1. Introduction
The enhancement and maintenance of soil productivity and sustainability through the long-term use of different silvicultural practices, such as varying planting density and spacing, introducing native and exotic beneficial species, agroforestry and intercropping, has been widely used worldwide [
1,
2,
3,
4,
5,
6]. Intercropping can enhance soil quality by incorporating a significant amount of topsoil and subsoil organic matter and releasing and recycling nutrients [
7,
8,
9]. It is the general opinion that intercropping is more suitable than mono-cropping for the long-term maintenance of soil fertility [
10]. Intercropping systems are rarely dependent on external inputs such as fertilizers [
11]; this also reduces the adverse environmental effects such as soil erosion [
12]. In China, trees/legume intercropping is most commonly used because it reduces crop failure risk and improves land use [
13].
Tea (
Camellia oleifera) is an evergreen shrub of the Theaceae family, and it is widely found in central and south China [
14]. Moreover, it can also survive in nutrient-depleted soils. Peanut (
Arachishypogaea L.) is a leguminous crop that can fix the atmospheric N and increase soil fertility [
15]. According to the US Department of Agriculture, it produces a considerable amount of organic N, improves soil organic matter (SOM), helps in nutrient release and recycling, and improves soil structure. When peanut was intercropped with maize, it played a crucial role in changing soil health by influencing soil microbes composition [
16]. Additionally, the dominant microbial species altered due to the peanut intercropping, which shows a close and significant relationship between improving available soil nutrients and soil enzyme activities [
17].
Soil enzymes are continually playing dynamic roles in the maintenance of soil health. Soil enzymes are the direct mediators for the biological catabolism of soil organic and mineral components. They are often closely associated with SOM, soil physical properties, and microbial activities and biomass. They are the better indicators of soil health as changes in enzymes occurred much earlier than other soil parameters, thus providing early indications of changes in soil health. Their activities can also be used as measures of microbial activity and soil productivity [
18,
19]. Although they are present in a very nominal quantity, their role in soil quality can never be ignored. Likewise, a soil nutrient’s total content and soil nutrient availability directly affect the plant’s growth and development, reflecting soil health [
20,
21,
22]. In the southern Chinese province of Hunan, the adoption of agroforestry (intercropping) in tea interests the growers because intercropping controls weed growth and soil erosion. However, many of the sites where these plantations were established are deficient in available macronutrients, albeit a high level of total phosphorus (P). The available P rapidly forms insoluble complexes with cations, particularly aluminum and iron. Less nutrient availability in forest soils is considered as one of the most important causes of productivity decline [
23]. Nutrient total status and availability are essential for plant growth and development, and thus, macronutrient availability is vital for sustainable productivity. Soil enzymes play a significant role in nutrient recycling in soil ecosystems. In addition, soil enzymes play a crucial role in maintaining soil quality. They can provide essential and early detection signals for soil metabolic activity and nutrient status changes. However, few publications have focused on how legume intercropping with tea plantation, specifically related soil enzymatic activities, influences soil fertility.
Therefore, the objectives of our current study were (1) to investigate the soil total nutrient content and soil nutrient availability; (2) and the activities of soil enzymes (sucrase, protease, urease, acid phosphatase, and catalase); and (3) to explore a correlation between soil enzymatic activity and soil nutrients in different cropping systems and soil depths. Three cropping systems, including peanut–tea intercropping, tea mono-cropping, peanut mono-cropping, and four different soil depths (0–10 cm, 10–20 cm, 20–30 cm, and 30–40 cm) were used in this study. Our hypotheses were that: (i) intercropping favors better nutrient availability and total nutrient content than monoculture systems due to better litter quality and decomposition, and nutrient availability will decrease with the increase in soil depth; (ii) soil enzymatic activity will be higher in the peanut–tea intercropping system than in tea monoculture because peanut influences soil microbial composition—thus, it can enhance soil microbial activity, which has a significant relationship with the improvement of soil enzyme activities; and (iii) there will be a positive and considerable relationship between soil nutrients and soil enzymatic activity across all cropping types and soil depths.
