
Foliar-Applied Potassium Silicate Coupled with Plant Growth-Promoting Rhizobacteria Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia faba L.) Irrigated with Saline Water in Salt-Affected Soil

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
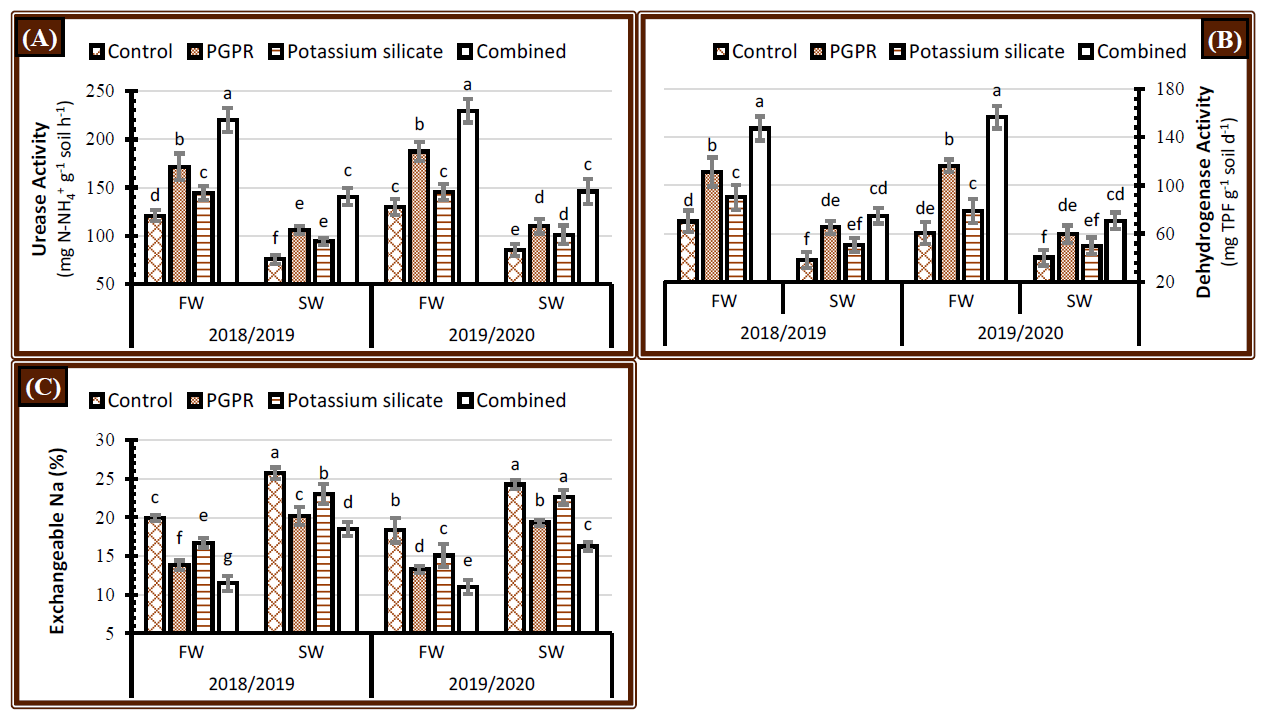
2.1. Soil Quality Indicators
2.1.1. Soil Enzyme Activity
2.1.2. Soil Exchangeable Na Percentage
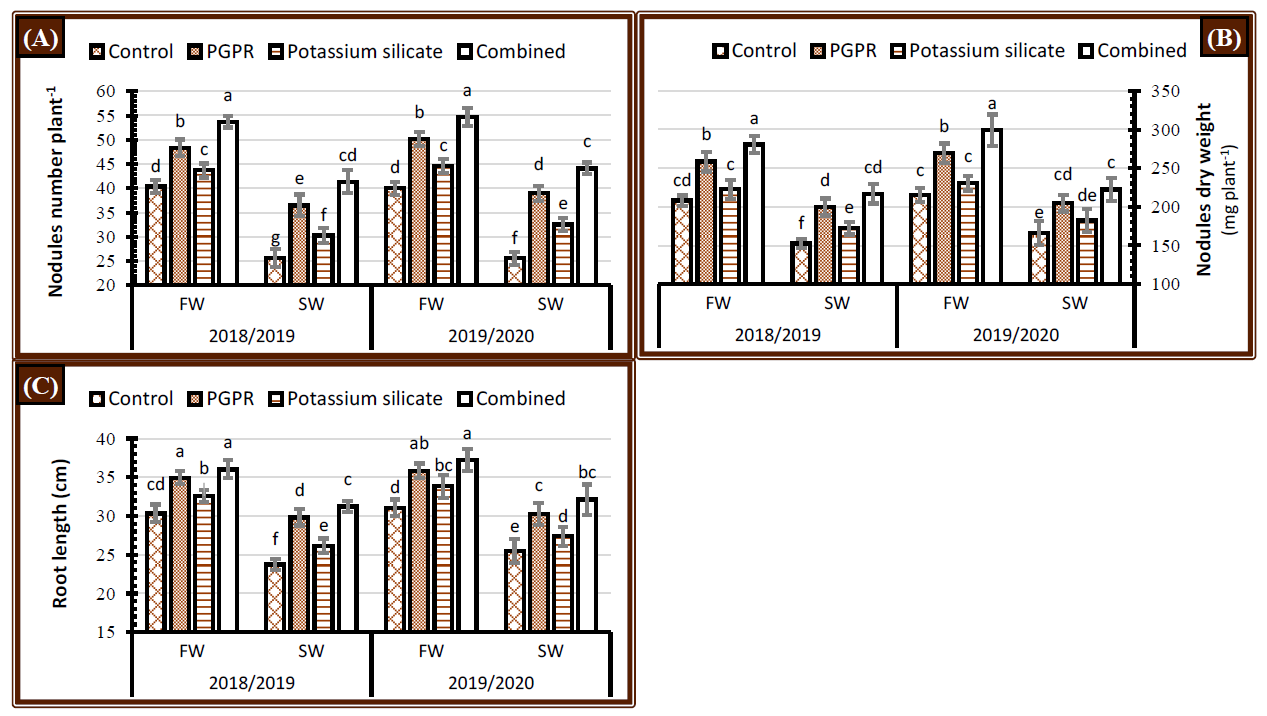
2.2. Nodules Number, Dry Weight, and Root Length
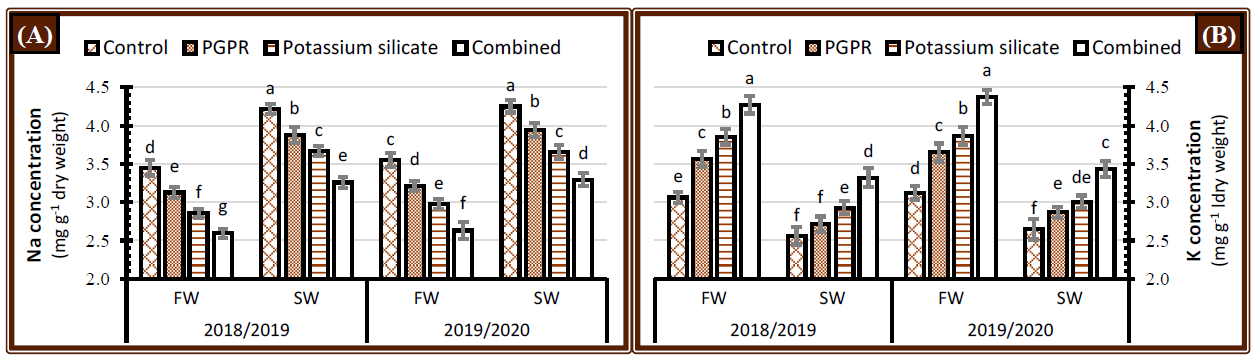
2.3. Inorganic Solutes
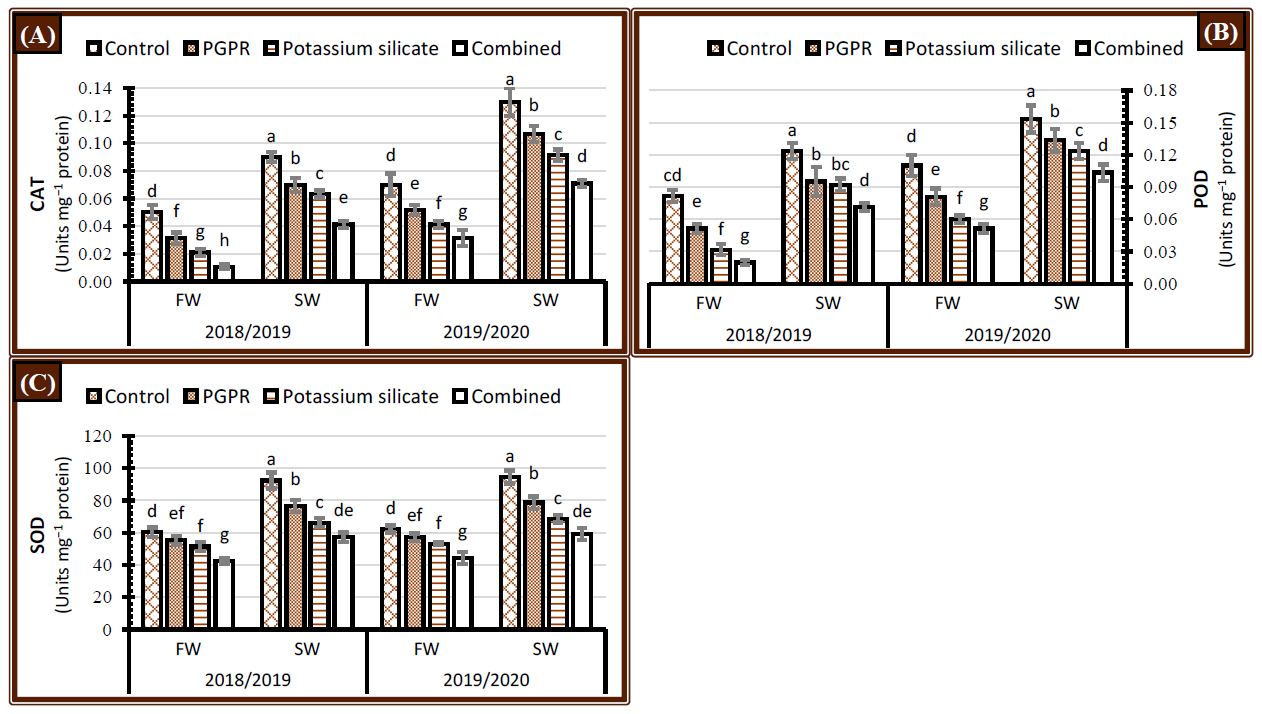
2.4. Activity of Enzymatic Antioxidants
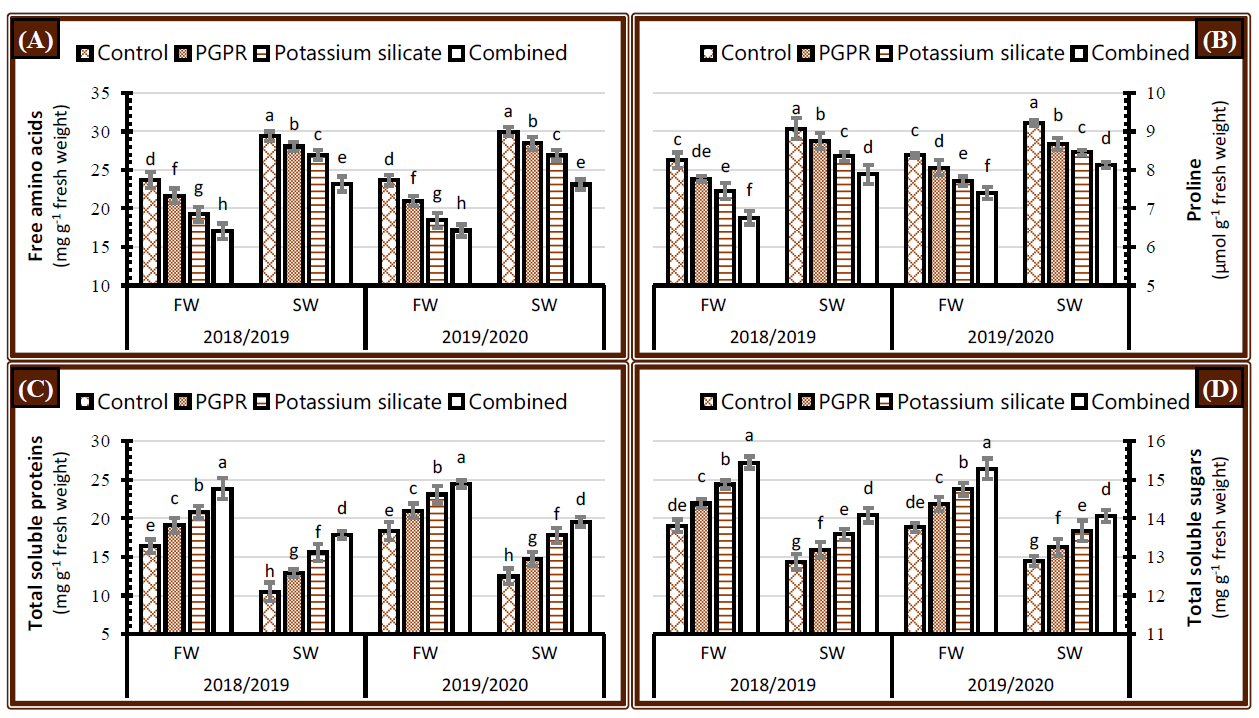
2.5. Non-Enzymatic Antioxidants
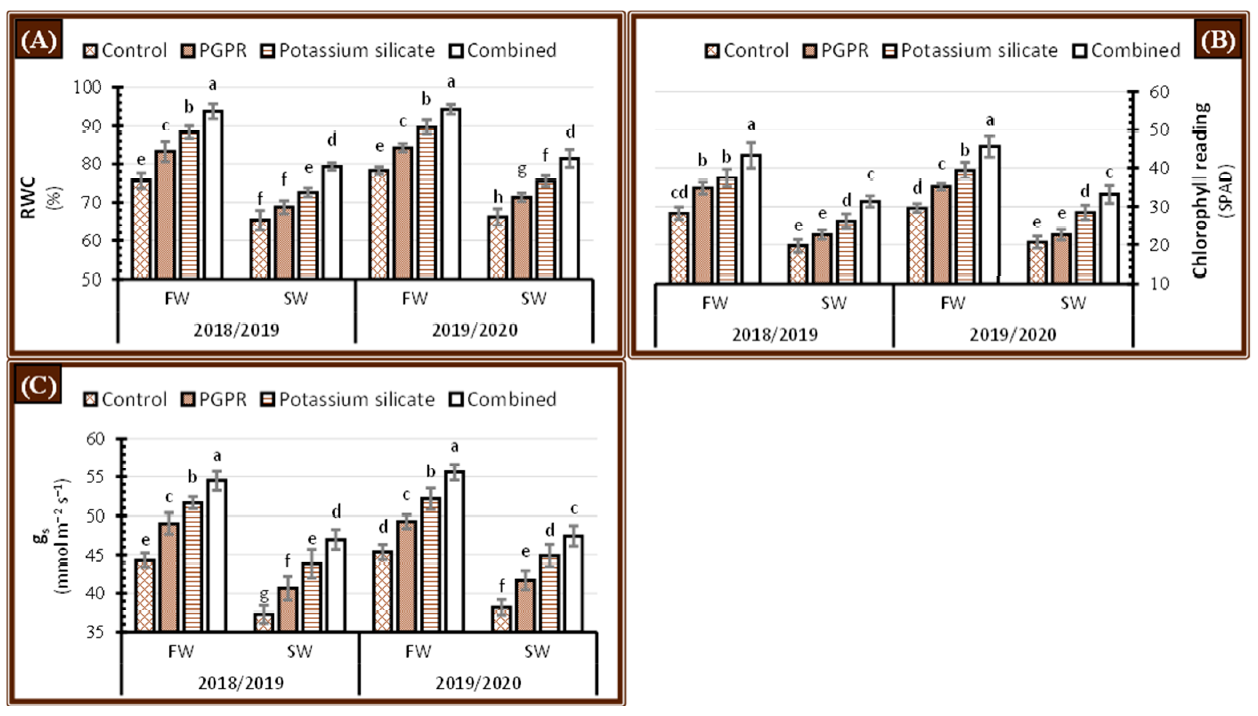
2.6. Physiological Processes
2.7. Yield and Yield-Related Traits
2.8. Grain N, P, and K Uptake
3. Discussion
3.1. Soil Quality Indicators
3.2. Nodulation and Root Length
3.3. Inorganic Solutes
3.4. Activity of Enzymatic Antioxidants
3.5. Non-Enzymatic Antioxidants
3.6. Physiological Processes
3.7. Yield Traits and Nutrients Uptake
4. Materials and Methods
4.1. Plant Material and Experimental Detail
4.2. PGPR and Potassium Silicate
4.3. Measurements
4.3.1. Soil Enzyme Activity
4.3.2. Soil Exchangeable Na Percentage Determination
4.3.3. Nodulation and Roots Assessment
4.3.4. Inorganic Solutes Assessment
4.3.5. Assessment the Activities of Antioxidant Enzymes
4.3.6. Osmo-Protectants and Soluble Protein Determination
4.3.7. Physiological Processes Assessment
4.3.8. Yield Attributes Assessment
4.3.9. Assessment of Nitrogen, Phosphorus and Potassium Uptake
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rawal, V.; Navarro, D.K. The Global Economy of Pulses; Food and Agriculture Organization of the United Nations(FAO): Rome, Italy, 2019. [Google Scholar]
- Etemadi, F.; Barker, A.V.; Hashemi, M.; Zandvakili, O.R.; Park, Y. Nutrient accumulation in faba bean varieties. Comm. Soil Sci. Plant Anal. 2018, 49, 2064–2073. [Google Scholar] [CrossRef]
- Mínguez, M.I.; Rubiales, D. Faba Bean. In Crop Physiology Case Histories for Major Crops; Sadras, V.O., Calderini, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 452–481. [Google Scholar]
- Sahab, S.; Suhani, I.; Srivastava, V.; Chauhan, P.S.; Singh, R.P.; Prasad, V. Potential risk assessment of soil salinity to agroecosystem sustainability: Current status and management strategies. Sci. Total Environ. 2021, 764, 144164. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.; Qureshi, A.S.; Cheraghi, S.A.M. Extent and characterisation of salt-affected soils in Iran and strategies for their amelioration and management. Land Degrad. Dev. 2008, 19, 214–227. [Google Scholar] [CrossRef]
