
The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill)

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Effect of Biostimulant on Seed Germination
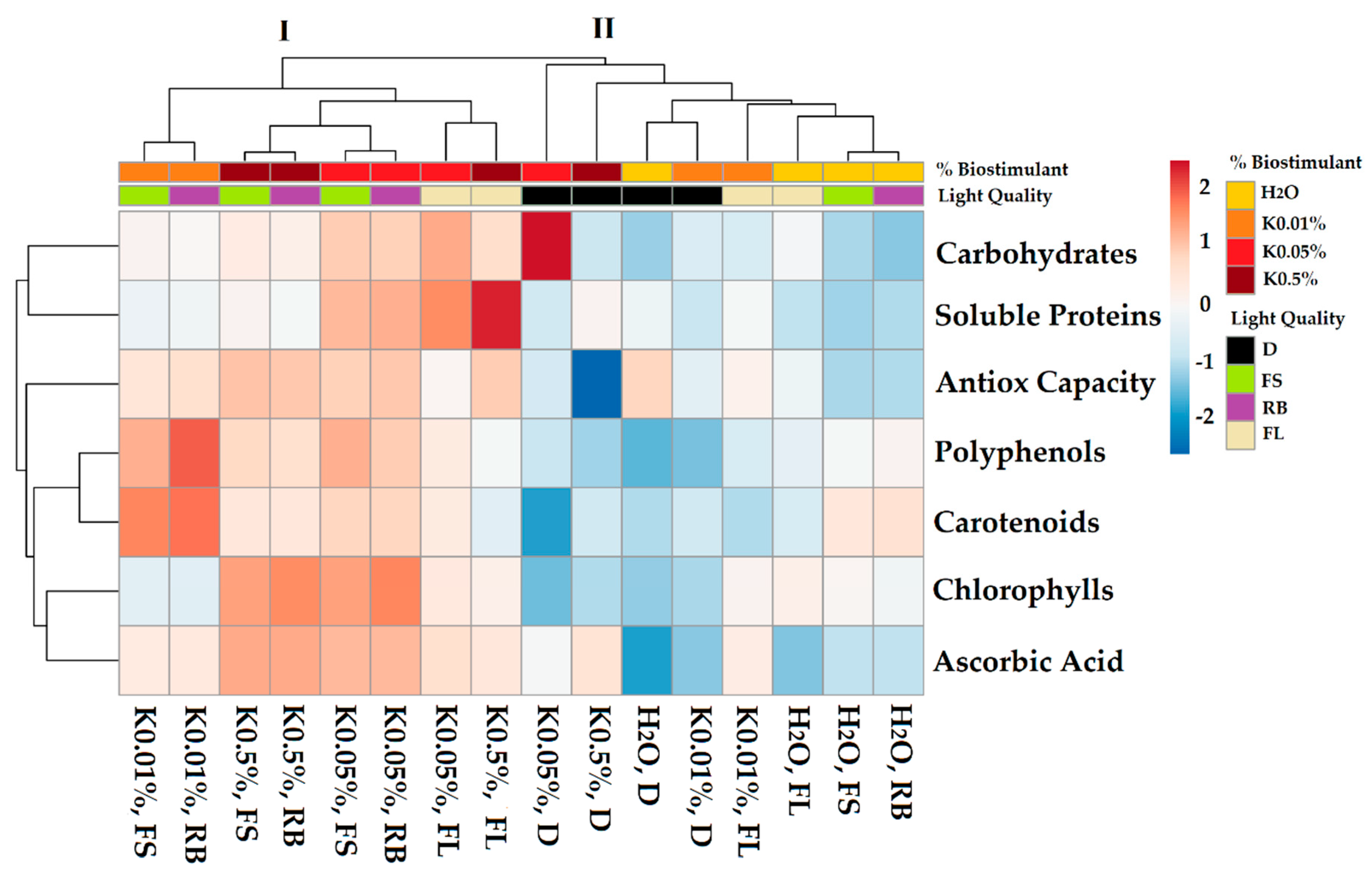
2.2. Effect of Biostimulant and Light Regimes on Sprout Bioactive Compounds, Proteins, and Sugars
2.3. Influence of Biostimulant and Light Quality on Seedling Morpho-Anatomical and Physiological Parameters
2.3.1. Morphological Traits and Leaf Anatomy
2.3.2. Pigments, Nitrogen Balance Index, and PSII Photochemistry
3. Discussion
3.1. Effects of Biostimulant Seed Pre-Treatment and Light Quality on Sprout Bioactive Compounds
3.2. Effects of Biostimulant Seed Pre-Treatment and Light Quality on Photosynthesis and Early Plant Development
4. Materials and Methods
4.1. Seed Pre-Treatment and Germination
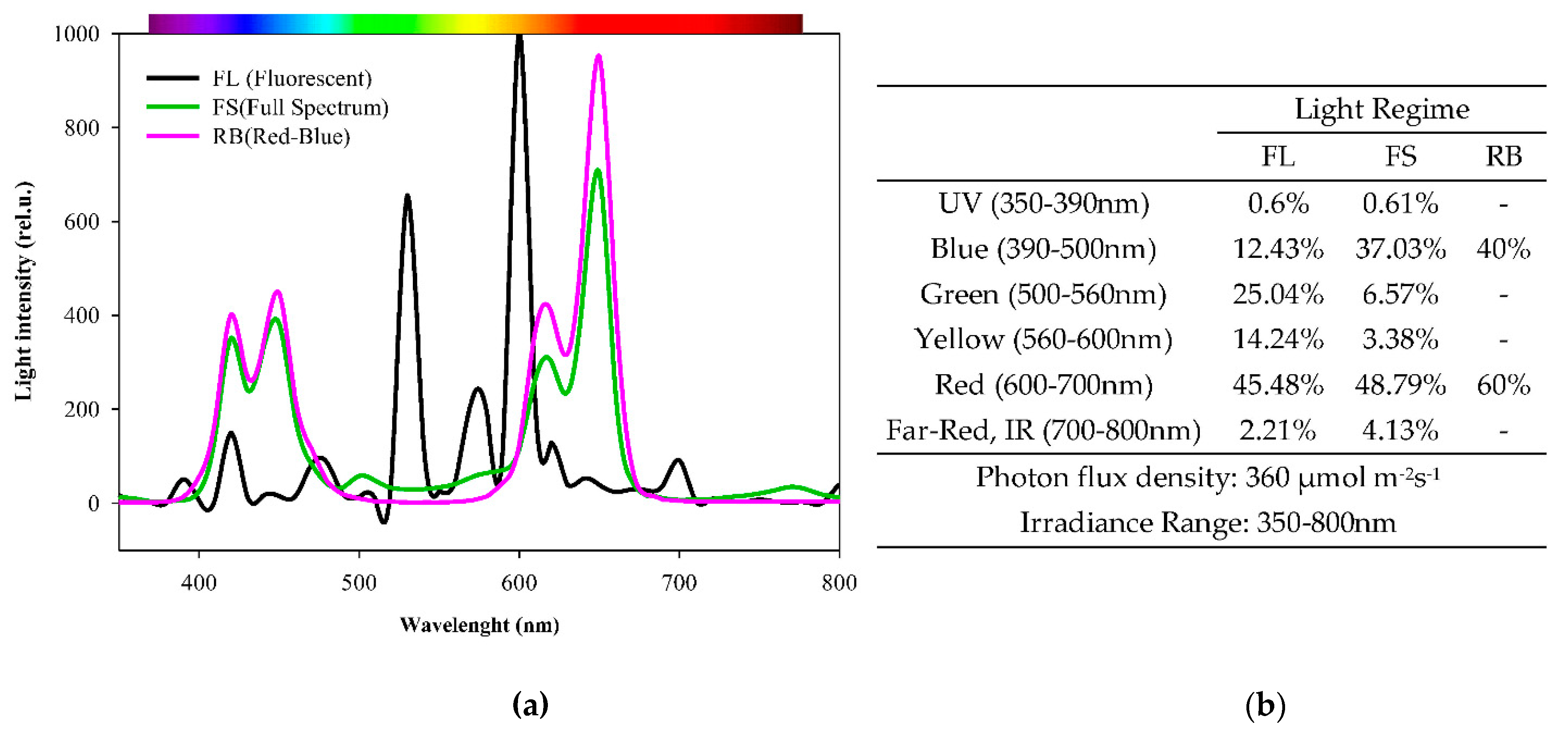
4.2. Growth Conditions
4.3. Analyses on Soybean Sprouts
Biochemical Analyses
4.4. Analyses on Soybean Seedlings
4.4.1. Morphological Parameters and Leaf Functional Attributes
4.4.2. Leaf Anatomy
4.4.3. Chlorophyll a Fluorescence Emission Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Derkzen, M.L.; Nagendra, H.; Van Teeffelen, A.J.; Purushotham, A.; Verburg, P.H. Shifts in ecosystem services in deprived urban areas: Understanding people’s responses and consequences for well-being. Ecol. Soc. 2017, 22, 51. [Google Scholar] [CrossRef] [Green Version]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Van Iersel, M.W. Optimizing LED Lighting in Controlled Environment Agriculture. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Taulavuori, E.; Taulavuori, K.; Holopainen, J.K.; Julkunen-Tiitto, R.; Acar, C.; Dincer, I. Targeted use of LEDs in improvement of production efficiency through phytochemical enrichment. J. Sci. Food Agric. 2017, 97, 5059–5064. [Google Scholar] [CrossRef]
- Shamshiri, R.R.; Kalantari, F.; Ting, K.C.; Thorp, K.R.; Hameed, I.A.; Weltzien, C.; Ahmad, D.; Shad, Z.M. Advances in greenhouses automation and controlled environment agriculture: A transition to plant factories and urban agriculture. Int. J. Agric. Biol. Eng. 2018, 11, 1–22. [Google Scholar] [CrossRef]
- Son, K.H.; Lee, J.H.; Oh, Y.; Kim, D.; Oh, M.M.; In, B.C. Growth and Bioactive Compound Synthesis in Cultivated Lettuce Subject to Light-quality Changes. HortScience 2017, 52, 584–591. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in con- trolled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Growth, photo-synthetic and antioxidant parameters of two lettuce cultivars as affected by red, green, and blue light-emitting diodes. Hortic. Environ. Biotechnol. 2015, 56, 639–653. [Google Scholar] [CrossRef]
- Son, K.H.; Park, J.H.; Kim, D.; Oh, M.M. Leaf shape, growth, and phytochemicals in two leaf lettuce cultivars grown under mono- chromatic light-emitting diodes. Korean J. Hortic. Sci. Technol. 2012, 30, 664–672. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-response of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Hernández, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Hasan, M.M.; Bashir, T.; Gosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the role of Red:Blue LED Lights on Resources Use Efficiency and Nutritional Properties of Indoor Grown Sweet Basil. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [Green Version]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red lighting strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta Gupta, L. Light Emitting Diodes for Agricolture. Smart Lighting; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Rouphael, J.; Colla, G. Toward a Sustainable Agriculture Through Plant Biostimulants: From Experimental Data to Practical Applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Vitale, L.; Vitale, E.; Guercia, G.; Turano, M.; Arena, C. Effects of different light quality and biofertilizers on structural and physiological traits of spinach plants. Photosynthetica 2020, 58, 932–943. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulant: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Techno. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- EBIC. European Biostimulants Industry Council. 2018. Available online: www.biostimulants.eu (accessed on 27 March 2021).