3. Discussion
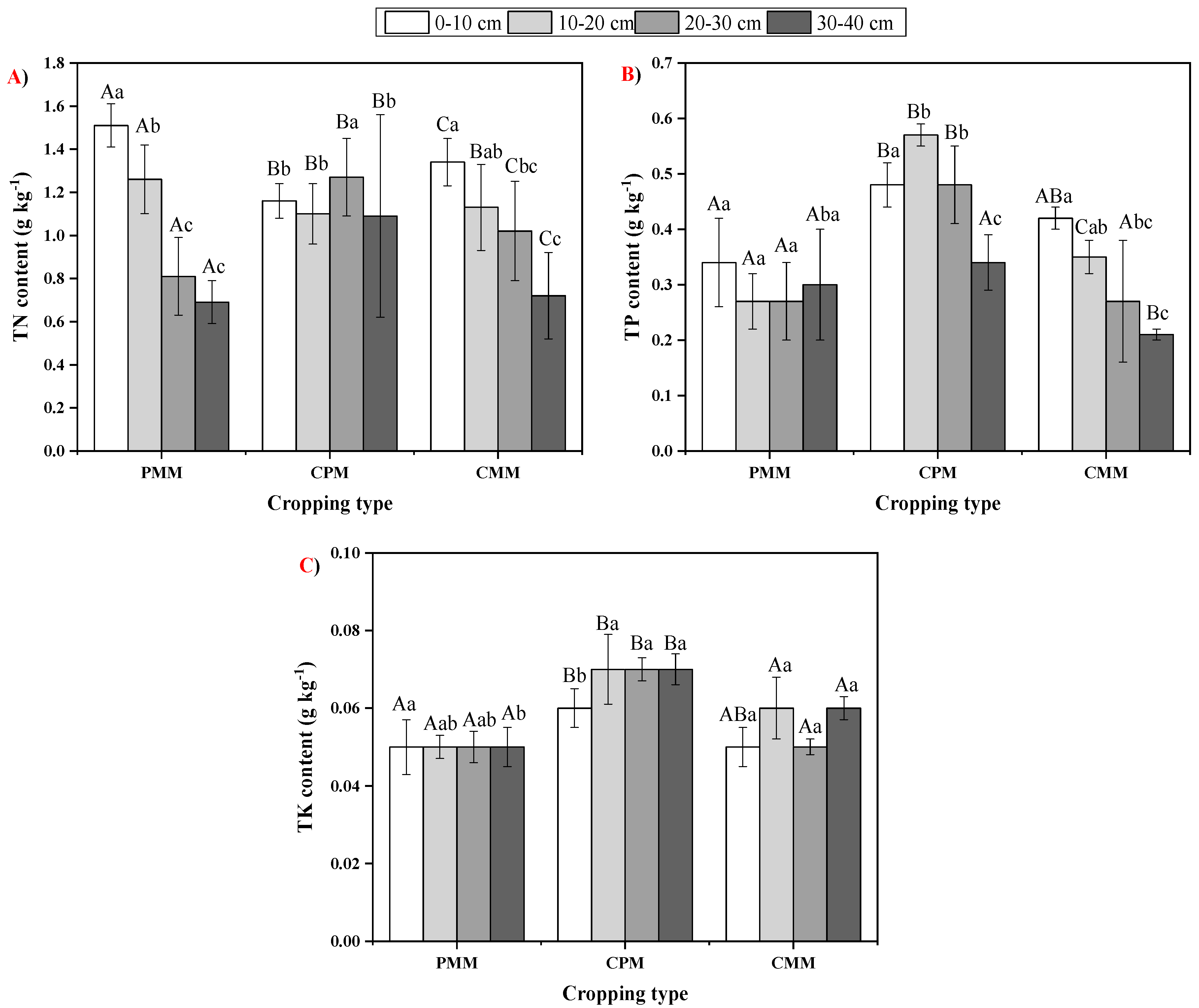
In the present study, SOC was unanimously higher in the 0–10 cm soil layer among all the cropping systems because of the higher litterfall on the topmost layer. Compared to the mono-cropping of tea, SOC was higher in peanut–tea intercropping, possibly due to the higher litterfall and tea biomass decomposition inputs. Growing different crops on the same land simultaneously helps maintain the SOC and improve nutrient cycling [
24,
25]. A similar kind of increase in tea soil SOC content was also observed by [
26,
27]. N is vital for plant growth and development [
22,
28]. It is available in many soil forms, such as nitrate and ammonia [
29,
30]. Peanut being a legume plant could fix atmospheric nitrogen [
31,
32] and as a result, we found out that PMM had the highest amount of TN for the upper two layers, while in the remaining two layers, it was observed to be highest in CPM. This could be due to the tea plant deep root system, which makes way in the soil deeper layers for the atmospheric N that the peanut plant fixes. Both TP and TK contents were higher in the CPM among all the soil layers than the tea and peanut mono-cropping system. The probable reason was that intercropping can provide a much more comprehensive ground cover along with better water use efficiency [
31,
32].
