
Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Growth Parameters
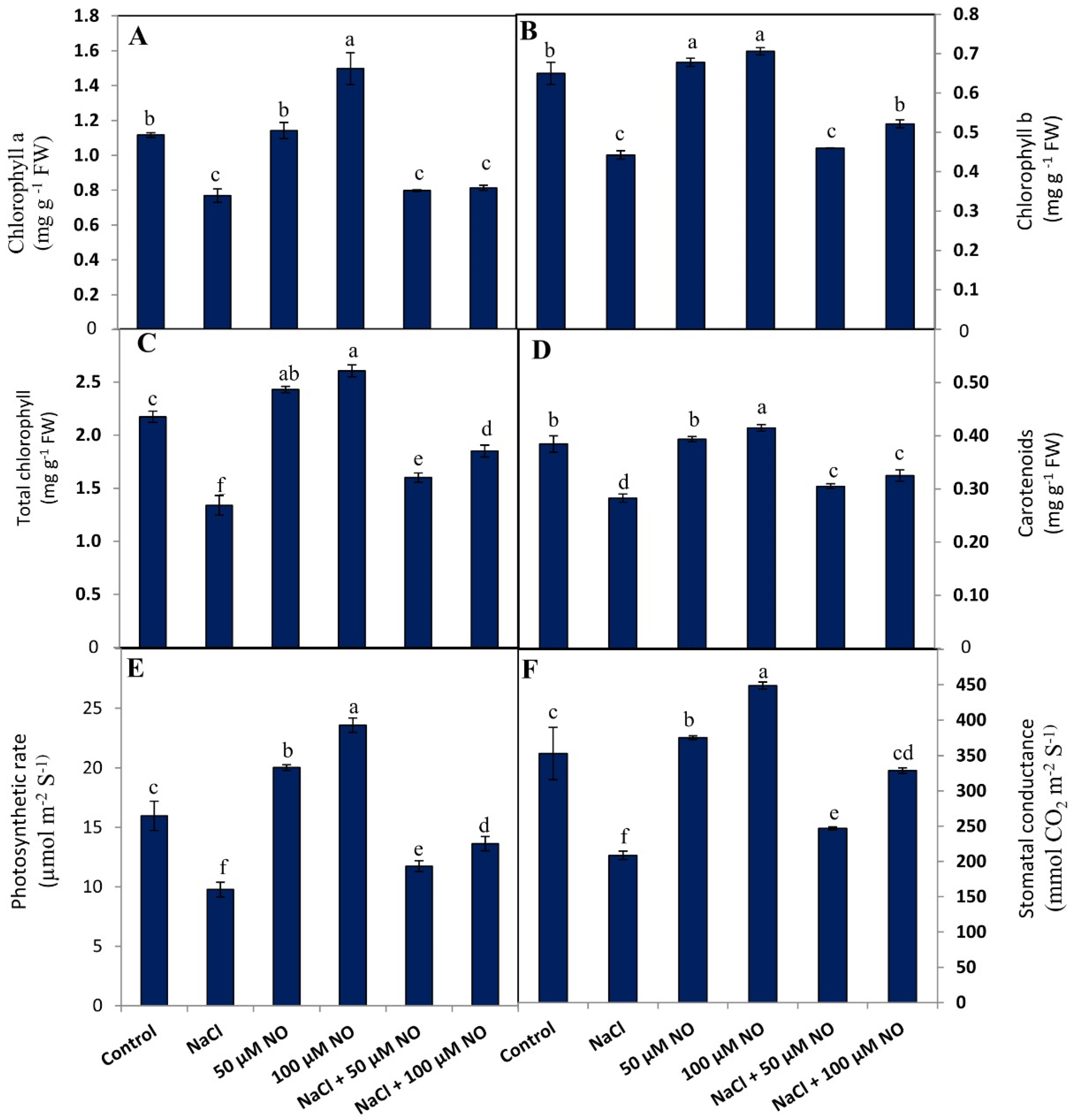
2.2. No Protects Chlorophyll and Photosynthetic Attributes in Salt-Stressed Wheat Plants
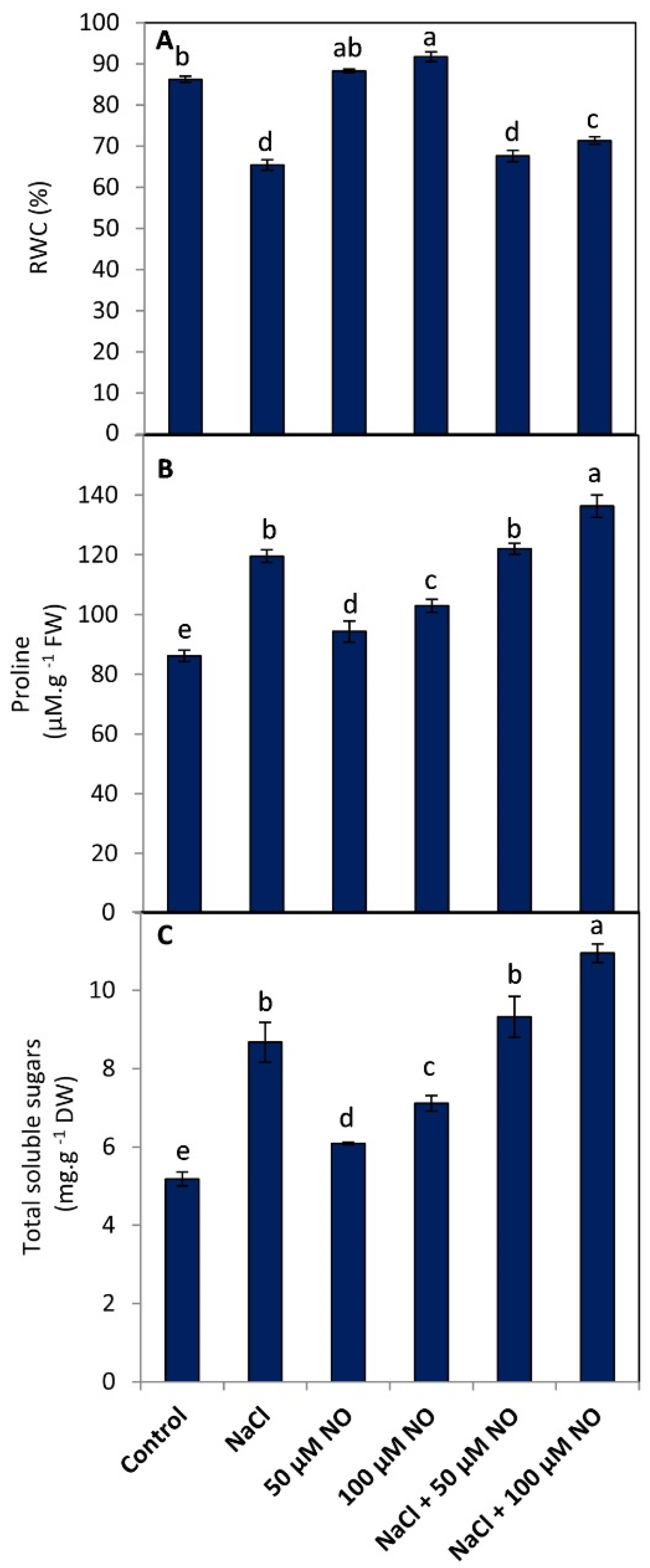
2.3. No Maintains Leaf RWC by Improving the Proline and Sugar Content
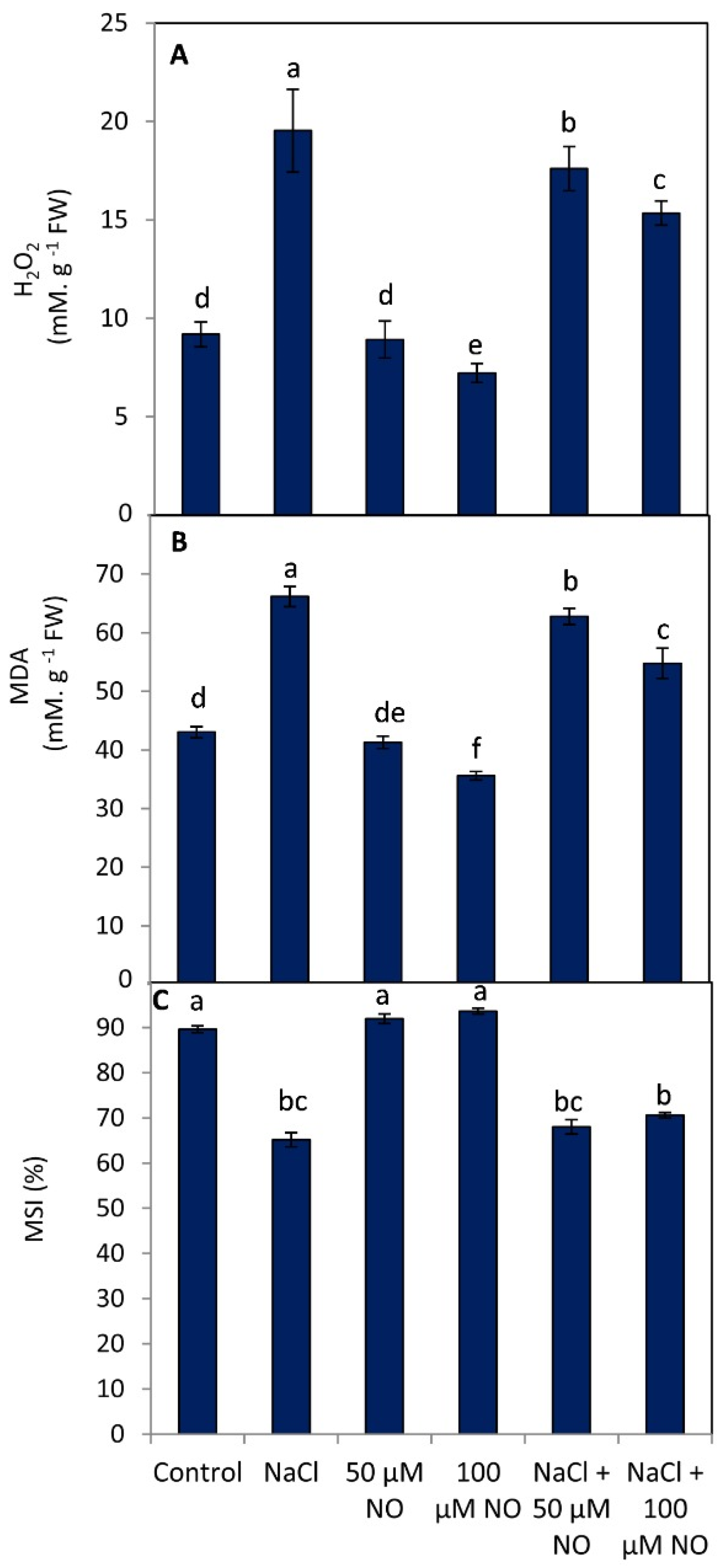
2.4. No Application Reduces Oxidative Damage by Preventing Lipid Peroxidation, Hydrogen Peroxide, and Improving Membrane Stability Index
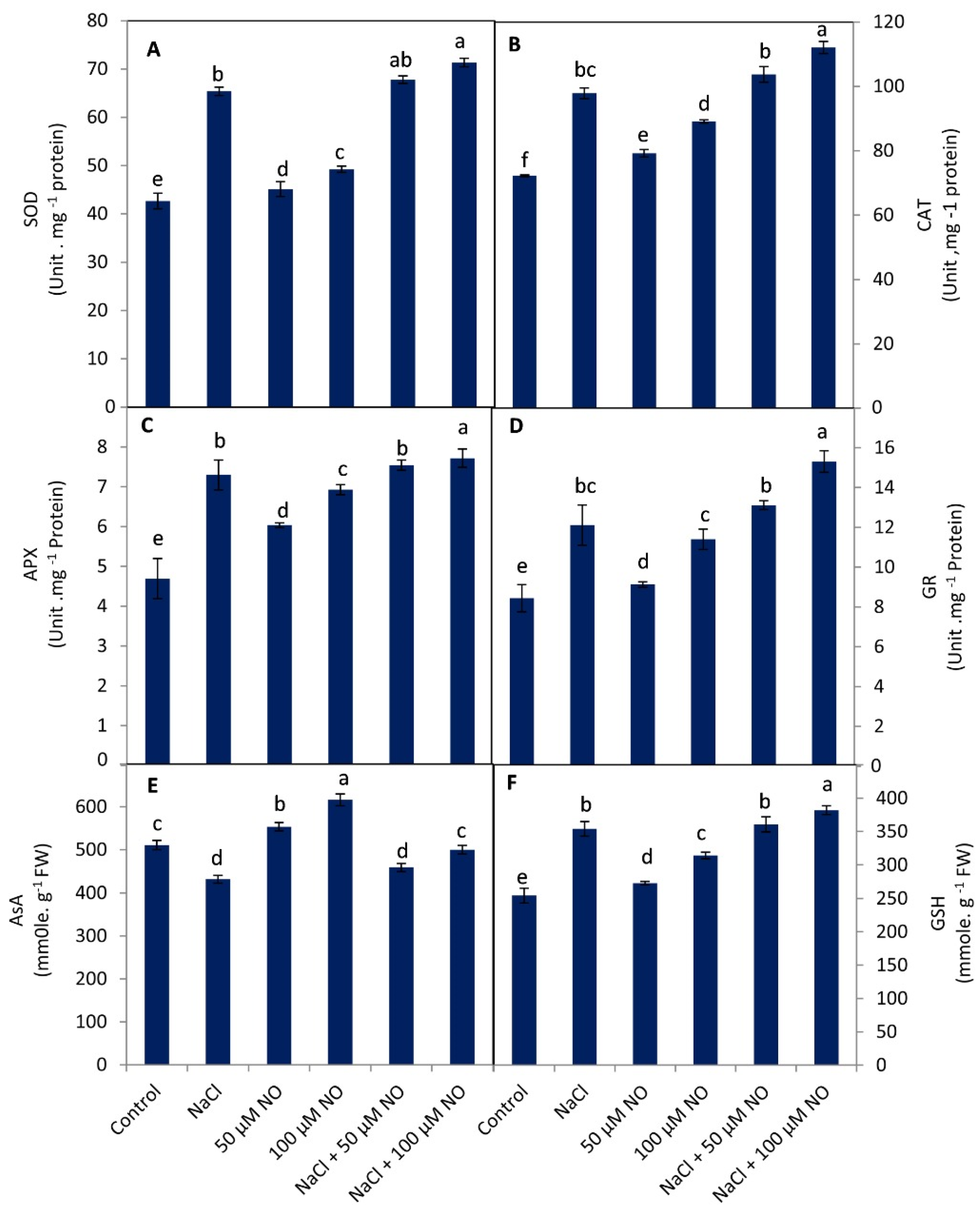
2.5. Exogenous NO Up-Regulates Antioxidant System in Salinity-Stressed Wheat
2.6. No Improves Uptake of N, K, and Ca under Salinity Stress
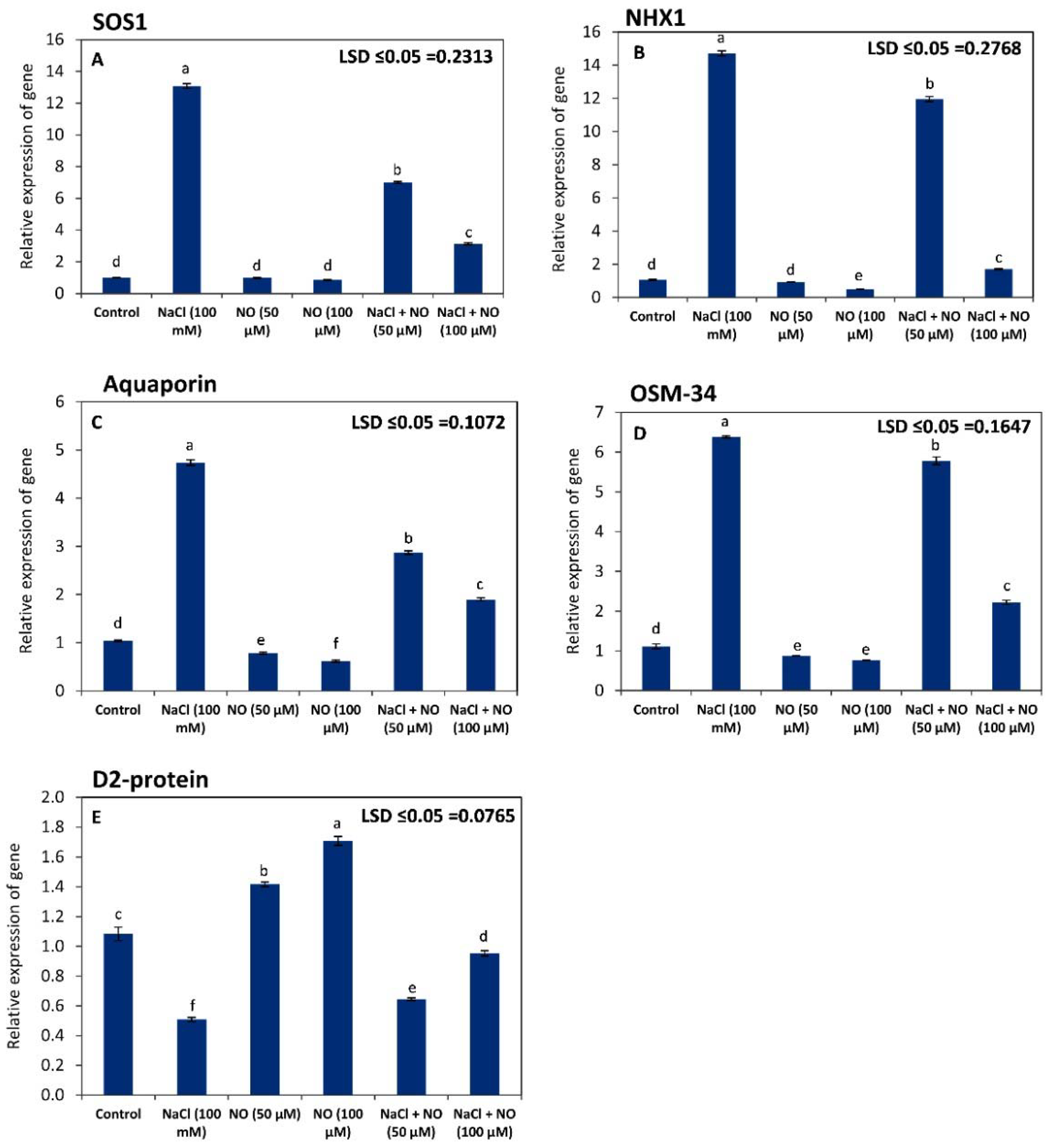