- Botta, A. Enhancing plant tolerance to temperature stress with amino acids: An approach to their mode of action. Acta Hortic. 2012, 1009, 29–36. [Google Scholar] [CrossRef]
- Kocira, S. Effect of amino acid biostimulant on the yield and nutraceutical potential of soybean. Chil. J. Agric. Res. 2019, 79, 17–25. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Hara, P.; Treder, K.; Findura, P.; Bartoš, P.; Filip, F. Biochemical and economical effect of application biostimulants containing seaweed extracts and amino acids as an element of agroecological management of bean cultivation. Sci. Rep. 2020, 10, 17759. [Google Scholar] [CrossRef]
- Hristozkova, M.; Geneva, M.; Stancheva, I.; Velikova, V. LED spectral composition effects on mycorrhizal symbiosis formation with tomato plants. Appl. Soil Ecol. 2017, 120, 189–196. [Google Scholar] [CrossRef]
- Ebert, A.W.; Chang, C.H.; Yan, M.R.; Yang, R.Y. Nutritional composition of mung bean and soybean sprouts compared to their adult growth stage. Food Chem. 2017, 237, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activities of two protein hydrolysates on the growth and nitrogen metabolism in maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Kunicki, E.; Grabowska, A.; Sekara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Gajc-Wolska, J.; Kowalczyk, K.; Nowecka, M.; Mazur, K.; Metera, A. Effect of organic-mineral fertilizers on the yield and quality of endive (Cichorium endivia L.). Acta Sci. Pol. Hortorum Cultus 2012, 11, 189–200. [Google Scholar]
- Trevisan, S.; Manoli, A.; Quaggiotti, S. A Novel Biostimulant, Belonging to Protein Hydrolysates, Mitigates Abiotic Stress Effects on Maize Seedlings Grown in hydroponics. Agronomy 2019, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Mahakman, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosybthesized silver nanoparticles. Sci. Rep. 2013, 7, 8263. [Google Scholar] [CrossRef]
- Afzal, I.; Hussain, B.; Basra, S.M.A.; Rehman, H. Priming with moring leaf extract reduced imbibitional chilling injury in spring maize. Seed Sci. Technol. 2012, 40, 271–276. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming memory invokes seed stress-tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Dai, L.Y.; Zhu, H.D.; Yin, K.D.; Du, J.D.; Zhan, Y.X. Seed priming mitigates the effects of saline-alkali stress in soybean seedlings. Chil. J. Agric. Res. 2017, 77. [Google Scholar] [CrossRef]
- Ugena, L.; Hylova, A.; Podlesakova, K.; Humplik, J.F.; Dolezak, K.; De Diego, N.; Spichal, L. Characterization of Biostimulant Mode of Action Using Novel-Multi-TraitHigh-Troughput Screening of Arabidopsis Germination and Rosette Growth. Front. Plant Sci. 2018, 13, 1327. [Google Scholar] [CrossRef] [Green Version]
- Alhadi, F.A.; Adnan, A.S.; AL, A.; Arif, S.A.A.; Qais, A.A.A. The effects of free amino acids profiles on seeds germination/dormancy and seedlings development of two genetically different cultivars of Yemeni Pomegranates. J. Stress Physiol. Biochem. 2012, 8, 114–137. [Google Scholar]
- Kim, I.D.; Dhungana, S.K.; Kim, J.H.; Ahn, H.; Kim, H.R.; Shin, D.H. Enhancement of yield and nutritional value of soybean sprouts by persimmon fruit powder. Afr. J. Biotechnol. 2016, 15, 2490–2496. [Google Scholar] [CrossRef] [Green Version]
