
The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection

 , and
, and 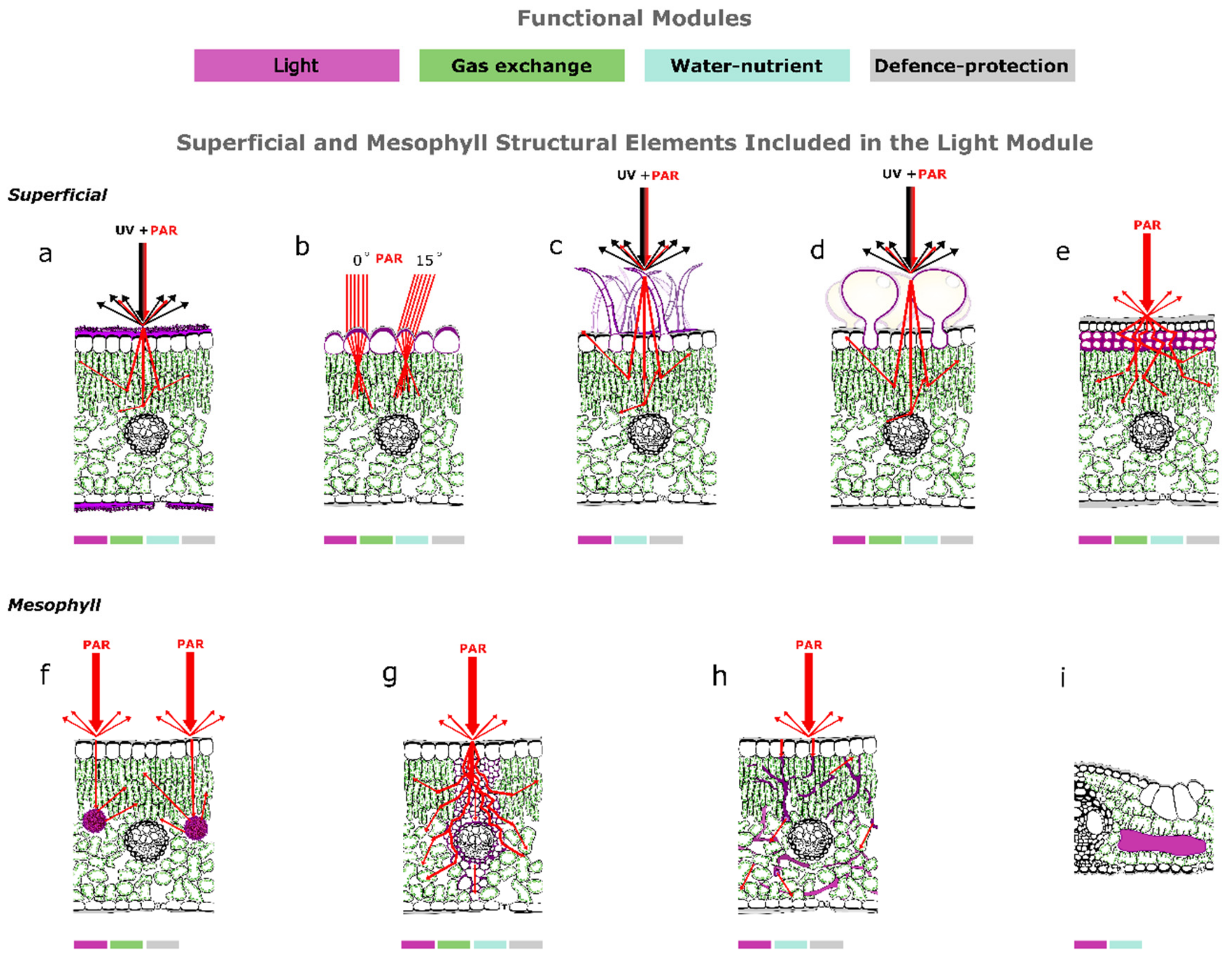{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Superficial Structural Elements and Light Interception
2.1. The Epidermis Is a Selective Optical Filter
2.2. The Epidermis Is a Reflector
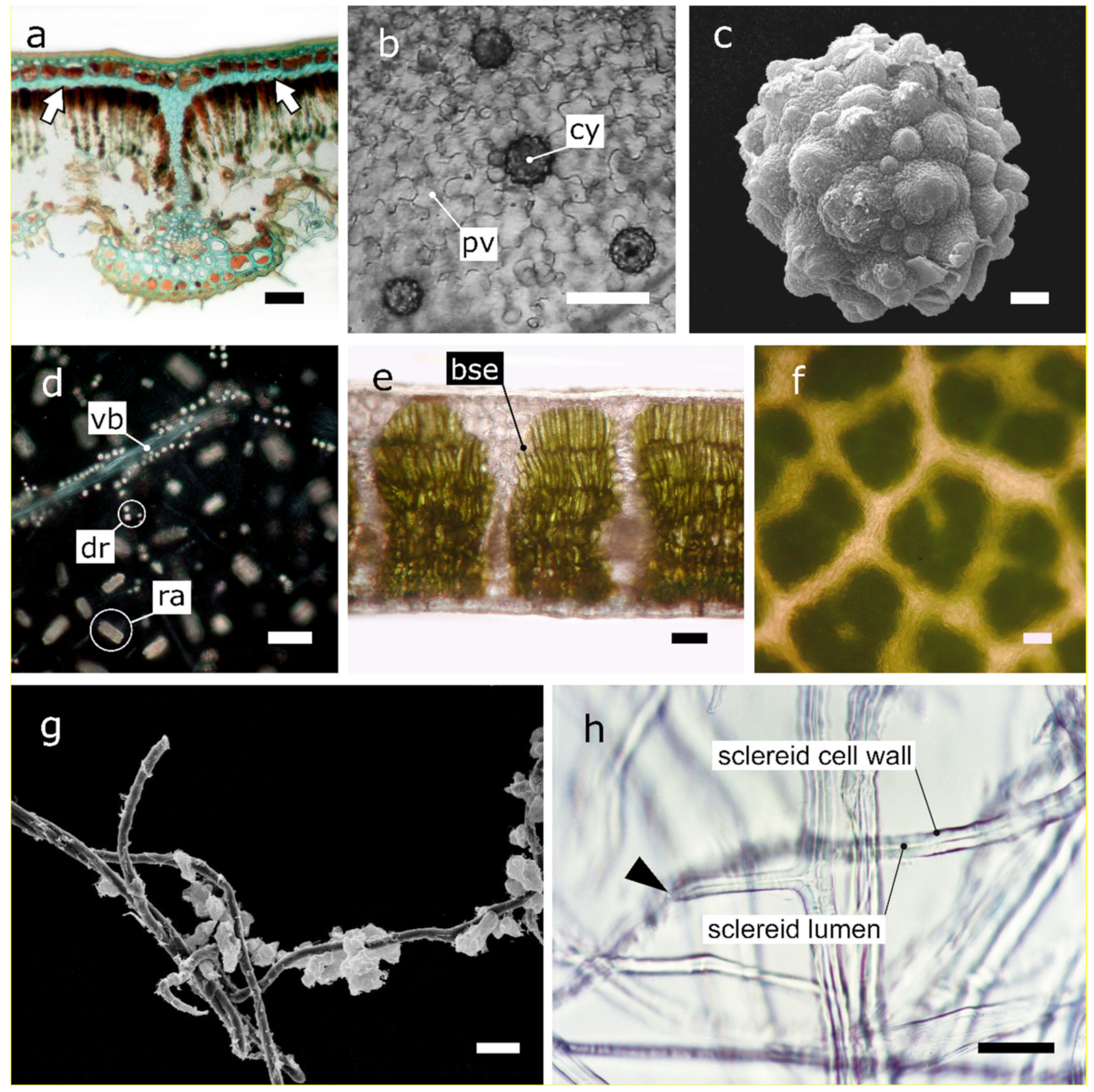
2.3. Phytoliths Protect the Underlying Tissues from Photo-Inhibition
2.4. The Epidermal Cells of Understory Plants Focus Light
2.5. Epidermal Windows Enable Underground Photosynthesis in Succulents
2.6. Non Glandular Trichomes Function as Reflectors and UV Screens
2.7. Superficial Salt Crystals Excreted by Glands Function as UV Screens and Reflectors
2.8. Hypodermal Sclerenchymatic Tissues Protect Mesophyll from UV and Par Damage
3. Mesophyll Structural Elements Allow Efficient Light Propagation and Internal Light Homogenization
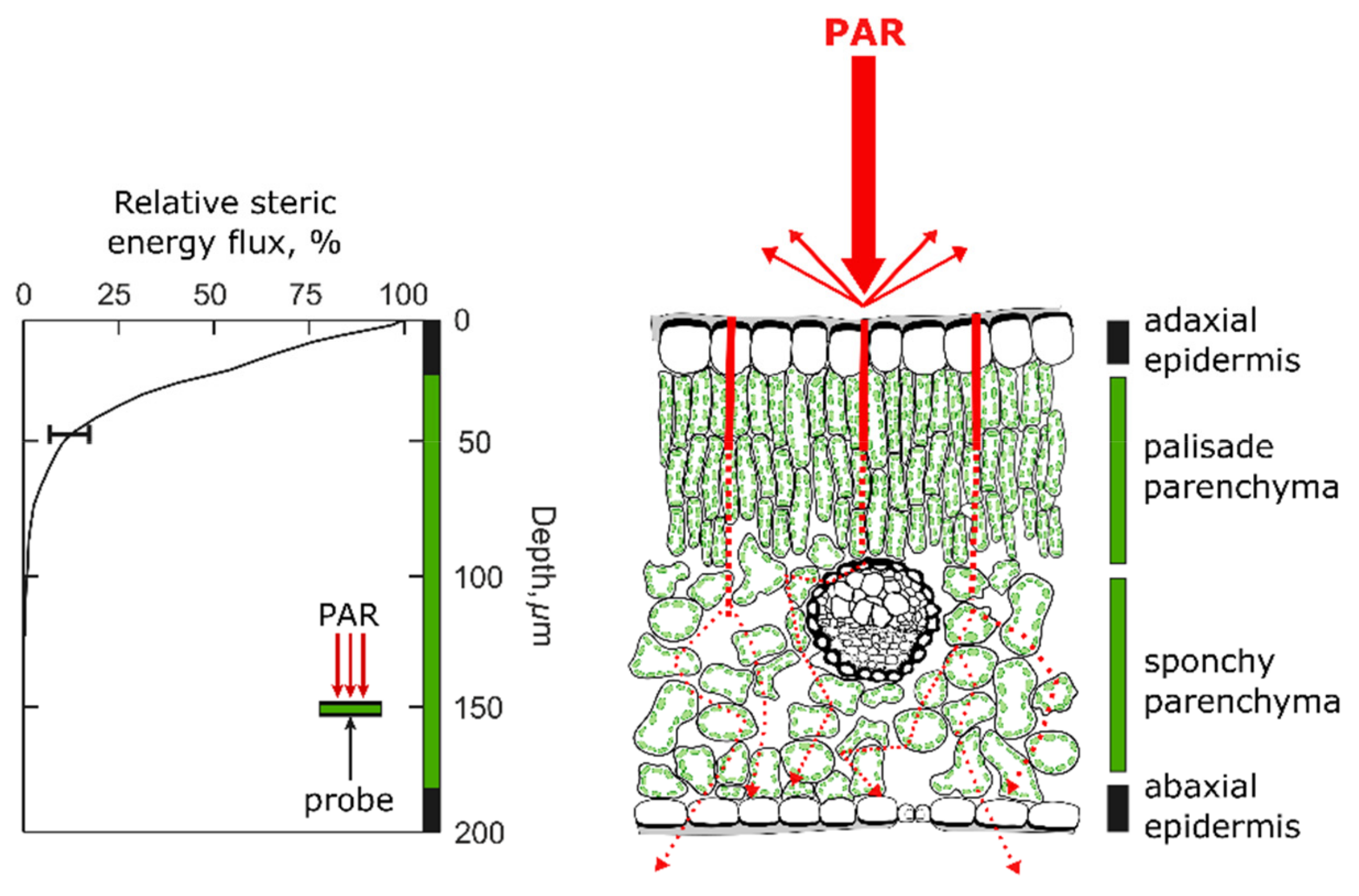
3.1. Mesophyll Cells Affect the Light Interception Efficiency of the Leaves

3.2. Calcium-Carbon Inclusions Improve the Light Microenvironment within Leaves

3.3. Bundle Sheath Extensions Function as Transparent Windows
3.4. Sclereids Function as Optical Fibres
3.5. Fusoid Cells Improve the Light Microenvironment within Mesophyll of Bamboos
4. The Functional Integration with the Other Modules
5. Acclimatization of Optical Structural Elements to Different Light Regimes
6. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, L.; Ma, Z.; Niinemets, Ü.; Guo, D. Three key sub-leaf modules and the diversity of leaf designs. Front. Plant Sci. 2017, 8, 1542. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W. Photosynthetic response to fluctuating environments and photoprotective strategies under abiotic stress. J. Plant Res. 2016, 129, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, T.C. Plant tissue optics. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 231–251. [Google Scholar] [CrossRef]
- Xiao, Y.; Tholen, D.; Zhu, X.G. The influence of leaf anatomy on the internal light environment and photosynthetic electron transport rate: Exploration with a new leaf ray tracing model. J. Exp. Bot. 2016, 67, 6021–6035. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.K.; Vogelmann, T.C.; DeLucia, E.H.; Bell, D.T.; Shepherd, K.A. Leaf form and photosynthesis. Bioscience 1997, 47, 785–793. [Google Scholar] [CrossRef]
- Niinemets, U.; Sack, L. Structural determinants of leaf light-harvesting capacity and photosynthetic potentials. In Progress in Botany. Progress in Botany (Genetics Physiology Systematics Ecology); Esser, K., Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 67, pp. 385–419. [Google Scholar] [CrossRef]
- Vogelmann, T.C.; Gorton, H.L. Leaf: Light capture in the photosynthetic organ. In The Structural Basis of Biological Energy Generation. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Hohmann-Marriott, M., Ed.; Springer: Dordrecht, The Netherlands, 2014; Volume 39, pp. 363–377. [Google Scholar] [CrossRef]
- Meireles, J.E.; Cavender-Bares, J.; Townsend, P.A.; Ustin, S.; Gamon, J.A.; Schweiger, A.K.; Schaepman, M.E.; Asner, G.P.; Martin, R.E.; Singh, A.; et al. Leaf reflectance spectra capture the evolutionary history of seed plants. New Phytol. 2020, 228, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Heredia, A. Biophysical and biochemical characteristics of cutin, a plant barrier biopolymer. Biochim. Biophys. Acta 2003, 1620, 1–7. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Fernández, V.; Bahamonde, H.A.; Peguero-Pina, J.J.; Gil-Pelegrín, E.; Sancho-Knapik, D.; Gil, L.; Goldbach, H.E.; Eichert, T. Physico-chemical properties of plant cuticles and their functional and ecological significance. J. Exp. Bot. 2017, 68, 5293–5306. [Google Scholar] [CrossRef]
- Gausman, H.W.; Rodriguez, R.P.; Escobar, D.E. Ultraviolet radiation reflectance, transmittance, and absorptance by plant leaf epidermises. Agron. J. 1975, 67, 720–724. [Google Scholar] [CrossRef]
- Day, T.A.; Vogelmann, T.C.; DeLucia, E.H. Are some plant life-forms more effective than others in screening out ultraviolet-B radiation? Oecologia 1992, 92, 513–519. [Google Scholar] [CrossRef]
- Day, T.A.; Martin, G.; Vogelmann, T.C. Penetration of UV-B radiation in foliage: Evidence that the epidermis behaves as a non-uniform filter. Plant Cell Environ. 1993, 16, 735–741. [Google Scholar] [CrossRef]
