
Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Materials
2.2. DNA Isolation
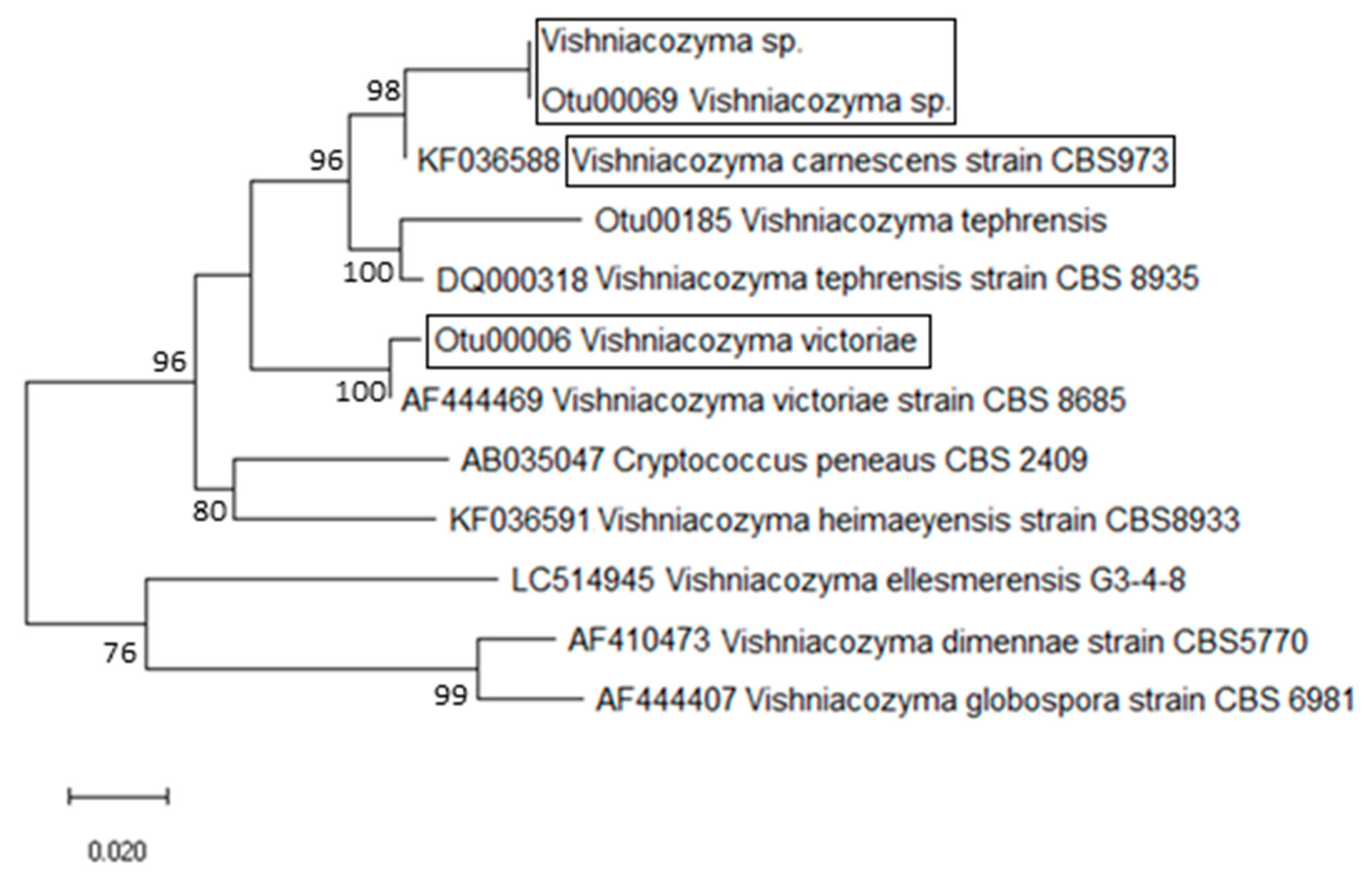
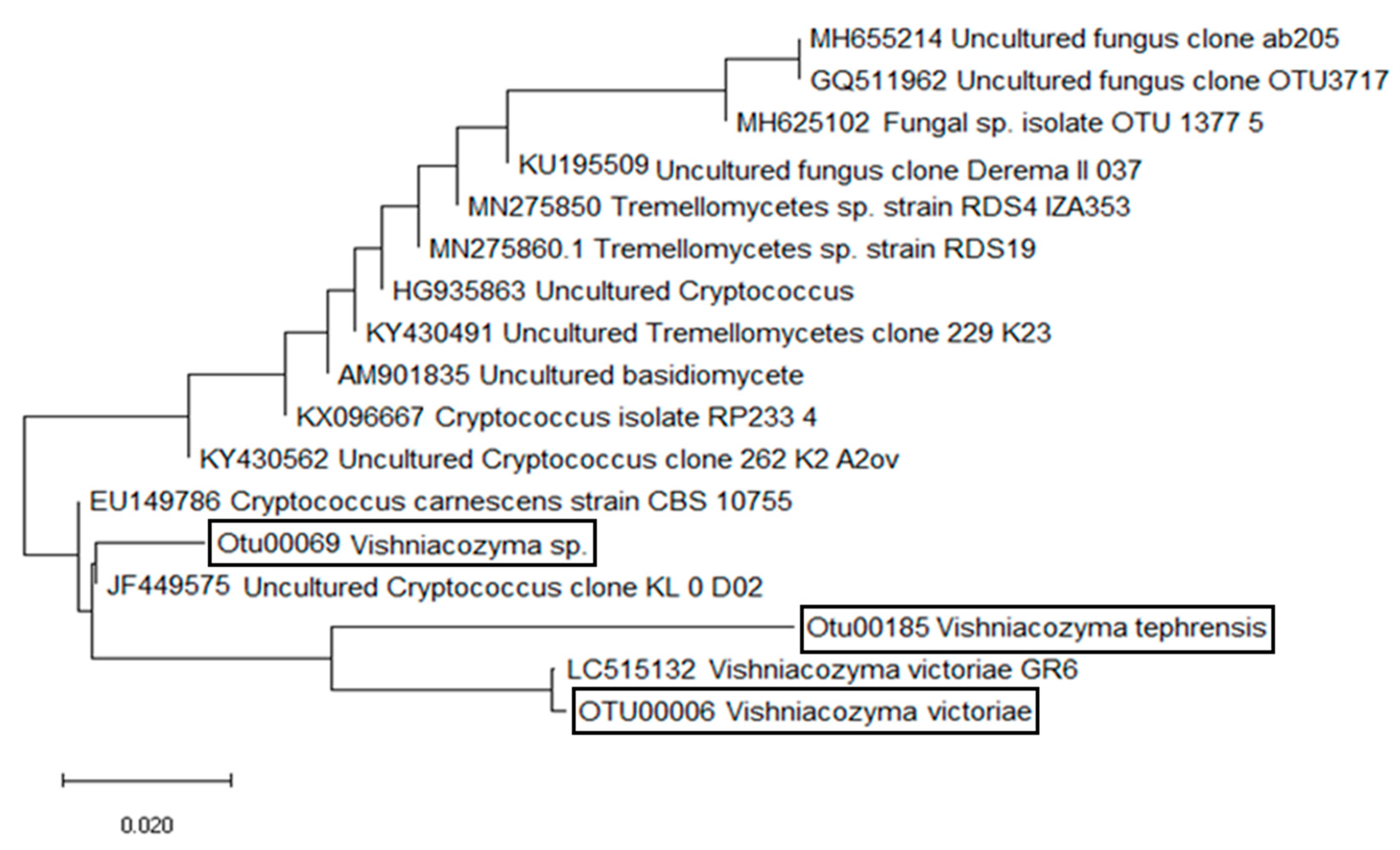
2.3. Phylogenetic Analysis
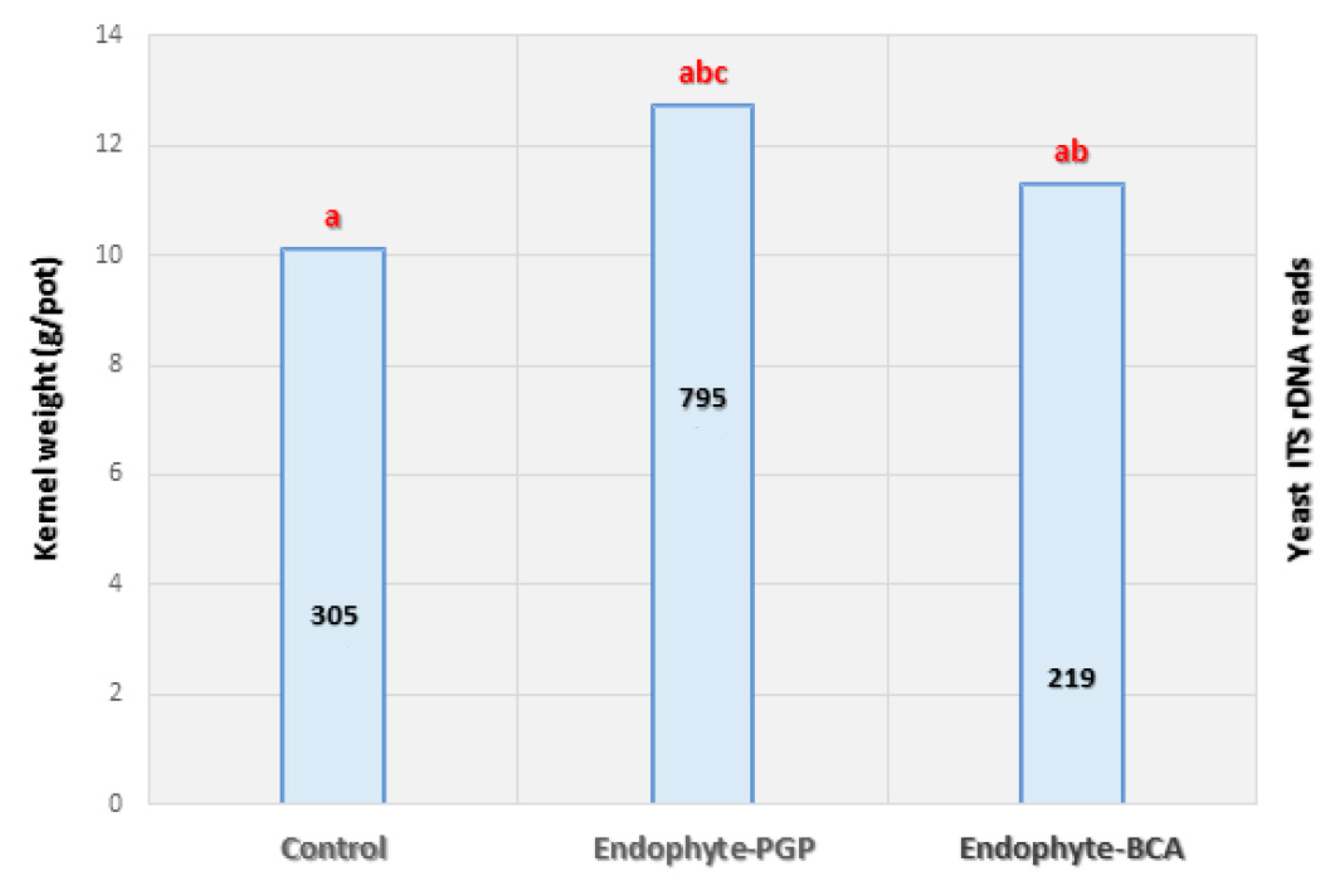
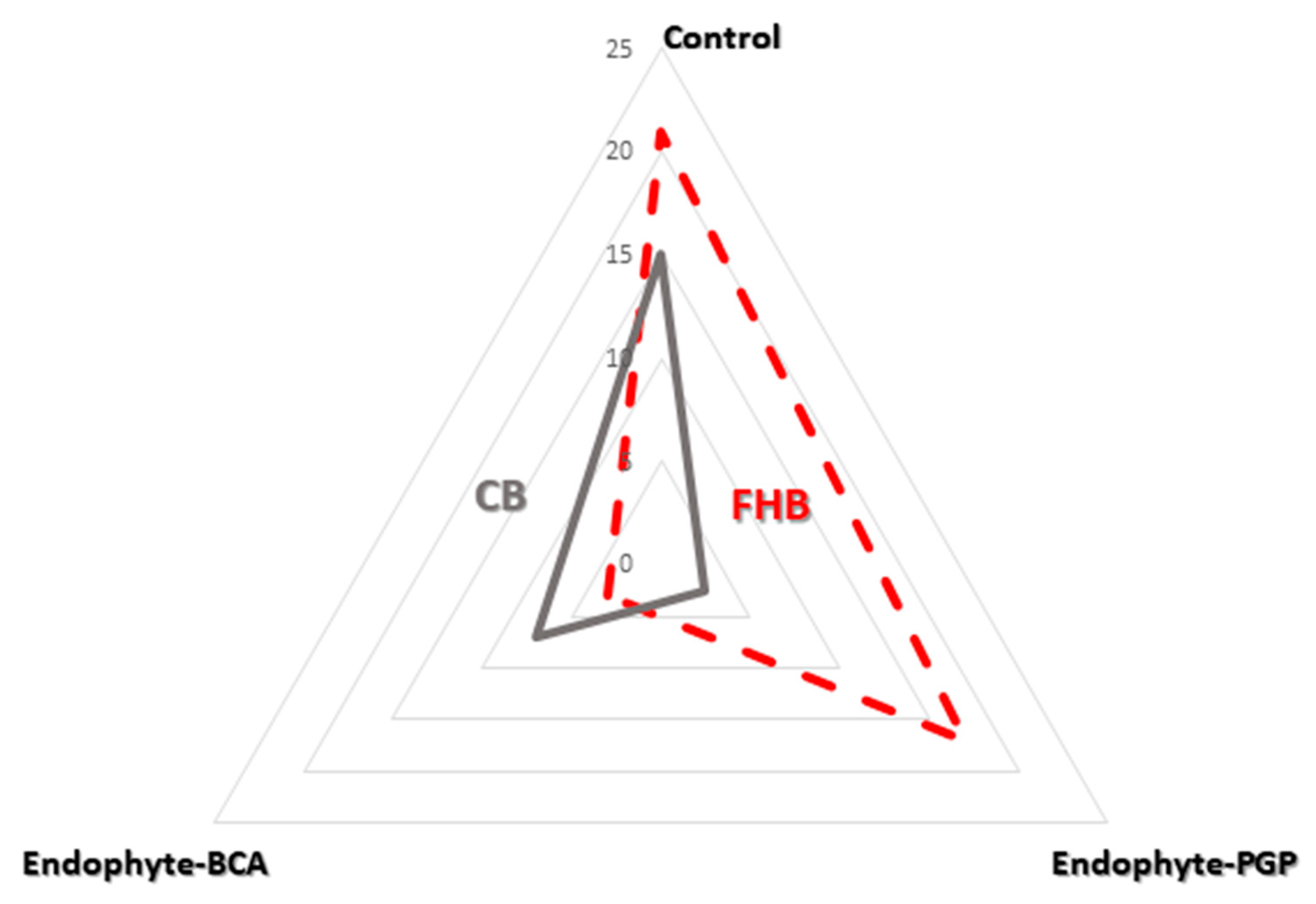
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier: London, UK, 2014. [Google Scholar]
- Vujanovic, V.; Islam, M.N.; Daida, P. Transgenerational role of seed mycobiome—An endosymbiotic fungal composition as a prerequisite to stress resilience and adaptive phenotypes in Triticum. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Vujanovic, V.; Vujanovic, J. Mycovitality and mycoheterotrophy: Where lies dormancy in terrestrial orchid and plants with minute seeds. Symbiosis 2007, 44, 93–99. [Google Scholar]
- Vujanovic, V.; Yuan, X.; Daida, P.; Milunovic, B.; Germida, J. Manipulation of cold stratification and endophytic effects on expression patterns of RSG and KAO genes in coleorhiza of wheat seeds. Plant Growth Regul. 2015, 79, 219–227. [Google Scholar] [CrossRef]