- Pessarakli, M.; Szabolcs, I. Soil Salinity and Sodicity as Particular Plant/Crop Stress Factors. In Handbook of Plant and Crop Stress, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 3–21. [Google Scholar]
- Machado, R.M.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Manchanda, G.; Garg, N. Salinity and its effects on the functional biology of legumes. Acta Physiol. Plant. 2008, 30, 595–618. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.L.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef]
- Shilev, S. Plant-growth-promoting bacteria mitigating soil salinity stress in plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Chen, Y.X.; Zou, L.; Penttinen, P.; Chen, Q.; Li, Q.Q.; Wang, C.Q.; Xu, K.W. Faba bean (Vicia faba L.) nodulating rhizobia in panxi, china, are diverse at species, plant growth promoting ability, and symbiosis related gene levels. Front. Microbiol. 2018, 9, 1338. [Google Scholar] [CrossRef]
- Yadav, J.; Verma, J.P. Effect of seed inoculation with indigenous Rhizobium and plant growth promoting rhizobacteria on nutrients uptake and yields of chickpea (Cicer arietinum L.). Eur. J. Soil Biol. 2014, 63, 70–77. [Google Scholar] [CrossRef]
- Denton, M.D.; Pearce, D.J.; Peoples, M.B. Nitrogen contributions from faba bean (Vicia faba L.) reliant on soil rhizobia or inoculation. Plants Soil 2012, 365, 363–374. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Zafar-ul-Hye, M.; Sajjad, S.; Naveed, M. Comparative effectiveness of Pseudomonas and Serratia sp. containing ACC-deaminase for coinoculation with Rhizobium leguminosarum to improve growth, nodulation, and yield of lentil. Biol. Fertil. Soils 2011, 47, 457–465. [Google Scholar] [CrossRef]
- Flores-Félix, J.D.; Menéndez, E.; Rivera, L.P.; Marcos-García, M.; Martínez-Hidalgo, P.; Mateos, P.F.; Martínez-Molina, E.; Velázquez, M.E.; García-Fraile, P.; Rivas, R. Use of Rhizobium leguminosarumas a potential biofertilizer for Lactuca sativa and Daucus carotacrops. J. Plant Nutr. Soil Sci. 2013, 176, 876–882. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; El-Enany, A.-W.E.; Nafady, N.A.; Khalaf, D.M.; Morsy, F.M. Synergistic interaction of Rhizobium leguminosarum bv. viciae and arbuscular mycorrhizal fungi as a plant growth promoting biofertilizers for faba bean (Vicia faba L.) in alkaline soil. Microbiol. Res. 2014, 169, 49–58. [Google Scholar] [CrossRef]