- Rozema, J.; Van De Staaij, J.; Björn, L.O. UV-B as an environmental factor in plant life: Stress and regulation. Trends Ecol. Evol. 1997, 12, 22–28. [Google Scholar] [CrossRef]
- Barnes, P.W.; Flint, S.D.; Ryel, R.J.; Tobler, M.A.; Barkley, A.E.; Wargent, J.J. Rediscovering leaf optical properties: New insights into plant acclimatizationacclimatization to solar UV radiation. Plant Physiol. Biochem. 2015, 93, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Pattison, D.I.; Rahmanto, A.S.; Davies, M.J. Photo-oxidation of proteins. Photochem. Photobiol. Sci. 2012, 11, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Hideg, E.; Jansen, M.A.; Strid, A. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Robson, T.M.; Aphalo, P.J.; Banaś, A.K.; Barnes, P.W.; Brelsford, C.C.; Jenkins, G.I.; Kotilainen, T.K.; Łabuz, J.; Martínez-Abaigar, J.; Morales, L.O.; et al. A perspective on ecologically relevant plant-UV research and its practical application. Photochem. Photobiol. Sci. 2019, 18, 970–988. [Google Scholar] [CrossRef] [Green Version]
- Krauss, P.; Markstädter, C.; Riederer, M. Attenuation of UV radiation by plant cuticles from woody species. Plant Cell Environ. 1997, 20, 1079–1085. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Tzobanoglou, D.; Nikolopoulos, D.; Liakopoulos, G. Epicuticular phenolics over guard cells: Exploitation for in situ stomatal counting by fluorescence microscopy and combined image analysis. Ann. Bot. 2001, 87, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, G.; Stavrianakou, S.; Karabourniotis, G. Analysis of epicuticular phenolics of Prunus persica and Olea europaea leaves: Evidence for the chemical origin of the UV-induced blue fluorescence of stomata. Ann. Bot. 2001, 87, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Solovchenko, A.; Merzlyak, M. Optical properties and contribution of cuticle to UV protection in plants: Experiments with apple fruit. Photochem. Photobiol. Sci. 2003, 2, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Karabourniotis, G.; Liakopoulos, G. Phenolic compounds in plant cuticles: Physiological and ecological aspects. Adv. Plant Physiol. 2005, 8, 33–47. [Google Scholar]
- Liakopoulos, G.; Stavrianakou, S.; Karabourniotis, G. Trichome layers versus dehaired lamina of Olea europaea leaves: Differences in flavonoid distribution, UV-absorbing capacity, and wax yield. Environ. Exp. Bot. 2006, 55, 294–304. [Google Scholar] [CrossRef]
- Pfündel, E.E.; Agati, G.; Cerovic, Z.G. Optical properties of plant surfaces. In Biology of the Plant Cuticle; Riederer, M., Müller, C., Eds.; Blackwell: Oxford, UK, 2006; pp. 216–249. [Google Scholar] [CrossRef]
- Bussotti, F.; Gravano, E.; Grossoni, P.; Tani, C. Occurrence of tannins in leaves of beech trees (Fagus sylvatica L.) along an ecological gradient, detected with an histochemical and ultrastructural analyses. New Phytol. 1998, 138, 469–479. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Ounis, A.; Cartelat, A.; Latouche, G.; Goulas, Y.; Meyer, S.; Moya, I. The use of chlorophyll fluorescence excitation spectra for the non-destructive in situ assessment of UV-absorbing compounds in leaves. Plant Cell Environ. 2002, 25, 1663–1676. [Google Scholar] [CrossRef]
- Manetas, Y. Why some leaves are anthocyanic and why most anthocyanic leaves are red? Flora 2006, 201, 163–177. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P.; Stavroulaki, V.; Sumbele, S. Carbon gain vs. water saving, growth vs. defence: Two dilemmas with soluble phenolics as a joker. Plant Sci. 2014, 227, 21–27. [Google Scholar] [CrossRef]
- Holmes, M.G.; Keiller, D.R. Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: A comparison of a range of species. Plant Cell Environ. 2002, 25, 85–93. [Google Scholar] [CrossRef]
- Koch, K.; Bhushan, B.; Barthlott, W. Multifunctional plant surfaces and smart materials. In Springer Handbook of Nanotechnology. Springer Handbooks; Bhushan, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1399–1436. [Google Scholar] [CrossRef]
- Grant, R.H.; Heisler, G.M.; Gao, W.; Jenks, M. Ultraviolet leaf reflectance of common urban trees and the prediction of reflectance from leaf surface characteristics. Agric. For. Met. 2003, 120, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Mulroy, T.W. Spectral properties of heavily glaucous and nonglaucous leaves of a succulent rosette-plant. Oecologia 1979, 38, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.J. Effect of the angle of incidence of light on the net photosynthesis rates of Sorghum almum leaves. Aust. J. Plant Physiol. 1981, 8, 335–346. [Google Scholar] [CrossRef]