- Vujanovic, V.; Kim, S.H.; Lahlali, R.; Karunakaran, C. Spectroscopy and SEM imaging reveal endosymbiont-dependent components changes in germinating kernel through direct and indirect coleorhiza-fungus interactions under stress. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Kumari, V.; Vujanovic, V. Transgenerational benefits of endophytes on resilience and antioxidant genes expressions in pea (Pisum sativum L.) under osmotic stress. Acta Physiol. Plant. 2020, 42, 1–11. [Google Scholar] [CrossRef]
- Vujanovic, V.; Germida, J. Seed endosymbiosis: A vital relationship in providing prenatal care to plants. Can. J. Plant Sci. 2017, 97, 972–981. [Google Scholar] [CrossRef] [Green Version]
- Yurkov, A.M.; Kurtzman, C.P. Three new species of Tremellomycetes isolated from maize and northern wild rice. FEMS Yeast Res. 2019, 19, foz004. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Barra, P.J.; Jorquera, M.A.; Viscardi, S.; Fernandez, C.; Paz, C.; Mora, M.D.L.L.; Bol, R. Occurrence of Soil Fungi in Antarctic Pristine Environments. Front. Bioeng. Biotechnol. 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Vishniacozyma ellesmerensis sp. nov., a psychrophilic yeast isolated from a retreating glacier in the Canadian High Arctic. Int. J. Syst. Evol. Microbiol. 2019, 69, 696–700. [Google Scholar] [CrossRef]
- Rojas, E.C.; Sapkota, R.; Jensen, B.; Jørgensen, H.J.L.; Henriksson, T.; Jørgensen, L.N.; Nicolaisen, M.; Collinge, D.B. Fusarium Head Blight Modifies Fungal Endophytic Communities During Infection of Wheat Spikes. Microb. Ecol. 2020, 79, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Sibanda, E.P.; Mabandla, M.; Chisango, T.; Nhidza, A.F.; Mduluza, T. Endophytic Fungi Isolated from the Medicinal Plants Kigelia africana and Warburgia salutaris. Curr. Biotechnol. 2018, 7, 323–328. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Teixeira, D.R.; Vieira, R.; Lírio, J.M.; Felizardo, J.P.; Abuchacra, R.C.; Cardoso, R.P.; Zani, C.L.; Alves, T.M.; Junior, P.A.; et al. Diversity and bioprospecting of cultivable fungal assemblages in sediments of lakes in the Antarctic Peninsula. Fungal Biol. 2020, 124, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.; Gorte, O.; Zhou, X.; Neumann, A.; Ochsenreither, K. In silico Proteomic Analysis Provides Insights into Phylogenomics and Plant Biomass Deconstruction Potentials of the Tremelalles. Front. Bioeng. Biotechnol. 