- Solyman, S.N.; Abdel-Monem, M.; Abou-Taleb, K.; Osman, H.S.; El-Sharkawy, R.M. Production of plant growth regulators by some fungi isolated under salt stress. South Asian J. Res. Microbiol. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Nour El-Din, M.; El-Shanshoury, A.R.; El-Akhdar, I.A. Isolation and evaluation of salt-tolerant microorganisms and their impact in adaptation of faba bean to salinity stress. J. Soil Sci. Agric. Eng. 2011, 2, 977–999. [Google Scholar] [CrossRef]
- Khalil, M.M.N. Effect of mineral fertilizers and biofertilization on some soil properties and faba bean productivity under saline soil conditions. J. Soil Sci. Agric. Eng. 2019, 10, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, M.A.; Kandil, E.E.; El-Dein, A.A.M.Z.; Abou-Donia, M.E.M.; Ali, H.M.; Abdelsalam, N.R. Increase maize productivity and water use efficiency through application of potassium silicate under water stress. Sci. Rep. 2021, 11, 224. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Duarte, H.S.S.; Domiciano, G.P.; Souza, C.A.; Korndörfer, G.H.; Zambolim, L. Foliar application of potassium silicate reduces the intensity of soybean rust. Australas. Plant Pathol. 2009, 38, 366–372. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.i.n.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Yaghubi, K.; Ghaderi, N.; Vafaee, Y.; Javadi, T. Potassium silicate alleviates deleterious effects of salinity on two strawberry cultivars grown under soilless pot culture. Sci. Hortic. 2016, 213, 87–95. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, S.; Al Mahmud, J.; Hossen, S.; Masud, A.A.C.; Moumita, M.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Abdel Latef, A.A.; Abd_Allah, E.F.; Hashem, A.; Sarwat, M.; Anjum, N.A.; Gucel, S. Calcium and potassium supplementation enhanced growth, osmolyte secondary metabolite production, and enzymatic antioxidant machinery in cadmium-exposed chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Osman, H.S.; Gowayed, S.M.; Okasha, S.A.; Omara, A.E.-D.; Sami, R.; Abd El-Monem, A.M.; Abd El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.-G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) supplementation alleviates NaCl toxicity in mung bean [Vigna radiata (L.) Wilczek] through the modifications of physio-biochemical attributes and key antioxidant enzymes. J. Plant Growth Regul. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Hikal, M.S.; Osman, H.S. Ameliorating the deleterious effects of saline water on the antioxidants defense system and yield of eggplant using foliar application of zinc sulphate. Ann. Agric. Sci. 2019, 64, 244–251. [Google Scholar] [CrossRef]
- Frankenberger, W.T., Jr.; Bingham, F.T. Influence of salinity on soil enzyme activities. Soil Sci. Soc. Am. J. 1982, 46, 1173–1177. [Google Scholar] [CrossRef]