- Nikolopoulos, D.; Bresta, P.; Daliani, V.; Haghiou, V.; Darra, N.; Liati, M.; Mavrogianni, E.; Papanastasiou, A.; Porfyraki, T.; Psaroudi, V.; et al. Photosynthetic rate under oblique illumination depends on the anatomy and the position of the leaf in relation to the direction of light rays. Unpublished work. 2021. [Google Scholar]
- Grant, L. Diffuse and specular characteristics of leaf reflectance. Rem. Sens. Rev. 1987, 22, 309–322. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef]
- Goto, M.; Ehara, H.S.; Karita, S.; Takabe, K.; Ogawa, N.; Yamada, Y.; Ogawa, S.; Yahaya, M.S.; Morita, O. Protective effect of silicon on phenolic biosynthesis and ultraviolet spectral stress in rice crop. Plant Sci. 2003, 164, 349–356. [Google Scholar] [CrossRef]
- Klančnik, K.; Vogel-Mikuš, K.; Gaberščik, A. Silicified structures affect leaf optical properties in grasses and sedge. J. Photochem. Photobiol. B Biol. 2014, 130, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Klančnik, K.; Vogel-Mikuš, K.; Kelemen, M.; Vavpetic, P.; Pelicon, P.; Kump, P.; Jezeršek, D.; Gianoncelli, A.; Gaberščik, A. Leaf optical properties are affected by the location and type of deposited biominerals. J. Photochem. Photobiol. B Biol. 2014, 140, 276–285. [Google Scholar] [CrossRef]
- Pierantoni, M.; Tenne, R.; Brumfeld, V.; Kiss, V.; Oron, D.; Addadi, L.; Weiner, S. Plants and light manipulation: The integrated mineral system in okra leaves. Adv. Sci. 2017, 4, 1600416. [Google Scholar] [CrossRef]
- Agarie, S.; Agata, W.; Uchida, H.; Kubota, F.; Kaufman, P.B. Function of silica bodies in the epidermal system of rice (Oryza sativa L.): Testing the window hypothesis. J. Exp. Bot. 1996, 47, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Haberlandt, G. Physiologische Pflanzenanatomie; W. Engelmann: Leipzig, Germany, 1914. [Google Scholar]
- Oguchi, R.; Onoda, Y.; Terashima, I.; Tholen, D. Leaf anatomy and function. In The Leaf: A Platform for Performing Photosynthesis. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 44, pp. 97–139. [Google Scholar] [CrossRef]
- Ustin, S.L.; Jacquemoud, S. How the optical properties of leaves modify the absorption and scattering of energy and enhance leaf functionality. In Remote Sensing of Plant Biodiversity; Cavender-Bares, J., Gamon, J., Townsend, P., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 349–384. [Google Scholar] [CrossRef]
- Bone, R.A.; Lee, D.; Norman, J.M. Epidermal cells functioning as lenses in leaves of tropical rain-forest shade plants. Appl. Opt. 1985, 24, 1408–1412. [Google Scholar] [CrossRef]
- Vogelmann, T.C.; Bornman, J.F.; Yates, D.J. Focusing of light by leaf epidermal cells. Physiol. Plant. 1996, 98, 43–56. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Vogelmann, T.C. Do epidermal lens cells facilitate the absorptance of diffuse light? Am. J. Bot. 2007, 94, 1061–1066. [Google Scholar] [CrossRef] [Green Version]
- Gorton, H.L.; Vogelmann, T.C. Effects of epidermal cell shape and pigmentation on optical properties of Antirrhinum petals in the visible and ultraviolet. Plant Physiol. 1996, 112, 879–888. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.E.; Brandmeyer, E.A.; Ross, R.D. Ecophysiological function of leaf ‘windows’ in Lithops species—‘Living Stones’ that grow underground. Plant Biol. 2013, 15, 243–247. [Google Scholar] [CrossRef]
- Krulik, G.A. Light transmission in window-leaved plants. Can. J. Bot. 1980, 58, 1591–1600. [Google Scholar] [CrossRef]
- Morre, R.; Langekamp, M. Tissue partitioning during leaf development in ornamentally-grown Frithia pulchra (Mesembryanthemaceae), a ‘Window Plant’. Ann. Bot. 1991, 67, 279–283. [Google Scholar] [CrossRef]
- Egbert, K.J.; Martin, C.E. Light penetration via leaf windows does not increase photosynthesis in three species of desert succulents. J. Plant Physiol. 2000, 157, 521–525. [Google Scholar] [CrossRef]
- Eller, B.M.; Brinckmann, E.; von Willert, D.J. Optical properties and succulence of plants in the arid Richtersveld (Cp., Rep. South Africa). Bot. Helv. 1983, 93, 47–55. [Google Scholar] [CrossRef]
- Egbert, K.J.; Martin, C.E.; Vogelmann, T.C. The influence of epidermal windows on the light environment within the leaves of six succulents. J. Exp. Bot. 2008, 59, 1863–1873. [Google Scholar] [CrossRef]