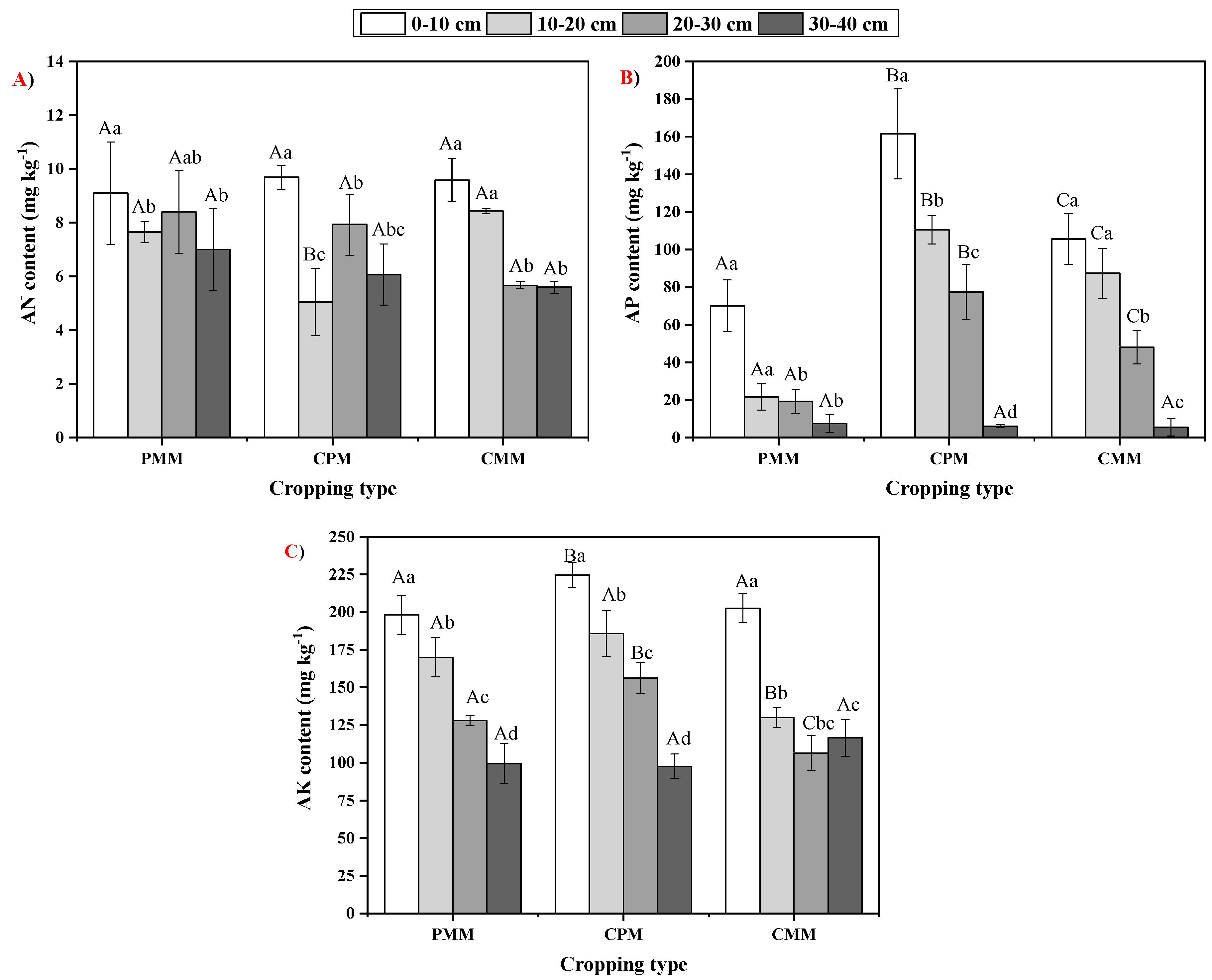
Leguminous plants can convert the unavailable N form into a useable form [
33]. The comparison of CPM and PMM depicted that AN was 51.78%, 5.92%, and 15.32% lower in the 10–20 cm, 20–30 cm, and 30–40 cm layers of the intercropping system than in the peanut mono-cropping system because the of higher urease activity in CPM, which led to an increase in the NH
3 loss from soil [
34,
35]. AP in the intercropping of tea and peanut was higher in all the soil layers than the tea and peanut monoculture cropping. This is because leguminous plant led to the acidification of the rhizosphere with the help of roots, and these roots release the organic acids [
36,
37,
38,
39]. Due to acidification, the enzyme acid phosphatase starts the dissolution of P-based minerals, which increases the P availability. Maurya and Lal [
40] obtained similar results, where the amount of P increased due to the release of acid phosphatase by the chickpea, which led to the conversion of organic P into inorganic P. AK content was higher in CPM compared to CMM and PMM in all soil layers. AK was 11.1%, 43.06%, 46.79% higher in 0–10 cm, 10–20 cm, and 20–30 cm layers of the intercropping system than the mono-cropping of tea. Similarly, AK was 13.4%, 9.29%, and 22.06% higher in 0–10 cm, 10–20 cm, and 20–30 cm layers of intercropping system than the mono-cropping system of peanut. The increase in AK in the intercropping system can be attributed to the increase in the soil enzymatic activity.