- Youssif, N.E.E.; Osman, H.S.M.; Salama, Y.A.M.; Zaghlool, S.A.M. Effect of rice straw and applications of potassium silicate, potassium humate and seaweed extract on growth and some macronutrients of sweet pepper plants under irrigation deficit. Arab Univ. J. Agric. Sci. 2018, 26, 755–773. [Google Scholar] [CrossRef]
- Samaddar, S.; Truu, J.; Chatterjee, P.; Truu, M.; Kim, K.; Kim, S.; Seshadri, S.; Sa, T. Long-term silicate fertilization increases the abundance of Actinobacterial population in paddy soils. Biol. Fertil. Soils 2019, 55, 109–120. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.M.S.; Ali, O.A.M.; Hafez, E.M.; ElShamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.e.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2021, 277, 111388. [Google Scholar] [CrossRef]
- Bechtaoui, N.; Raklami, A.; Tahiri, A.-I.; Benidire, L.; El Alaoui, A.; Meddich, A.; Göttfert, M.; Oufdou, K. Characterization of plant growth promoting rhizobacteria and their benefits on growth and phosphate nutrition of faba bean and wheat. Biol. Open 2019, 8, bio043968. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.A.; Behr, J.H.; Erban, A.; Kopka, J.; Zörb, C. Ion-dependent metabolic responses of Vicia faba L. to salt stress. Plant Cell Environ. 2018, 42, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant 2020, 1–16. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Mohammed, A.F. Effectiveness of exopolysaccharides and biofilm forming plant growth promoting rhizobacteria on salinity tolerance of faba bean (Vicia faba L.). Afr. J. Microbiol. Res. 2018, 12, 399–404. [Google Scholar]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative effects of rice-straw biochar and silicon on oil and seed quality, yield and physiological traits of Helianthus annuus L. Grown under water deficit stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.; Ahsen, S.; Ali, M.A.; Hussain, M.B.; Hussain, S.B.; Rasheed, M.K.; Butt, B.; Irshad, I.; Danish, S. Rhizobacteria and silicon synergy modulates the growth, nutrition and yield of mungbean under saline soil. Pak. J. Bot. 2020, 52, 9–15. [Google Scholar] [CrossRef]
- Rawat, J.; Sanwal, P.; Saxena, J. Potassium and Its Role in Sustainable Agriculture. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 235–253. [Google Scholar]
- Etesami, H.; Adl, S.M. Can interaction between silicon and non–rhizobial bacteria help in improving nodulation and nitrogen fixation in salinity–stressed legumes? A review. Rhizosphere 2020, 15, 100229. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gharib, H.S. Effect of exogenous application of ascorbic acid on physiological and biochemical characteristics of wheat under water stress. Int. J. Plant Prod. 2016, 10, 579–596. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, H.I.; Sallam, A.M.; Shaban, K.A. Impact of irrigation rates and potassium silicate fertilizer on seed production and quality of Fahl Egyptian clover and soil properties under saline conditions. Am. Eurasian J. Agric. Environ. Sci. 2015, 15, 1245–1255. [Google Scholar]