2020, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Turchetti, B.; Yurkov, A. Extremophilic yeasts: The toughest yeasts around? Yeast 2018, 35, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, Á.; Inácio, J. Phylloplane yeasts. In Biodiversity and Ecophysiology of Yeasts; Péter, G., Rosa, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Kemler, M.; Witfeld, F.; Begerow, D.; Yurkov, A. Phylloplane Yeasts in Temperate Climates. In Yeasts in Natural Ecosystems: Diversity; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Streletskii, R.A.; Kachalkin, A.V.; Glushakova, A.M.; Yurkov, A.M.; Demin, V.V. Yeasts producing zeatin. PeerJ 2019, 7, e6474. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2011; p. 2354. [Google Scholar]
- Zugmaier, W.; Bauer, R.; Oberwinkler, F. Mycoparasitism of some Tremella species. Mycologia 1994, 86, 49–56. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.-Z.; Bai, F.-Y. Four new species of Tremella (Tremellales, Basidiomycota) based on morphology and DNA sequence data. MycoKeys 2019, 47, 75–95. [Google Scholar] [CrossRef]
- Weiß, M.; Bauer, R.; Sampaio, J.P.; Oberwinkler, F. Tremellomycetes and related groups. systematics and evolution. In The Mycota VII Part A, 2nd ed.; McLaughlin, D.J., Spatafora, J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Liu, X.Z.; Wang, Q.-M.; Theelen, B.; Groenewald, M.; Bai, F.Y.; Boekhout, T. Phylogeny of tremellomycetous yeasts and related dimorphic and filamentous basidiomycetes recon-structed from multiple gene sequence analyses. Stud. Mycol. 2015, 81, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.-H.; Yuan, F.-X.; Groenewald, M.; Bensch, K.; Yurkov, A.; Li, K.; Han, P.-J.; Guo, L.-D.; Aime, M.; Sampaio, J.; et al. Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: Proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud. Mycol. 2020, 96, 17–140. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P. Index Fungorum Database; CABI: Kew, UK, 2020; Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 5 January 2020).