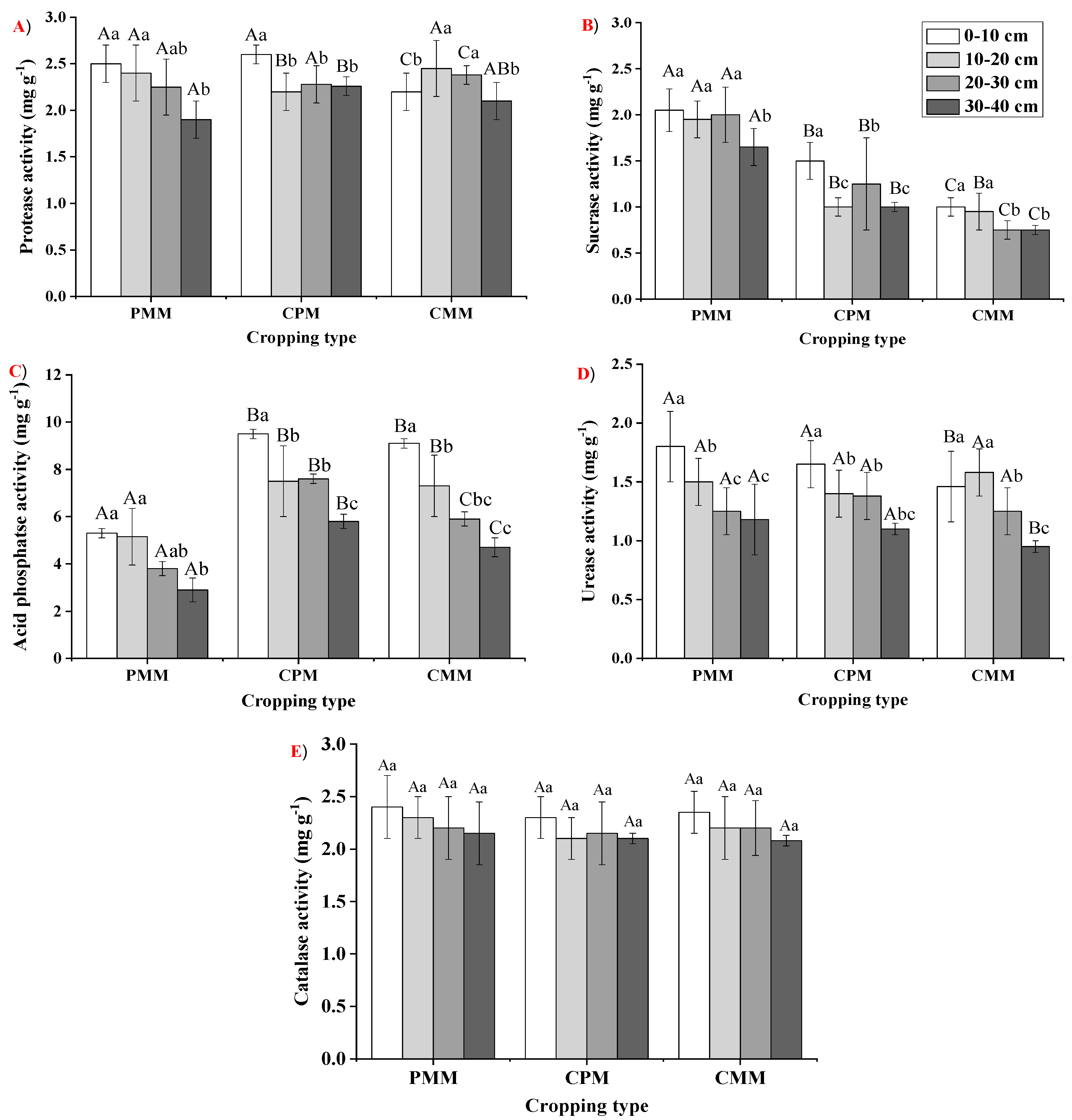
Soil enzymes play a significant role in the soil ecosystem’s overall biochemical functioning [
41,
42,
43]. A better understanding of the soil enzymatic activity in different cropping systems with a depth effect can provide better knowledge about how intercropping systems can improve soil fertility. It is evident from the previous studies that the mono-cropping system could potentially harm the soil enzyme mechanism, which results in a significant decrease in the soil enzymatic activity [
44]. In our current study, a very considerable increase in the enzyme activities was observed in the intercropping system of the tea and peanut than in the mono-cropping systems. Protease is the main enzyme involved in the catalysis of N minerals and N cycling [
45]. The protease activity was higher in the layers of 0–10 cm and 30–40 cm for the intercropping system than both mono-cropping systems. This change in protease activity might be attributed to a higher SOC content in the topsoil [
46]. However, in the 30–40 cm layer, the increase might be due to a higher N content. Soil sucrase enzymes catalyze sucrase to glucose and fructose with hydrolysis, and it is also connected with the biomass of the soil microbes [
47,
48]. The mono-cropping of peanut had the highest sucrase enzyme activity, possibly due to a higher SOM. These findings are supported by Li et al. [
41], who observed that soil enzyme activity was significantly enhanced due to intercropping. Acid phosphatase is the main enzyme involved in ester hydrolysis and phosphoric acid anhydrides [
49,
50]. It converts the esters and anhydrides into phosphate, and is a key enzyme in P cycling in soil [
51].
In our current study, acid phosphatase activity was higher in the intercropping of peanut–tea than tea mono-cropping because the peanut was identified as a species whose roots release an ample amount of acid phosphatase in the soil [
52,
53]. The soil urease enzyme catalyzes urea into NH
3 and CO
2 with hydrolysis [
54]. This is a necessary process that regulates N availability to the plants after the application of urea. To some level, urease indicates the availability of N in different cropping systems. The catalase’s primary function is to decompose the organic matter into the plant’s useable form [
55,
56]. In our current study, the mono-cropping of peanut had a higher activity of urease and catalase than intercropping because of the SOM content of the mono-cropping of peanut. In this study, the enzyme activity was more or less associated with the content and distribution of SOC, TN and AK among all the cropping systems. Our results are in line with Tian et al. [
57] and Udawatta et al. [
58]; they also observed a stronger correlation between SOC and enzymes. An increase in the SOM and litter quantity enhances the soil activity [
59,
60] and this increment has direct involvement in the improvement of nutrient cycling, along with greenhouse emission [
61,
62,
63], which, in return, has a positive impact on the ecosystem, plant growth and overall SOC.