- Egbert, K.J.; Martin, C.E. The Influence of leaf windows on the utilization and absorption of radiant energy in seven desert succulents. Photosynthetica 2002, 40, 35–39. [Google Scholar] [CrossRef]
- Field, K.J.; George, R.; Fearn, B.; Quick, W.P.; Davey, M.P. Best of both worlds: Simultaneous high-light and shade-tolerance adaptations within individual leaves of the living stone Lithops aucampiae. PLoS ONE 2013, 8, e75671. [Google Scholar] [CrossRef] [Green Version]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar] [CrossRef]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant glandular trichomes: Natural cell factories of high biotechnological interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [Green Version]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure-function coordination. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Karabourniotis, G.; Bornman, J.F. Penetration of UV-A, UV-B and blue light through the leaf trichome layers of two xeromorphic plants, olive and oak, measured by optical fibre microprobes. Physiol. Plant. 1999, 105, 655–661. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Bornman, J.F.; Liakoura, V. Different leaf surface characteristics of three grape cultivars affect leaf optical properties as measured with fibre optics: Possible implication in stress tolerance. Aust. J. Plant Physiol. 1999, 26, 47–53. [Google Scholar] [CrossRef]
- Liakoura, V.; Stefanou, M.; Manetas, Y.; Cholevas, C.; Karabourniotis, G. Trichome density and its UV-B protective potential are affected by shading and leaf position on the canopy. Environ. Exp. Bot. 1997, 38, 223–229. [Google Scholar] [CrossRef]
- Yan, A.; Pan, J.; An, L.; Gan, Y.; Feng, H. The responses of trichome mutants to enhanced ultraviolet-B radiation in Arabidopsis thaliana. J. Photochem. Photobiol. B Biol. 2012, 113, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Skaltsa, H.; Verykokidou, E.; Harvala, C.; Karanourniotis, G.; Manetas, Y. UV-B protective potential and flavonoid content of leaf hairs of Quercus ilex. Phytochemistry 1994, 37, 987–990. [Google Scholar] [CrossRef]
- Ntefidou, M.; Manetas, Y. Optical properties of hairs during the early stages of leaf development in Platanus orientalis. Aust. J. Plant Physiol. 1996, 23, 535–538. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Kofidis, G.; Fasseas, C.; Liakoura, V.; Drossopoulos, I. Polyphenol deposition on leaf hairs of Olea europaea (Oleaceae) and Quercus ilex (Fagaceae). Am. J. Bot. 1998, 85, 1007–1012. [Google Scholar] [CrossRef]
- Manetas, Y. The importance of being hairy: The adverse effects of hair removal on stem photosynthesis of Verbascum speciosum are due to solar UV-B radiation. New Phytol. 2003, 158, 503–508. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Bickford, C.P. Ecophysiology of leaf trichomes. Funct. Plant Biol. 2016, 43, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Ehleringer, J.R. Ecology and ecophysiology of leaf pubescence in North American plants. In Biology and Chemistry of Plant Trichomes; Rodriguez, E., Healey, P.L., Mehta, I., Eds.; Plenum Press: New York, NY, USA, 1984; pp. 113–132. [Google Scholar]
- Ehleringer, J. Comparative microclimatology and plant responses in Encelia species from contrasting habitats. J. Arid Environ. 1985, 8, 45–56. [Google Scholar] [CrossRef]
- Lang, M.; Schindler, C. The effect of leaf-hairs on blue and red fluorescence emission and on zeaxanthin cycle performance of Senecio medley L. J. Plant Physiol. 1994, 144, 680–685. [Google Scholar] [CrossRef]
- Bisba, A.; Petropoulou, Y.; Manetas, Y. The transiently pubescent young leaves of plane (Platanus orientalis) are deficient in photodissipative capacity. Physiol. Plant. 1997, 101, 373–378. [Google Scholar] [CrossRef]
- Liakopoulos, G.; Nikolopoulos, D.; Klouvatou, A.; Vekkos, K.-A.; Manetas, Y.; Karabourniotis, G. The photoprotective role of epidermal anthocyanins and surface pubescence in young leaves of grapevine (Vitis vinifera). Ann. Bot. 2006, 98, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.-J.; Chow, W.S.; Liu, X.-T.; Zhang, P.; Liu, N.; Peng, C.-L. A magic red coat on the surface of young leaves: Anthocyanins distributed in trichome layer protect Castanopsis fissa leaves from photoinhibition. Tree Physiol. 2016, 36, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, T.C.; Martin, G. The functional significance of palisade tissue: Penetration of directional versus diffuse light. Plant Cell Environ. 1993, 16, 65–72. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Vogelmann, T.C.; Williams, W.E.; Gorton, H.L. A new paradigm in leaf-level photosynthesis: Direct and diffuse lights are not equal. Plant Cell Environ. 2008, 31, 159–164. [Google Scholar] [CrossRef]