- Osman, H.S. Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.S.; Salim, B.B.M. Enhancing Antioxidants Defense System of Snap Bean under NaCl Salinity Using Foliar Application of Salicylic Acid, Spermidine and Glycine Betaine. Am. Eurasian J. Agric. Environ. Sci. 2016, 16, 1200–1210. [Google Scholar]
- Qados, A.M.A. Mechanism of nanosilicon-mediated alleviation of salinity stress in faba bean (Vicia faba L.) plants. Am. J. Exp. Agric. 2015, 7, 78–95. [Google Scholar] [CrossRef]
- Chiappero, J.; Cappellari, L.d.R.; Sosa Alderete, L.G.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crop. Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Osman, H.S.; Salim, B.B.M. Influence of exogenous application of some phytoprotectants on growth, yield and pod quality of snap bean under NaCl salinity. Ann. Agric. Sci. 2016, 61, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.S.; Salim, B.B. Improving yield and quality of kohlrabi stems growing under NaCl salinity using foliar application of urea and seaweed extract. J. Hort. Sci. Ornament. Plants 2016, 8, 149–160. [Google Scholar]
- Hafez, E.M.; Kheir, A.M.S.; Badawy, S.A.; Rashwan, E.; Farig, M.; Osman, H.S. Differences in physiological and biochemical attributes of wheat in response to single and combined salicylic acid and biochar subjected to limited water irrigation in saline sodic soil. Plants 2020, 9, 1346. [Google Scholar] [CrossRef]
- Moussa, H.R.; Hassan, M.A.E.-F. Growth enhancers to mitigate salinity stress in Vicia Faba. Int. J. Veg. Sci. 2016, 22, 243–250. [Google Scholar] [CrossRef]
- Hafez, E. Influence of salicylic acid on ion distribution, enzymatic activity and some agromorphological characteristics of wheat under salt-affected soil. Egypt J. Agron. 2016, 38, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Hellal, F.; Abdelhameid, M.; Abo-Basha, D.M.; Zewainy, R. Alleviation of the adverse effects of soil salinity stress by foliar application of silicon on faba bean (Vica faba L.). J. Appl. Sci. Res. 2012, 8, 4428–4433. [Google Scholar]
- Calero Hurtado, A.; Aparecida Chiconato, D.; de Mello Prado, R.; da Silveira Sousa, G., Jr.; Felisberto, G. Silicon attenuates sodium toxicity by improving nutritional efficiency in sorghum and sunflower plants. Plant Physiol. Biochem. 2019, 142, 224–233. [Google Scholar] [CrossRef]