4. Materials and Methods
4.1. Study Site
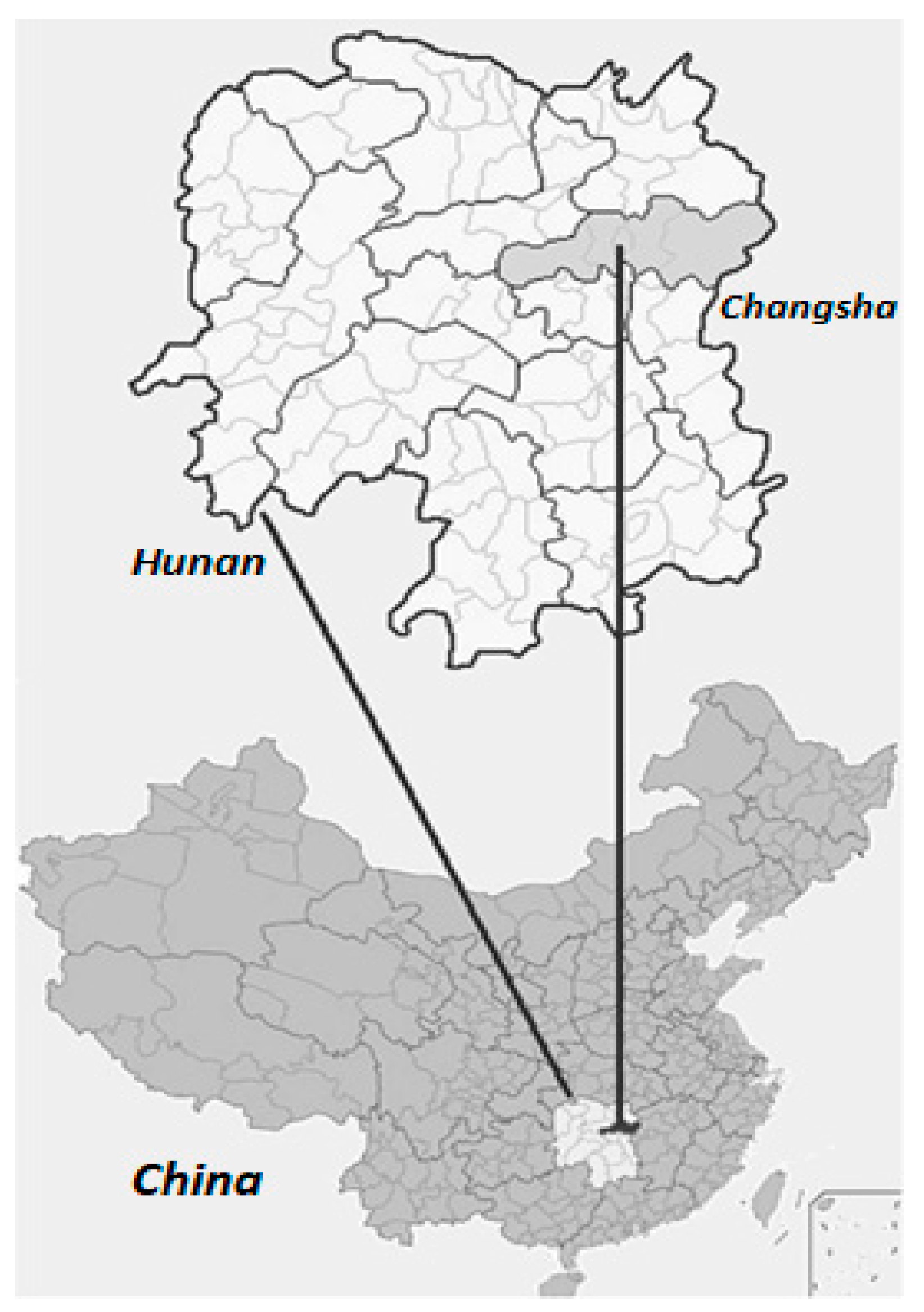
The study was conducted in Hunan Botanical Garden in Changsha city, Hunan Province, China (113°02′–113°03′ E, 28°06′–28°07′ N) (
Figure 4). The study area had a typical subtropical humid monsoon climate, with a mean annual precipitation of 1378 mm, mean annual temperature of 17.2 °C, and a mean annual relative humidity of 81%. The mean annual average sunshine was 1814.8 h and the frost-free period was 275 days. The site was the hillside, with a slope of about 5–15°. The soil was classified as typical red earth developed from the quaternary red clay reticulated parent material. The soil texture ranged from clay loam to sandy loam, with a depth of about 1 m. The soil was acid with a pH of 4.5–5.5. Soil bulk density was ranged between 1.16 to 1.22 (g/m
3).
4.2. Experimental Design
In this study, three cropping models were set up which included: (1) a C. oleifera mono-cropping model (CMM); (2) a peanut mono-cropping model (PMM), and (3) a Camellia–peanut inter-cropping model (CPM). The C. oleifera monoculture stand was established in 2010, and the line-row spacing of the C. oleifera was planted in 4 m × 3 m. In CMM, an area with three lines and three rows of C. oleifera trees was selected as a plot (about 110 m2). Four plots were set up for CMM in this study.