- Gorton, H.L.; Brodersen, C.R.; Williams, W.E.; Vogelmann, T.C. Measurement of the optical properties of leaves under diffuse light. Photochem. Photobiol. 2010, 86, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Bingying, L.; Wang, B. Progress in studying salt secretion from the salt glands in recretohalophytes: How do plants secrete salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Fernández-Marín, B.; Hernandez, A.; Jiménez, E.T.; León, A.; García-Mauriño, S.; Silva, C.D.; Dolmus, J.R.; Dolmus, C.M.; Molina, M.J.; et al. Salt crystal deposition as a reversible mechanism to enhance photoprotection in black mangrove. Trees 2013, 27, 229–237. [Google Scholar] [CrossRef]
- Grašič, M.; Budak, V.; Klančnik, K.; Gaberščik, A. Optical properties of halophyte leaves are affected by the presence of salt on the leaf surface. Biologia 2017, 72, 1131–1139. [Google Scholar] [CrossRef]
- Jordan, G.J.; Carpenter, R.J.; Hill, R.S. The macrofossil record of Proteaceae in Tasmania: A review with new species. Aust. Syst. Bot. 1998, 11, 465–501. [Google Scholar] [CrossRef] [Green Version]
- Jordan, G.J.; Dillon, R.A.; Weston, P.H. Solar radiation as a factor in the evolution of scleromorphic leaf anatomy in Proteaceae. Am. J. Bot. 2005, 92, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Jordan, G.J.; Brodribb, T.; Blackman, C.; Weston, P. Climate drives vein anatomy in Proteaceae. Am. J. Bot. 2013, 100, 1483–1493. [Google Scholar] [CrossRef]
- Terashima, I.; Saeki, T. Light environment within a leaf. I. Optical properties of paradermal sections of Camellia leaves with special reference to differences in the optical properties of palisade and spongy tissues. Plant Cell Physiol. 1983, 24, 1493–1501. [Google Scholar] [CrossRef]
- Vogelmann, T.C.; Evans, J.R. Profiles of light absorption and chlorophyll within spinach leaves from chlorophyll fluorescence. Plant Cell Environ. 2002, 25, 1313–1323. [Google Scholar] [CrossRef]
- Vogelman, T.C. Penetration of light into plants. Photochem. Photobiol. 1989, 50, 895–902. [Google Scholar] [CrossRef]
- Terashima, I. Anatomy of non-uniform leaf photosynthesis. Photosynth. Res. 1992, 31, 195–212. [Google Scholar] [CrossRef] [PubMed]
- DeLucia, E.H.; Nelson, K.; Vogelmann, T.C.; Smith, W.K. Contribution of intercellular reflectance to photosynthesis in shade leaves. Plant Cell Environ. 1996, 19, 159–170. [Google Scholar] [CrossRef]
- Evans, J.R.; Vogelmann, T.C. Profiles of 14C fixation through spinach leaves in relation to light absorption and photosynthetic capacity. Plant Cell Environ. 2003, 26, 547–560. [Google Scholar] [CrossRef]
- Théroux-Rancourt, G.; Roddy, A.B.; Earles, J.M.; Gilbert, M.E.; Zwieniecki, M.A.; Boyce, C.K.; Tholen, D.; McElrone, A.J.; Simonin, K.A.; Brodersen, C.R. Maximum CO2 diffusion inside leaves is limited by the scaling of cell size and genome size. Proc. R. Soc. B 2020, 288, 20203145. [Google Scholar] [CrossRef] [PubMed]
- Giannopoulos, A.; Nikolopoulos, D.; Bresta, P.; Samantas, A.; Reppa, C.; Karaboiki, K.; Dotsika, E.; Fasseas, C.; Liakopoulos, G.; Karabourniotis, G. Cystoliths of Parietaria judaica can serve as an internal source of CO2 for photosynthetic assimilation when stomata are closed. J. Exp. Bot. 2019, 70, 5753–5763. [Google Scholar] [CrossRef] [PubMed]
- Bauer, P.; Elbaum, R.; Weiss, I.M. Calcium and silicon mineralization in land plants: Transport, structure and function. Plant Sci. 2012, 180, 746–756. [Google Scholar] [CrossRef]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2013, 19, 166–174. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Horner, H.T.; Bresta, P.; Nikolopoulos, D.; Liakopoulos, G. New insights into the functions of carbon-calcium inclusions in plants. New Phytol. 2020, 228, 845–854. [Google Scholar] [CrossRef]
- Horner, H.T.; Wanke, S.; Samain, M. A comparison of leaf crystal macropatterns in the two sister genera Piper and Peperomia (Piperaceae). Am. J. Bot. 2012, 99, 983–997. [Google Scholar] [CrossRef] [Green Version]
