limited products available (only 13 registered products for organic farming; only one is based on plant extract) Nonselective products Too expensive considering the necessary rates Marginal efficacy | [2,27,32] |
Table 2.
Examples of frequently tested families for bioherbicidal activity.
| Species | Family | Bio Herbicidal Effect | Reference |
|---|---|---|---|
| Xanthium strumarium L. | Asteraceae | Significant inhibition of germination and growth of the noxious weed Bidens pilosa L. | [35] |
| Thymus fontanesii Boiss. et Reut. Satureja calamintha subsp. nepeta Briq. | Lamiaceae | Wide herbicidal effect on seed germination and 3–4 leaf stage of Sinapis arvensis L, Avena fatua L., Sonchus oleraceus L., Xanthium strumarium L., Cyperus rotundus L. | [36] |
| Ulex europaeus L. Cytisus scoparius L. | Fabaceae | Pure volatile organic compounds extracted caused irreversible phytotoxicity for Digitaria sanguinalis L. | [37] |
| Trachyspermum copticum L. | Apiaceae | Germination and shoot/root length of Zea mays L. and Lepidium sativum L. significantly reduced by all concentrations of EO and methanol extract. | [38] |
| Eucalyptus citriodora Hook | Myrtaceae | Parthenium hysterophorus L.: Germination completely inhibited. Chlorophyll content and respiratory activity decreased for 4-week-old plants. | [39] |
Table 3.
Examples of monoterpenes and their phytotoxic effect.
| Monoterpene | Representative Structure | Chemical Function | Containing Plant Species | Germination Inhibition | Reference |
|---|---|---|---|---|---|
| α-pinene |  | Monoene | Eucalyptus grandis W. Hill ex Maiden Rosmarinus officinalis L. | Amaranthus hybridus L. * Portulaca oleracea L. * Pisum sativum L. Cicer arietinum L. | [47] [48] |
| Limonene |  | Monoene | Citrus limon (L.) Burm. f. Apium graveolens L. | Amaranthus viridis L. * | [49] |
| 1,8-cineole |  | Ether | Eucalyptus spp. R. officinalis L. | Ageratum conyzoides L. * | [50] |
| α-phellandrene |  | Diene | Ligusticum marginatum C.B. Clarke | Raphanus sativus L. * | [44] |
| Linalool |  | Alcohol | Mentha spp. Lavandula hybrida L. | Echinochloa crus-galli L. ** at highest concentration | [51] |
| Camphor |  | Ketone | Lavandula abrialis L. R. officinalis L. | Amaranthus retroflexus L. ** L. multiflorum L.** at low concentation | [14] |
| Pulegone |  | Ketone | Mentha piperita L. Calamintha arkansana (Nutt.) Shinners | R. sativus L. ** at low concentation | [44] |
| Menthol |  | Alcohol | Helianthus annuus L. Mentha spp. | A. retroflexus L. ** Lolium multiflorum L. ** Lactuca sativa L.* | [14] [49] |
| Citronellol |  | Alcohol | Rosa spp. Eucalyptus spp. | Chenopodium album L. * A. retroflexus L. * E. crus-galli L. ** at highest concentration | [18] [52] |
| Borneol |  | Alcohol | Salvia officinalis L. R. officinalis L. | Lepidium sativum L. R. sativus L. * | [53] |
| Carvacrol |  | Alcohol | Origanum vulgare L. Thymus capitatus L. | L. perenne L. ** A. retroflexus L. ** | [31] |
* Significant effect; ** Total inhibition; N: no significant effect.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
De Mastro, G.; El Mahdi, J.; Ruta, C. Bioherbicidal Potential of the Essential Oils from Mediterranean Lamiaceae for Weed Control in Organic Farming. Plants 2021, 10, 818. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040818
AMA Style
De Mastro G, El Mahdi J, Ruta C. Bioherbicidal Potential of the Essential Oils from Mediterranean Lamiaceae for Weed Control in Organic Farming. Plants. 2021; 10(4):818. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040818
Chicago/Turabian StyleDe Mastro, Giuseppe, Jihane El Mahdi, and Claudia Ruta. 2021. "Bioherbicidal Potential of the Essential Oils from Mediterranean Lamiaceae for Weed Control in Organic Farming" Plants 10, no. 4: 818. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040818
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.