However, in PMM, the peanuts were planted in the open space beside the C. oleifera forests according to the 0.25 m × 0.1 m spacing in April 2018. An area with 40 lines and 100 rows of peanut plants was selected as a plot (about 100 m2). Four plots were set up for PMM in this study. Moreover, in CPM, an area with three lines and three rows of C. oleifera trees was selected as a plot (about 110 m2). Peanuts were intercropped between C. oleifera trees in April 2018. The spacing between the peanut and tea plant was 1 m. Four plots of CPM were set up in this study. The three cropping models employed similar field management practices during the study period.
4.3. Soil Sampling
Soil samples were collected in September 2019. Four plots of 20 × 20 m were established in each of CMM, PMM, and CPM plots; four pits were dug diagonally. A soil corer was used to obtain soil samples from 0–10, 10–20, 20–30, and 30–40 cm depths from each pit. Four replicates were taken from each plot for each depth. The rocks and plant residues were removed from the soil samples. Soil samples were placed into self-sealing plastic bags, labelled, and delivered to the laboratory for further analysis. For enzymatic activity analysis, the soil samples were placed in −80 °C until further use. For nutrients analysis, after air drying, the soil samples were passed through a 0.15 mm and 0.20 mm sieve for the determination of soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), total potassium (TK), available nitrogen (AN), available phosphorus (AP), and available potassium (AK).
4.4. Soil Chemical Analysis
Urease activity was measured following the method described in [
64]. Five grams of soil was incubated with 10 mL of citrate phosphate buffer (pH 6.7) and 5 mL of 10 % urea solution at 38 °C for 3 h. Activity was determined by measuring the released NH
4+ with a spectrophotometer at 578 nm. Acid phosphatase activity was analyzed with nitrophenyl phosphate disodium (PhOH mg g
1, 37 °C, 24 h), and catalase with KMnO
4 (0.1 mol L
−1 KMnO
4 ug g
−1, 30 °C, 20 h) [
65]. Sucrase activity was determined by the method of [
66]. For sucrose, the air-dried soil (5 g) was incubated with 15 mL sucrose. Five microliters (5 mL) of phosphate buffer (pH 5.5) and five drops of toluene at 37 °C for 24 h, and the reaction solution was filtered through the quantitative filter paper as rapidly as possible after incubation. Filtrate (1 mL) was mixed with 3 mL salicylic acid at 100 °C for 5 min in the water bath, and the mixture was adjusted to 50 mL and cooled with deionized water. Sucrase activity was determined spectrophotometrically at 508 nm. The protease activity was determined by ninhydrin colorimetry, expressed in milligrams of amino nitrogen in 1 g of soil cultured for 24 h in a 37 °C incubator [
67].
SOC was determined by the hydrated potassium thermo-dichromate oxidation method. TN was determined using the CN elemental analyzer, while the colorimetric method of the molybdenum–antimony solution with royal acid was used to determine TP. The flame photometer method was used to determine TK. AN was analyzed by the Kjeldahl method [
68]; AP was determined by the diacid extraction spectrophotometric colorimetry method [
69], and AK was determined using a flame photometer method by ammonium acetate extraction [
70]. Detailed information about the total and available nutrients was also mentioned in our published paper [
6].
4.5. Statistical Analysis
Two-way analysis of variance (ANOVA) was performed to analyze the difference in soil enzymatic activity, soil total nutrient status and soil nutrient availability between different cropping systems at four different soil depths. Correlation analysis was performed using the Pearson statistical method to analyze the association among the studied soil parameters. Moreover, the path analysis using multiple linear analysis was also performed to study the relationships betweesn soil enzyme and soil nutrients. Results were declared statistically significant at p < 0.05 and the means that exhibited significant differences were compared using Tukey’s significance test. All statistical analyses were performed using the SPSS Statistical Package (SPSS 17.0, Chicago, IL, USA).
The multiple linear regression equations of soil sucrase activity (Y1), protease activity (Y2), urease activity (Y3), acid phosphatase activity (Y4), catalase activity (Y5) and soil nutrient factors are as follows:
The multiple linear regression equations in PMM are:
The multiple linear regression equations in CPM are:
The multiple linear regression equations in CMM are:
X1, X2, X3, X4, X5, X6 and X7 represent SOC, TN, TP, TK, AN, AP and AK, respectively. These equations present the direct path coefficients. The indirect path coefficients were obtained using the direct path coefficients multiplied by each soil nutrient’s correlation coefficient. The direct path coefficient was the direct effect of soil nutrient factors on enzyme activity.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}