- Kałużewicz, A.; Gąsecka, M.; Spiżewski, T. Influence of biostimulants on phenolic content in broccoli heads directly after harvest and after storage. Folia Hortic. 2017, 29, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulates short term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Yuan, M.; Jia, X.; Ding, C.; Zeng, H.; Du, L.; Yuan, S.; Zhang, Z.; Wu, Q.; Hu, C.; Liu, J. Effect of Fluorescence Light on Phenolic Compounds and Antioxidant Activities of Soybeans (Glycine max L. Merrill) during Germination. Food Sci. Biotechnol. 2015, 24, 1859–1865. [Google Scholar] [CrossRef]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Dipierro, N.; Paciolla, C. Effects of Darkness and Light Spectra on Nutrients and Pigments in Radish, Soybean, Mung Bean and Pumpkin Sprouts. Antioxidants 2020, 9, 558. [Google Scholar] [CrossRef]
- Chen, Y.; Chang, S.C.K. Macronutrients, Phytochemicals, and Antioxidant Activity of Soybean Sprout Germinated with or without Light Exposure. J. Food Sci. 2015, 80, S1391–S1398. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Cuong, D.M.; Ha, T.W.; Park, C.H.; Kim, N.S.; Yeo, H.J.; Chun, S.W.; Kim, C.; Park, S.U. Effects of LED lights on Expression of Genes Involved in Phenylpropanoid Biosynthesis and Accumulation of Phenylpropanoids in Wheat Sprout. Agronomy 2019, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Hori, A.; McCue, P.; Shetty, K. Seed vigour studies in corn, soybean and tomato in response to fish protein hydrolysates and consequences on phenolic-linked responses. Bioresour. Technol. 2007, 98, 2170–2177. [Google Scholar] [CrossRef]
- Poorter, H.; De Jong, R. A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity. New Phytol. 1999, 143, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving Spinach, Radish, and Lettuce Growth under Red Light emitting Diodes (LEDs) with Blue Light Supplementation. Hortic. Sci. 2001, 36, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Canamero, R.C.; Bakrim, N.; Bouly, J.P.; Garay, A.; Dudkin, E.E.; Habricot, Y.; Ahmad, M. Cryptochrome photoreceptors cry1 and cry2 antagonistically regulate primary root elongation in Arabidopsis thaliana. Planta 2006, 224, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Dougher, T.A.O.; Bugbee, B. Long-term blue light effects on the histology of lettuce and soybean leaves and stems. J. Am. Soc. Hort Sci. 2004, 129, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Dutta Gupta, S.; Barman, M.; Mitra, A. Photosynthetic apparatus plays a central role in photosensitive physiological acclimations affecting spinach (Spinacia oleracea L.) growth in response to blue and red photon flux ratios. Environ. Exp. Bot. 2018, 156, 170–182. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; De Micco, V.; Giordano, M.; Aronne, G.; De Pascale, S. Changes in Leaf Anatomical Traits Enhanced Photosynthetic Activity of Soybean Grown in Hydroponics with Plant Growth-Promoting Microorganisms. Front. Plant Sci. 2017, 8, 674. [Google Scholar] [CrossRef] [Green Version]
- Demming-Adams, B.; Garab, G.; Adams, W.W., III. Advances in Photosynthesis and Respiration Including Bioenergy and Related Processes. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cuanobacteria; Demming-Adams, B., Garab, G., Adams, W.W., III, Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2014; Volume 40. [Google Scholar] [CrossRef] [Green Version]