- Liu, X.-Z.; Wang, Q.-M.; Göker, M.; Groenewald, M.; Kachalkin, A.; Lumbsch, H.; Millanes, A.; Wedin, M.; Yurkov, A.; Boekhout, T.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [Green Version]
- Fell, J.W. Collection and identification of marine yeasts. In Methods in Microbiology; John, H.P., Ed.; Academic Press: Burlington, VT, USA, 2001; Volume 30, pp. 347–356. [Google Scholar]
- Yurkov, A.M. Yeasts of the soil—Obscure but precious. Yeast 2018, 35, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Vujanovic, V. Relationship between mycoparasites lifestyles and biocontrol behaviors against Fusarium spp. and mycotoxins production. Appl. Microbiol. Biotechnol. 2016, 100, 5257–5272. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Goh, Y.K. Sphaerodes mycoparasitica sp. nov., a new biotrophic mycoparasite on Fusarium avenaceum, F. graminearum and F. oxysporum. Mycol. Res. 2009, 113, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Goh, Y.K. qPCR quantification of Sphaerodes mycoparasitica biotrophic mycoparasite interaction with Fusarium graminearum: In vitro and in planta assays. Arch. Microbiol. 2012, 194, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Paul, P.A.; El-Allaf, S.M.; Lipps, P.E.; Madden, L.V. Relationships between incidence and severity of Fusarium head blight on winter wheat in Ohio. Phytopathology 2005, 95, 1049–1060. [Google Scholar] [CrossRef] [Green Version]
- Riccioni, L.; Inman, A.; Magnus, H.A.; Valvassori, M.; Porta-Puglia, A.; Conca, G.; Di Giambattista, G.; Hughes, K.; Coates, M.; Bowyer, R.; et al. Susceptibility of European bread and durum wheat cultivars to Tilletia indica. Plant Pathol. 2008, 57, 612–622. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggia, F.; Dalanaj, N.; Prodi, A.; Nipoti, P.; Pisi, A.; Biavati, B.; Di Gioia, D. Microbial inoculants for the biocontrol of Fusarium spp. in durum wheat. BMC Microbiol. 2015, 15, 242. [Google Scholar] [CrossRef]
- Rzhetsky, A.; Nei, M. A simple method for estimating and testing minimum evolution trees. Mol. Biol. Evol. 1992, 9, 945–967. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Eberlein, C.; Hénault, M.; Fijarczyk, A.; Charron, G.; Bouvier, M.; Kohn, L.M.; Anderson, J.B.; Landry, C.R. Hybridization is a recurrent evolutionary stimulus in wild yeast speciation. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kulakovskaya, T.V.; Golubev, W.I.; Tomashevskaya, M.A.; Shashkov, A.S.; Grachev, A.A.; Chizhov, A.S.; Nifantiev, N.E. Production of Antifungal Cellobiose Lipids by Trichosporon porosum. Mycopathologia 2010, 169, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, A.M.; Golubev, W.I. Phylogenetic study of Cryptococcus laurentii mycocinogenic strains. Mycol. Prog. 2013, 12, 777–782. [Google Scholar] [CrossRef]
- Prior, R.; Feige, A.; Begerow, D. Antagonistic activity of the phyllosphere fungal community. Sydowia 2017, 69, 183–198. [Google Scholar]
- Gaudet, D.A.; Fuentes-Davila, G.; Burnett, P.A.; De Pauw, R.M. Reactions of western Canadian spring wheat and triticale varieties to Tilletia indica, the causal agent of Karnal bunt. Can. J. Plant Sci. 2001, 81, 503–508. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Theis, K.R. Host biology in light of the microbiome: Ten principles of holobionts and hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [Green Version]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungalinteractions: Hyphens between agricultural, clinical, environmental and food microbiologists. Microbiol. Mol Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [Green Version]
- Morella, N.M.; Weng, F.C.-H.; Joubert, P.M.; Metcalf, C.J.E.; Lindow, S.; Koskella, B. Successive passaging of a plant-associated microbiome reveals robust habitat and host genotype-dependent selection. Proc. Natl. Acad. Sci. USA 2019, 117, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.P.; Wenger, S.; Maurhofer, M.; Defago, G.; Duffy, B. Signaling between bacterial and fungal biocontrol agents in a strain mixture. FEMS Microbiol. Ecol. 2004, 48, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Lahlali, R.; Karunakaran, C.; Vujanovic, V. Specific Mycoparasite-Fusarium Graminearum Molecular Signatures in Germinating Seeds Disabled Fusarium Head Blight Pathogen’s Infection. Int. J. Mol. Sci. 2021, 22, 2461. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment * | FHB | CB | ||||
|---|---|---|---|---|---|---|
| DI (%) | DS(%) | FHB Index | DI (%) | DS (%) | CB Index | |
| Control | 21 b | 13.8 b | 2.9 b | 15 b | 20.6 b | 3.1 |
| Endo-PGP | 17 ab | 7.1 ab | 1.2 ab | 2.5 a | 0.6 a | 0.4 a |
| Endo-BCA | 3.0 ab | 3.3 ab | 0.1 a | 7.0 ab | 10 ab | 0.7 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujanovic, V. Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota. Plants 2021, 10, 905. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050905
Vujanovic V. Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota. Plants. 2021; 10(5):905. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050905
Chicago/Turabian StyleVujanovic, Vladimir. 2021. "Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota" Plants 10, no. 5: 905. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050905