- Cochrane, T.T.; Cochrane, T.A. The vital role of potassium in the osmotic mechanism of stomata aperture modulation and its link with potassium deficiency. Plant Signal. Behav. 2009, 4, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Melesse, T.; Caesar, K. Stomatal and non-stomatal effects of salinity on photosynthesis in faba beans (Vicia faba L.). J. Agron. Crop Sci. 1992, 168, 345–353. [Google Scholar] [CrossRef]
- Hussein, M.; Embiale, A.; Husen, A.; Aref, I.M.; Iqbal, M. Salinity-induced modulation of plant growth and photosynthetic parameters in faba bean (Vicia faba) cultivars. Pak. J. Bot. 2017, 49, 867–877. [Google Scholar]
- Metwali, E.M.; Abdelmoneim, T.S.; Bakheit, M.A.; Kadasa, N.M. Alleviation of salinity stress in faba bean (Vicia faba L.) plants by inoculation with plant growth promoting rhizobacteria (PGPR). Plant Omics 2015, 8, 449. [Google Scholar]
- Salem, H.; Abo-Setta, Y.; Aiad, M.; Hussein, H.-A.; El-Awady, R. Effect of potassium humate and potassium silicate on growth and productivity of wheat plants grown under saline conditions. J. Soil Sci. Agric. Eng. 2017, 8, 577–582. [Google Scholar] [CrossRef]
- Ali, N.; Réthoré, E.; Yvin, J.-C.; Hosseini, S.A. The regulatory role of silicon in mitigating plant nutritional stresses. Plants 2020, 9, 1779. [Google Scholar] [CrossRef]
- Hafez, E.; Farig, M. Efficacy of salicylic acid as a cofactor for ameliorating effects of water stress and enhancing wheat yield and water use efficiency in saline soil. Inter. J. Plant Prod. 2019, 13, 163–176. [Google Scholar] [CrossRef]
- Sivanesan, I.; Jeong, B.R. Silicon promotes adventitious shoot regeneration and enhances salinity tolerance of Ajuga multiflora bunge by altering activity of antioxidant enzyme. Sci. World J. 2014, 2014, 521703. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 2040. [Google Scholar]
- Kandeler, E. Methods in Soil Chemistry: Ammonium. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 406–408. [Google Scholar]
- Mersi, W.V. Dehydrogenase Activity with the Substrate INT. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 243–245. [Google Scholar]
- Seilsepour, M.; Rashidi, M.; Khabbaz, B.G. Prediction of soil exchangeable sodium percentage based on soil sodium adsorption ratio. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 1–4. [Google Scholar]