- Izzo, L.G.; Hay Mele, B.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Brown, C.S.; Stryjewski, E.C. Anatomical features of pepper plants (Capsicum annum L.) grown under light-emitting diodes supplemented with blue or far-red light. Ann. Bot. 1997, 79, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Tsonev, T.; Doneva, D.; De Micco, V.; Michelozzi, M.; Brunetti, C.; Centritto, M.; Fineschi, S.; Velikova, V.; Loreto, F. The effect of light quality on growth, photosynthesis, leaf anatomy and volatile isoprenoids of a monoterpene-emitting herbaceous species (Solanum lycopersicum L.) and an isoprene-emitting tree (Platanus orientalis L.). Environ. Exp. Bot. 2016, 130, 122–132. [Google Scholar] [CrossRef]
- Oguchi, R.; Hikosaka, K.; Hirose, T. Does the photosynthetic light-acclimation need change in leaf anatomy? Plant Cell Environ. 2003, 26, 505–512. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Vogelmann, T.C. Do changes in light direction affect absorption profiles in leaves? Funct. Plant Biol. 2010, 37, 403–412. [Google Scholar] [CrossRef]
- Smith, H.L.; Mc Ausland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse role in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Pintilie, O.; Stoleru, T.; Burducea, M.; Oroian, M.; Zamfirache, M.M. Blue and red LED illumination improves growth and bioactive compounds contents in acyanic and cyanic Ocimum basilicum L. microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef] [Green Version]
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Thu, N.T.; Hien, N.T.T.; Khanh, T.D.; Ha, C.H. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes. Plants 2020, 9, 1344. [Google Scholar] [CrossRef]
- Carvalho, K.F. Green light control of anthocyanin production in microgreens. Acta Hortic. 2016, 1134, 13–18. [Google Scholar] [CrossRef]
- Hofmann, R.W.; Jahufer, M.Z.Z. Tradeoff between biomass and flavonoid accumulation in white clover reflects contrasting plant strategies. PLoS ONE 2011, 6, e18949. [Google Scholar] [CrossRef] [Green Version]
- Hatier, J.H.B.; Clearwater, M.J.; Gould, K.S. The functional significance of black-pigmented leaves: Photosynthesis, photoprotection and productivity in Ophiopogon planiscapus ‘Nigrescens’. PLoS ONE 2013, 8, e67850. [Google Scholar] [CrossRef] [Green Version]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fang, S.; Yang, W.; Shang, X.; Fu, X. Light quality affects flavonoid production and related gene expression in Cyclocarva paliurus. J. Photochem. Photobiol. B 2018, 179, 66–73. [Google Scholar] [CrossRef]
- Noman, A.; Ali, Q.; Maqsood, J.; Iqbal, N.; Javed, M.T.; Rasool, N.; Naseem, J. Deciphering physio-biochemical, yield, and nutritional quality attributes of water-stressed radish (Raphanus sativus L.) plants grown from Zn-Lys primed seeds. Chemosphere 2018, 195, 175–189. [Google Scholar] [CrossRef]
- George, B.; Kaur, C.; Khurdiya, D.S.; Kapoor, H.C. Antioxidants in tomato (Lycopersicum esculentum) as a function of genotype. Food Chem. 2004, 84, 45–51. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, E.; Hay Mele, B.; Cataletto, P.R.; Turano, M.; Simoniello, P.; De Micco, V. Suitability of Solanum lycopesicum L. ‘Microtom’ for growth in Bioregenerative Life Support Systems: Exploring the effect of high-LET ionising radiation on photosynthesis, leaf structure and fruit trait. Plant Biol. 2019, 21, 615–626. [Google Scholar] [CrossRef]
- Costanzo, G.; Iesce, M.R.; Naviglio, D.; Ciaravolo, M.; Vitale, E.; Arena, C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants 2020, 9, 517. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method. Enzymol. 1987, 148, 350–382. [Google Scholar]