- Kuo-Huang, L.L.; Ku, M.S.B.; Franceschi, V.R. Correlations between calcium oxalate crystals and photosynthetic activities in palisade cells of shade-adapted Peperonia glabella. Bot. Stud. 2007, 48, 155–164. [Google Scholar]
- Gal, A.; Brumfeld, V.; Weiner, S.; Addadi, L.; Oron, D. Certain biominerals in leaves function as light scatterers. Adv. Mater. 2012, 24, OP77–OP83. [Google Scholar] [CrossRef]
- Pierantoni, M.; Tenne, R.; Rephael, B.; Brumfeld, V.; van Casteren, A.; Kupczik, K.; Oron, D.; Addadi, L.; Weiner, S. Mineral deposits in Ficus leaves: Morphologies and locations in relation to function. Plant Physiol. 2018, 176, 1751–1763. [Google Scholar] [CrossRef] [Green Version]
- Karabourniotis, G.; Bornman, J.F.; Nikolopoulos, D. A possible optical role of the bundle sheath extensions of the heterobaric leaves of Vitis vinifera and Quercus coccifera. Plant Cell Environ. 2000, 23, 423–430. [Google Scholar] [CrossRef]
- Horner, H.T.; Wanke, S.; Oelschlägel, B.; Samain, M.-S. Peruvian window-leaved Peperomia taxa display unique crystal macropatterns in high-altitude environments. Int. J. Plant Sci. 2017, 178, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Wylie, R.B. The bundle sheath extension in leaves of dicotyledons. Am. J. Bot. 1952, 39, 645–651. [Google Scholar] [CrossRef]
- McClendon, J.H. Photographic survey of the occurrence of bundle-sheath extensions in deciduous dicots. Plant Physiol. 1992, 99, 1677–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, K. Plant Anatomy, 2nd ed.; Wiley: New York, NY, USA, 1965. [Google Scholar]
- Nikolopoulos, D.; Liakopoulos, G.; Drossopoulos, I.; Karabourniotis, G. The relationship between anatomy and photosynthetic performance of heterobaric leaves. Plant Physiol. 2002, 129, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Liakoura, V.; Fotelli, M.N.; Rennenberg, H.; Karabourniotis, G. Should structure-function relations be considered separately for homobaric vs. heterobaric leaves? Am. J. Bot. 2009, 96, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.G.; Maiti, R.; Aruna Kumari, C.H. Comparative anatomy of leaf lamina of twenty six woody species of Tamaulipan thornscrub from northeastern Mexico and its significance in taxonomic delimitation and adaptation of the species to xeric environments. Pak. J. Bot. 2017, 49, 589–596. [Google Scholar]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Hiromi, T. Ecological distribution of homobaric and heterobaric leaves in tree species of Malaysian lowland tropical rainforest. Am. J. Bot. 2007, 94, 764–775. [Google Scholar] [CrossRef]
- Boeger, M.R.T.; Silva, M.M.; Nogueira, G.; Alvarenga, A.; Pereto, S.S. Occurrence of homobaric and heterobaric leaves in two forest types of southern Brazil. Acta Bot. Bras. 2016, 30, 304–312. [Google Scholar] [CrossRef] [Green Version]
- Kenzo, T.; Tanaka-Oda, A.; Mastuura, Y.; Hinzman, L.D. Morphological and physicochemical traits of leaves of different life-forms of various broadleaf woody plants in interior Alaska. Can. J. For. Res. 2016, 46, 1475–1482. [Google Scholar] [CrossRef]
- Roden, J.S.; Pearcy, R.W. Effect of leaf flutter on the light environment of poplars. Oecologia 1993, 93, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.S.; Pearcy, R.W. Photosynthetic gas exchange response of poplars to steady-state and dynamic light environments. Oecologia 1993, 93, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.S. Modeling the light interception and carbon gain of individual fluttering aspen (Populus tremuloides Michx) leaves. Trees Struct. Funct. 2003, 17, 117–126. [Google Scholar] [CrossRef]
- Dickison, W.G. Integrative Plant Anatomy; Harcourt/Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Karabourniotis, G.; Papastergiou, N.; Kabanopoulou, E.; Fasseas, C. Foliar sclereids of Olea europaea may function as optical fibres. Can. J. Bot. 1994, 72, 330–336. [Google Scholar] [CrossRef]
- Karabourniotis, G. Light-guiding function of foliar sclereids in the evergreen sclerophyll Phillyrea latifolia: A quantitative approach. J. Exp. Bot. 1998, 49, 739–746. [Google Scholar] [CrossRef]
- March, R.H.; Clark, L.G. Sun-shade variation in bamboo (Poaceae: Bambusoideae) leaves. Telopea 2011, 13, 93–104. [Google Scholar] [CrossRef]
- Sack, L.; Buckley, T.N. Trait multi-functionality in plant stress response. Integr. Comp. Biol. 2020, 60, 98–112. [Google Scholar] [CrossRef]