- Aebi, H. Catalase In Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme assay, quantitative determination of peroxidase in sweet corn. J. Agric. Food Chem. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Phenol sulphuric acid method for total carbohydrate. Anal. Chem. 1956, 26, 350. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Swamy, P.M. Laboratory Manual on Biotechnology; Rastogi Publications: Uttar Preadesh, India, 2008; pp. 80–82. [Google Scholar]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosyn. Res. 2010, 107, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pods No. Plant−1 | Seeds No. Pod−1 | 100-Seed Weight (g) | Seed Yield (t ha−1) | ||
|---|---|---|---|---|---|---|
| 2018/2019 | FW ‡ | Control | 15.5 ± 0.9 e | 4.5 ± 0.36 cd | 54.3 ± 2.1 e | 2.2 ± 0.10 d |
| PGPR | 19.7 ± 0.8 c | 5.7 ± 0.42 b | 65.5 ± 2.4 c | 2.8 ± 0.04 bc | ||
| K silicate † | 22.8 ± 0.8 b | 6.1 ± 0.56 b | 76.7 ± 2.7 b | 3.1 ± 0.17 b | ||
| PGPR + K silicate | 28.3 ± 1.2 a | 6.7 ± 0.15 a | 83.7 ± 2.6 a | 3.6 ± 0.21 a | ||
| SW § | Control | 10.7 ± 1.2 g | 3.2 ± 0.20 e | 32.3 ± 1.3 h | 1.5 ± 0.18 f | |
| PGPR | 12.9 ± 1.7 f | 4.1 ± 0.17 d | 40.8 ± 1.4 g | 1.7 ± 0.22 ef | ||
| K silicate † | 14.4 ± 1.2 ef | 4.3 ± 0.14 d | 47.7 ± 1.4 f | 2.1 ± 0.16 de | ||
| PGPR + K silicate | 17.6 ± 1.1 d | 5.0 ± 0.39 c | 57.3 ± 1.8 d | 2.7 ± 0.25 c | ||
| 2019/2020 | FW ‡ | Control | 17.7 ± 1.3 de | 4.9 ± 0.20 de | 55.8 ± 2.7 d | 2.4 ± 0.12 d |
| PGPR | 20.2 ± 1.6 c | 5.7 ± 0.21 c | 66.4 ± 5.9 c | 3.0 ± 0.15 bc | ||
| K silicate † | 24.9 ± 2.0 b | 6.2 ± 0.19 b | 79.5 ± 1.4 b | 3.2 ± 0.23 b | ||
| PGPR + K silicate | 29.7 ± 1.6 a | 6.8 ± 0.27 a | 85.5 ± 2.9 a | 3.9 ± 0.17 a | ||
| SW § | Control | 11.4 ± 1.4 f | 3.3 ± 0.3 g | 33.6 ± 2.7 g | 1.6 ± 0.14 f | |
| PGPR | 13.5 ± 1.8 f | 4.4 ± 0.19 f | 44.2 ± 1.6 f | 2.0 ± 0.14 e | ||
| K silicate † | 15.7 ± 1.1 e | 4.6 ± 0.22 ef | 49.6 ± 1.8 e | 2.3 ± 0.13 de | ||
| PGPR + K silicate | 18.8 ± 1.0 cd | 5.2 ± 0.20 d | 59.3 ± 2.0 d | 2.7 ± 0.16 c | ||
| Water treatments | *** | ** | ** | ** | ||
| Soil and foliar treatments | *** | *** | *** | *** | ||
| Interaction | * | ns | ** | ns | ||
| Treatments | N Uptake (kg ha−1) | P Uptake (kg ha−1) | K Uptake (kg ha−1) | ||||
|---|---|---|---|---|---|---|---|
| 2018/2019 | 2019/2020 | 2018/2019 | 2019/2020 | 2018/2019 | 2019/2020 | ||
| FW ‡ | Control | 108.2 ± 2.3 e | 112.4 ± 2.3 e | 16.4± 0.5 e | 16.9 ± 0.4 e | 17.7 ± 0.6 e | 19.8 ± 0.4 f |
| PGPR | 119.8 ± 2.5 c | 123.1 ± 2.5 c | 17.8 ± 0.7 c | 18.0 ± 0.6 c | 24.1 ± 0.5 c | 26.1 ± 0.6 c | |
| K silicate † | 126.6 ± 2.9 b | 129.7 ± 2.8 b | 18.4 ± 0.8 b | 18.9 ± 0.7 b | 29.2 ± 0.7 b | 30.3 ± 0.6 d | |
| PGPR + K silicate | 138.1 ± 3.2 a | 142.4 ± 2.9 a | 20.2 ± 0.9 a | 21.3 ± 0.9 a | 36.3 ± 0.8 a | 39.6 ± 0.7 a | |
| SW § | Control | 86.4 ± 2.4 h | 91.4 ± 2.1 h | 14.1 ± 0.3 h | 15.6 ± 0.5 h | 6.6 ± 0.5 h | 8.6 ± 0.5 h |
| PGPR | 95.8 ± 2.6 g | 100.3 ± 2.3 g | 14.9 ± 0.5 g | 16.2 ± 0.6 g | 9.6 ± 0.7 g | 11.8 ± 0.6 g | |