- Hedge, J.E.; Hofreiter, B.T. Carbohydrate chemistry. In Methods in Carbohydrate Chemistry; Whistler, R.L., Be Miller, J.N., Eds.; Academic Press: New York, NY, USA, 1962; Volume 17. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Im, J.; Ko, J.; Kim, H.Y.; Ha, B.K. Biochemical Responses of Soybean (Glycine max L. Merr.) to Proton Beam Irradiation. Plant Breed. Biotechnol. 2017, 5, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Soukup, A.; Tylová, E. Essential Methods of Plant Sample Preparation for Light Microscopy. Methods Mol. Biol. 2019, 1992, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Compounds | |||||||
|---|---|---|---|---|---|---|---|
| TAC | TPC | CHL | CAR | CARB | AsA | SP | |
| B | |||||||
| H2O | 1.48 b | 0.77 c | 0.33 b | 0.034 a | 59 c | 7.5 c | 55 c |
| K-0.01% | 1.59 a | 0.92 a | 0.30 b | 0.038 a | 65 b | 12 b | 60 b |
| K-0.05% | 1.62 a | 0.95 a | 0.44 a | 0.037 a | 82 a | 15 a | 72 a |
| K-0.5% | 1.58 a | 0.88 b | 0.44 a | 0.035 a | 69 b | 15 a | 70 c |
| LQ | |||||||
| D | 1.40 b | 0.62 c | 0.21 c | 0.028 c | 67 a | 9.7 c | 59 c |
| FL | 1.60 a | 0.83 b | 0.40 b | 0.023 b | 71 a | 12 b | 72 a |
| FS | 1.64 a | 1.02 a | 0.45 a | 0.042 a | 68 a | 14 a | 63 b |
| RB | 1.64 a | 1.04 a | 0.46 a | 0.042 a | 67 a | 14 a | 63 b |
| Interaction | |||||||
| H2O × D | 1.72 a | 0.56 e | 0.20 c | 0.028 b | 57 c | 5.3 d | 61 c |
| K-0.01% × D | 1.47 bc | 0.59 e | 0.22 c | 0.029 b | 62 c | 7.4 c | 54 d |
| K-0.05% × D | 1.41 bc | 0.69 d | 0.17 c | 0.024 b | 92 a | 12 b | 56 d |
| K-0.5% × D | 1.02 d | 0.64 e | 0.23 c | 0.031 b | 60 c | 14 b | 64 c |
| H2O × FL | 1.51 b | 0.79 d | 0.39 b | 0.031 b | 67 b | 7.2 c | 54 d |
| K-0.01% × FL | 1.58 b | 0.74 d | 0.38 b | 0.028 b | 62 c | 13 b | 63 c |
| K-0.05% × FL | 1.56 b | 0.93 c | 0.42 b | 0.038 ab | 80 b | 14 b | 81 b |
| K-0.5% × FL | 1.74 a | 0.85 c | 0.40 b | 0.033 b | 75 b | 14 b | 89 a |
| H2O × FS | 1.34 c | 0.85 c | 0.38 b | 0.038 ab | 57 c | 8.8 c | 51 d |
| K-0.01% × FS | 1.65 a | 1.11 b | 0.31 b | 0.048 a | 69 b | 13 b | 60 c |
| K-0.05% × FS | 1.74 a | 1.11 b | 0.56 a | 0.043 a | 77 b | 16 a | 76 b |
| K-0.5% × FS | 1.77 a | 1.02 b | 0.56 a | 0.038 ab | 71 b | 17 a | 65 c |
| H2O × RB | 1.35 c | 0.88 c | 0.35 b | 0.039 ab | 55 c | 8.8 c | 52 d |
| K-0.01% × RB | 1.68 a | 1.25 a | 0.30 b | 0.049 a | 67 b | 14 b | 62 c |
| K-0.05% × RB | 1.76 a | 1.05 b | 0.59 a | 0.043 a | 77 b | 17 a | 77 b |
| K-0.5% × RB | 1.76 a | 0.99 b | 0.59 a | 0.038 ab | 69 b | 17 a | 63 c |
| Significance | |||||||
| B | *** | *** | *** | NS | *** | *** | *** |
| LQ | *** | *** | *** | *** | NS | *** | *** |
| B × LQ | *** | *** | *** | ** | *** | *** | *** |
| Morphological Parameters | Anatomical Traits | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| TLA | SLA | Lenght | Biomass | S/R | LT | PT | ST | IS | |
| B | |||||||||
| H2O | 20 a | 221 b | 40 a | 0.37 a | 1.47 a | 140 a | 71 a | 48 a | 14 a |
| K-0.01% | 22 a | 215 b | 38 a | 0.37 a | 1.67 a | 134 b | 74 a | 45 a | 15 a |
| K-0.05% | 21 a | 286 a | 40 a | 0.35 a | 1.41 a | 130 c | 73 a | 45 a | 14 a |
| K-0.5% | 23 a | 199 b | 36 a | 0.35 a | 1.37 a | 127 c | 68 b | 45 a | 14 a |
| LQ | |||||||||
| FL | 25 a | 214 b | 38 a | 0.37 a | 1.72 a | 124 c | 69 b | 41 b | 14 a |
| FS | 20 b | 240 a | 41 a | 0.35 a | 1.26 b | 132 b | 70 b | 47 a | 15 a |
| RB | 19 b | 237 a | 37 a | 0.36 a | 1.45 b | 143 a | 76 a | 49 a | 13 b |
| Interaction | |||||||||
| H2O × FL | 21 a | 210 b | 45 a | 0.38 a | 1.89 a | 125 c | 66 d | 41 b | 17 ab |
| K-0.01% × FL | 24 a | 198 b | 40 a | 0.37 a | 1.73 a | 124 c | 67 d | 42 b | 20 ab |
| K-0.05% × FL | 29 a | 271 a | 35 a | 0.36 a | 1.88 a | 131 bc | 82 b | 41 b | 14 b |