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Plant Anatomy: A Concept-Based Approach to the Structure of Seed Plants; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Javelle, M.; Vernoud, V.; Rogowsky, P.M.; Ingram, G.C. Epidermis: The formation and functions of a fundamental plant tissue. New Phytol. 2011, 189, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.; Furstner, R.; Barthlott, W.; Neinhuis, C. Quantitative assessment to the structural basis of water repellency in natural and technical surfaces. J. Exp. Bot. 2003, 54, 1295–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhushan, B.; Jung, Y.C. Micro- and nanoscale characterization of hydrophobic and hydrophilic leaf surfaces. Nanotechnology 2006, 17, 2758–2772. [Google Scholar] [CrossRef]
- Majer, P.; Neugar, S.; Krumbein, A.; Schreiner, M.; Hideg, É. Singlet oxygen scavenging by leaf flavonoids contributes to sunlight acclimatizationacclimatization in Tilia platyphyllos. Environ. Exp. Bot. 2014, 100, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, C.R. Some basic types of cells and tissues. In Anatomy of the Dicotyledons, 2nd ed.; Metcalfe, C.R., Chalk, L., Eds.; Clarendon Press: Oxford, UK, 1979; Volume 1, pp. 54–62, 207–214. [Google Scholar]
- Kawai, K.; Miyoshi, R.; Okada, N. Bundle sheath extensions are linked to water relations but not to mechanical and structural properties of leaves. Trees 2017, 31, 1227–1237. [Google Scholar] [CrossRef]
- Heide-Jorgensen, H.S. Xeromorphic leaves of Hakea suaveolens R. Br. IV. Ontogeny, structure and function of the sclereids. Aust. J. Bot. 1990, 38, 25–43. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Brodribb, T.J.; Holbrook, N.M. Hydraulic design of leaves: Insights from rehydration kinetics. Plant Cell Environ. 2007, 30, 910–921. [Google Scholar] [CrossRef]
- Scoffoni, C.; Pou, A.; Aasamaa, K.; Sack, L. The rapid light response of leaf hydraulic conductance: New evidence from two experimental methods. Plant Cell Environ. 2008, 31, 1803–1812. [Google Scholar] [CrossRef]
- Zsögön, A.; Alves Negrini, A.C.; Peres, L.E.P.; Nguyen, H.T.; Ball, M.C. A mutation that eliminates bundle sheath extensions reduces leaf hydraulic conductance, stomatal conductance and assimilation rates in tomato (Solanum lycopersicum). New Phytol. 2015, 205, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Leegood, R.C. Roles of the bundle sheath cells in leaves of C3 plants. J. Exp. Bot. 2008, 59, 1663–1673. [Google Scholar] [CrossRef]
- Buckley, T.N.; Sack, L.; Gilbert, M.E. The role of bundle sheath extensions and life form in stomatal responses to leaf water status. Plant Physiol. 2011, 156, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Buckley, T.N.; John, G.P.; Scoffoni, C.; Sack, L. How does leaf anatomy influence water transport outside the xylem? Plant Physiol. 2015, 168, 1616–1635. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Glenn, E.P.; Volkov, V. Could vesicular transport of Na+ and Cl− be a feature of salt tolerance in halophytes? Ann. Bot. 2019, 123, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Yun, P.; Shabala, S. Ion transport in salt glands and bladders in halophyte species. In Handbook of Halophytes; Grigore, M.-N., Ed.; Springer: Cham, Switzerland, 2020; pp. 1–19. [Google Scholar] [CrossRef]
- Vieira, R.C.; Gomes, D.M.S.; Sarhyba, L.S.; Arruda, R.C.O. Leaf anatomy of three herbaceous bamboo species. Braz. J. Biol. 2002, 62, 907–922. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, H.; Lin, S.; Hse, C.; Ding, Y. Anatomical characteristics of fusoid cells and vascular bundles in Fargesia yunnanensis leaves. J. For. Res. 2016, 27, 1237–1247. [Google Scholar] [CrossRef]
- Tooulakou, G.; Giannopoulos, A.; Nikolopoulos, D.; Bresta, P.; Dotsika, E.; Orkoula, M.G.; Kontoyannis, C.G.; Fasseas, C.; Liakopoulos, G.; Klapa, M.I.; et al. Alarm photosynthesis: Calcium oxalate crystals as an internal CO2 source in plants. Plant Physiol. 2016, 171, 2577–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strömberg, C.A.E.; Di Stilio, V.S.; Song, Z. Functions of phytoliths in vascular plants: An evolutionary perspective. Funct. Ecol. 2016, 30, 1286–1297. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, E.T.; Orcutt, D.M. The Physiology of Plants under Stress; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Korgiopoulou, C.; Bresta, P.; Nikolopoulos, D.; Karabourniotis, G. Sex-specific structural and functional leaf traits and sun-shade acclimatizationacclimatization in the dioecious tree Pistacia vera (Anacardiaceae). Funct. Plant Biol. 2019, 46, 649–659. [Google Scholar] [CrossRef]