| K silicate † | 104.0 ± 2.8 f | 108.4 ± 2.7 f | 15.6 ± 0.7 f | 16.9 ± 0.7 f | 13.7 ± 0.8 f | 16.9 ± 0.8 e | |
| PGPR + K silicate | 112.4 ± 2.9 d | 119.4 ±3.2 d | 16.9 ± 0.9 d | 17.5 ± 0.8 d | 19.8 ± 0.9 d | 22.0 ± 0.9 d | |
| Water treatments | *** | ** | *** | ||||
| Soil and foliar treatments | *** | *** | *** | ||||
| Interaction | ns | ns | * | ||||
| Character | Fresh Water | Saline Water | ||
|---|---|---|---|---|
| 2018/2019 | 2019/2020 | 2018/2019 | 2019/2020 | |
| pH | 7.28 | 7.34 | 8.36 | 8.39 |
| EC (dS m−1) | 0.59 | 0.56 | 3.43 | 3.57 |
| SAR | 1.46 | 1.42 | 7.69 | 7.78 |
| Na+ (meq L−1) | 1.88 | 1.97 | 16.44 | 16.31 |
| Cl− (meq L−1) | 3.40 | 3.28 | 11.52 | 11.70 |
| SO42− (meq L−1) | 0.12 | 0.13 | 7.83 | 8.13 |
| NH4+ (meq L−1) | 1.74 | 1.80 | 2.02 | 2.13 |
| Soil Traits | 2018/2019 | 2019/2020 |
|---|---|---|
| clay % | 49.40 | 54.00 |
| Sand % | 15.53 | 13.00 |
| Silt % | 35.07 | 33.00 |
| Soil texture | Clayey | Clayey |
| pH (1:2.5 water suspension) | 8.24 | 8.02 |
| EC (dS m−1) | 5.52 | 5.24 |
| Organic matter | 1.19 | 1.29 |
| Available P mg kg−1 | 9.44 | 8.54 |
| Available N mg kg−1 | 9.40 | 8.60 |
| Available K mg kg−1 | 309 | 294 |
| Cations (meq L−1) | ||
| Ca2+ | 5.18 | 4.91 |
| Mg2+ | 1.34 | 1.22 |
| Na+ | 16.00 | 19.00 |
| K+ | 0.41 | 0.39 |
| Anions (meq L −1) | ||
| HCO3− | 4.13 | 4.10 |
| Cl− | 17.10 | 20.00 |
| SO42− | 1.70 | 1.42 |
| CO3− | 0.00 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafez, E.M.; Osman, H.S.; El-Razek, U.A.A.; Elbagory, M.; Omara, A.E.-D.; Eid, M.A.; Gowayed, S.M. Foliar-Applied Potassium Silicate Coupled with Plant Growth-Promoting Rhizobacteria Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia faba L.) Irrigated with Saline Water in Salt-Affected Soil. Plants 2021, 10, 894. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050894
Hafez EM, Osman HS, El-Razek UAA, Elbagory M, Omara AE-D, Eid MA, Gowayed SM. Foliar-Applied Potassium Silicate Coupled with Plant Growth-Promoting Rhizobacteria Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia faba L.) Irrigated with Saline Water in Salt-Affected Soil. Plants. 2021; 10(5):894. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050894
Chicago/Turabian StyleHafez, Emad M., Hany S. Osman, Usama A. Abd El-Razek, Mohssen Elbagory, Alaa El-Dein Omara, Mohamed A. Eid, and Salah M. Gowayed. 2021. "Foliar-Applied Potassium Silicate Coupled with Plant Growth-Promoting Rhizobacteria Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia faba L.) Irrigated with Saline Water in Salt-Affected Soil" Plants 10, no. 5: 894. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050894