| K-0.5% × FL | 27 a | 179 c | 34 a | 0.35 a | 1.41 a | 117 d | 62 e | 39 b | 10 c |
| H2O × FS | 21 a | 206 b | 40 a | 0.38 a | 1.11 a | 147 a | 71 cd | 51 a | 17 ab |
| K-0.01% × FS | 22 a | 224 b | 37 a | 0.37 a | 1.73 a | 127 c | 68 cd | 43 b | 11 bc |
| K-0.05% × FS | 15 a | 299 a | 49 a | 0.33 a | 1.13 a | 129 bc | 70 cd | 52 a | 13 b |
| K-0.5% × FS | 23 a | 232 b | 37 a | 0.37 a | 1.10 a | 126 c | 70 cd | 44 b | 20 a |
| H2O × RB | 19 a | 248 b | 34 a | 0.36 a | 1.41 a | 149 a | 75 c | 51 a | 8.3 c |
| K-0.01% × RB | 20 a | 226 b | 41 a | 0.39 a | 1.56 a | 152 a | 87 a | 51 a | 18 a |
| K-0.05% × RB | 19 a | 291 a | 35 a | 0.35 a | 1.22 a | 132 bc | 70 cd | 42 b | 14 b |
| K-0.5% × RB | 18 a | 185 c | 39 a | 0.39 a | 1.62 a | 139 b | 72 cd | 54 a | 12 b |
| Significance | |||||||||
| B | NS | *** | NS | NS | NS | *** | *** | NS | NS |
| LQ | * | * | NS | NS | *** | *** | *** | *** | ** |
| B × LQ | NS | * | NS | NS | NS | *** | *** | *** | *** |
| Pigments | Functional Traits | ||||||
|---|---|---|---|---|---|---|---|
| CHL | FLAV | ANTH | NBI | ΦPSII | NPQ | Fv/Fm | |
| B | |||||||
| H2O | 37 a | 1.32 a | 0.208 a | 28 a | 0.45 b | 1.46 b | 0.741 b |
| K-0.01% | 36 a | 1.28 a | 0.207 a | 29 a | 0.44 b | 1.29 c | 0.748 b |
| K-0.05% | 38 a | 1.27 a | 0.205 a | 30 a | 0.51 a | 1.12 d | 0.765 a |
| K-0.5% | 34 b | 1.29 a | 0.211 a | 27 a | 0.41 c | 1.59 a | 0.750 b |
| LQ | |||||||
| FL | 34 b | 1.23 b | 0.210 a | 29 a | 0.36 b | 1.74 a | 0.753 a |
| FS | 37 a | 1.28 b | 0.204 b | 29 a | 0.49 a | 1.26 b | 0.751 a |
| RB | 37 a | 1.38 a | 0.211 a | 26 b | 0.51 a | 1.10 c | 0.747 a |
| Interaction | |||||||
| H2O × FL | 35 a | 1.23 b | 0.205 bc | 29 ac | 0.40 c | 1.88 b | 0.726 b |
| K-0.01% × FL | 32 b | 1.25 b | 0.213 ab | 26 bc | 0.28 d | 1.89 b | 0.749 ab |
| K-0.05% × FL | 40 a | 1.15 b | 0.198 c | 35 a | 0.51 ab | 1.09 d | 0.784 a |
| K-0.5% × FL | 30 b | 1.29 b | 0.222 a | 27 bc | 0.26 d | 2.11 a | 0.754 ab |
| H2O × FS | 40 a | 1.32 ab | 0.202 bc | 31 ab | 0.46 b | 1.39 c | 0.735 ab |
| K-0.01% × FS | 37 a | 1.28 ab | 0.207 bc | 29 ac | 0.52 ab | 1.02 d | 0.743 ab |
| K-0.05% × FS | 37 a | 1.16 b | 0.203 bc | 33 ab | 0.51 ab | 1.09 d | 0.773 ab |
| K-0.5% × FS | 34 a | 1.35 a | 0.203 bc | 28 bc | 0.45 b | 1.51 c | 0.754 ab |
| H2O × RB | 34 a | 1.43 a | 0.218 ab | 24 c | 0.50 ab | 1.10 d | 0.762 ab |
| K-0.01% × RB | 40 a | 1.33 ab | 0.202 bc | 30 ac | 0.53 a | 0.97 d | 0.751 ab |
| K-0.05% × RB | 33 a | 1.49 a | 0.216 ab | 22 c | 0.49 ab | 1.19 d | 0.735 ab |
| K-0.5% × RB | 37 a | 1.24 b | 0.209 bc | 30 ac | 0.51 ab | 1.15 d | 0.741 ab |
| Significance | |||||||
| B | * | NS | NS | NS | *** | *** | * |
| LQ | * | *** | ** | * | *** | *** | NS |
| B × LQ | *** | *** | *** | *** | *** | *** | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, E.; Velikova, V.; Tsonev, T.; Ferrandino, I.; Capriello, T.; Arena, C. The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill). Plants 2021, 10, 861. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050861
Vitale E, Velikova V, Tsonev T, Ferrandino I, Capriello T, Arena C. The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill). Plants. 2021; 10(5):861. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050861
Chicago/Turabian StyleVitale, Ermenegilda, Violeta Velikova, Tsonko Tsonev, Ida Ferrandino, Teresa Capriello, and Carmen Arena. 2021. "The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill)" Plants 10, no. 5: 861. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050861