- Ashton, P.M.S.; Berlyn, G.P. A comparison of leaf physiology and anatomy of Quercus (section Erythrobalanus-Fagaceae) species in different light environments. Am. J. Bot. 1994, 81, 589–597. [Google Scholar] [CrossRef]
- Talhouët, A.-C.; Meyer, S.; Baudin, X.; Streb, P. Dynamic acclimatizationacclimatization to sunlight in an alpine plant, Soldanella alpina L. Physiol. Plant. 2020, 168, 563–575. [Google Scholar] [CrossRef]
- Vega, C.; González, G.; Bahamonde, H.A.; Valbuena-Carabaña, M.; Gil, L.; Fernández, V. Effect of irradiation and canopy position on anatomical and physiological features of Fagus sylvatica and Quercus petraea leaves. Plant Physiol. Biochem. 2020, 152, 232–242. [Google Scholar] [CrossRef]
- Manetas, Y.; Petropoulou, Y.; Stamatakis, K.; Nikolopoulos, D.; Levizou, E.; Psaras, G.; Karabourniotis, G. Beneficial effects of enhanced UV-B radiation under field conditions: Improvement of needle water relations and survival capacity of Pinus pinea L. seedlings during the dry Mediterranean summer. Plant Ecol. 1997, 128, 101–108. [Google Scholar] [CrossRef]
- Drilias, P.; Karabourniotis, G.; Levizou, E.; Nikolopoulos, D.; Petropoulou, Y.; Manetas, Y. The effects of enhanced UV-B radiation on the Mediterranean evergreen sclerophyll Nerium oleander depend on the extent of summer precipitation. Funct. Plant Biol. 1997, 24, 301–306. [Google Scholar] [CrossRef]
- Bohlar-Nordenkampf, H.R.; Draxler, G. Functional leaf anatomy. In Photosynthesis and Production in a Changing Environment. A Field and Laboratory Manual; Hall, D.O., Schurlock, J.M.O., Bolhar-Nordenkampf, H.R., Leegood, R.C., Long, S.P., Eds.; Springer: Dordrecht, Germany, 1993; pp. 91–112. [Google Scholar] [CrossRef]
- Roth-Nebelsick, A.; Uhl, D.; Mosbrugger, V.; Kerp, H. Evolution and function of leaf venation architecture: A review. Ann. Bot. 2001, 87, 553–566. [Google Scholar] [CrossRef]
- Lynch, D.J.; McInerney, F.A.; Kouwenberg, L.L.; Gonzalez-Meler, M.A. Plasticity in bundle sheath extensions of heterobaric leaves. Am. J. Bot. 2012, 99, 1197–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, M.A.M.; Chitwood, D.H.; Azevedo, A.A.; Araújo, W.L.; Ribeiro, D.M.; Peres, L.E.P.; Martins, S.C.V.; Zsögön, A. Bundle sheath extensions affect leaf structural and physiological plasticity in response to irradiance. Plant Cell Environ. 2019, 42, 1575–1589. [Google Scholar] [CrossRef] [Green Version]
- Hauser, M.-T. Molecular basis of natural variation and environmental control of trichome patterning. Front. Plant. Sci. 2014, 5, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Václavík, T.; Beckmann, M.; Cord, A.F.; Bindewald, A.M. Effects of UV-B radiation on leaf hair traits of invasive plants-Combining historical herbarium records with novel remote sensing data. PLoS ONE 2017, 12, e0175671. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Gravano, E.; Pinelli, P.; Mulinaci, N.; Romani, A. Flavonoids accumulate in leaves and glandular trichomes of Phillyrea latifolia exposed to excess solar radiation. New Phytol. 2000, 148, 69–77. [Google Scholar] [CrossRef]
- Yamasaki, S.; Noguchi, N.; Mimaki, K. Continuous UV-B irradiation induces morphological changes and the accumulation of polyphenolic compounds on the surface of cucumber cotyledons. J. Rad. Res. 2007, 48, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, S.; Murakami, Y. Continuous UV-B irradiation induces endoreduplication and trichome formation in cotyledons, and reduces epidermal cell division and expansion in the first leaves of pumpkin seedlings (Cucurbita maxima Duch. × C. moschata Duch.). Environ. Cont. Biol. 2014, 52, 203–209. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karabourniotis, G.; Liakopoulos, G.; Bresta, P.; Nikolopoulos, D. The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection. Plants 2021, 10, 1455. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071455
Karabourniotis G, Liakopoulos G, Bresta P, Nikolopoulos D. The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection. Plants. 2021; 10(7):1455. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071455
Chicago/Turabian StyleKarabourniotis, George, Georgios Liakopoulos, Panagiota Bresta, and Dimosthenis Nikolopoulos. 2021. "The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection" Plants 10, no. 7: 1455. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071455