2.7. NO Regulates the Expression of SOS1, NHX1, AQP, OSM-34, and D2-Protein
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Treatment
4.2. Estimation of Photosynthetic Pigments and Measurement of Stomatal Conductance and Photosynthetic Efficiency
4.3. Determination of Leaf Water Content, Proline, and Soluble Sugars
4.4. Measurement of Membrane Stability Index, Lipid Peroxidation, and Hydrogen Peroxide
4.5. Assay of Antioxidant Enzymes
4.6. Determination of Ascorbate and Reduced Glutathione
4.7. Estimation of Mineral Ions
4.8. Gene Expression
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ibrahim, M.F.M.; Elbar, O.H.A.; Farag, R.; Hikal, M.; El-Kelish, A.; El-Yazied, A.A.; Alkahtani, J.; El-Gawad, H.G.A. Melatonin Counteracts Drought Induced Oxidative Damage and Stimulates Growth, Productivity and Fruit Quality Properties of Tomato Plants. Plants 2020, 9, 1276. [Google Scholar] [CrossRef]
- Abd Elhady, S.A.; El-Gawad, H.G.A.; Ibrahim, M.F.M.; Mukherjee, S.; Elkelish, A.; Azab, E.; Gobouri, A.A.; Farag, R.; Ibrahim, H.A.; El-Azm, N.A. Hydrogen Peroxide Supplementation in Irrigation Water Alleviates Drought Stress and Boosts Growth and Productivity of Potato Plants. Sustainability 2021, 13, 899. [Google Scholar] [CrossRef]
- Eid, A.M.; Fouda, A.; Abdel-Rahman, M.A.; Salem, S.S.; Elsaied, A.; Oelmüller, R.; Hijri, M.; Bhowmik, A.; Elkelish, A.; Hassan, S.E.-D. Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview. Plants 2021, 10, 935. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.-D.; Mahgoub, H.A.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A.; et al. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus Vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef] [PubMed]
- Nahhas, N.E.; Abdelaal, K.A.; AlKahtani, M.D.; Al Husnain, L.; AlGwaiz, H.I.; Hafez, Y.M.; Attia, K.A.; El-Esawi, M.A.; Ibrahim, M.F.; Elkelish, A. Biochar and Jasmonic Acid Application Attenuate Antioxidative Systems and Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia Faba L.) Irrigated with Saline Water. Plant Physiol. Biochem. 2021, 166, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra Pachyclada as Plant Growth-Promoting. Biomolecules 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Elsaeed, S.M.; Zaki, E.G.; Ibrahim, T.M.; Ibrahim Talha, N.; Saad, H.A.; Gobouri, A.A.; Elkelish, A.; Mohamed el-kousy, S. Biochar Grafted on CMC-Terpolymer by Green Microwave Route for Sustainable Agriculture. Agriculture 2021, 11, 350. [Google Scholar] [CrossRef]
- Kibria, M.G.; Hossain, M.; Murata, Y.; Hoque, M.A. Antioxidant Defense Mechanisms of Salinity Tolerance in Rice Genotypes. Rice Sci. 2017, 24, 155–162. [Google Scholar] [CrossRef]
- Ibrahim, M.F.M.; Ibrahim, H.A.; Abd El-Gawad, H.G. Folic acid as a protective agent in snap bean plants under water deficit conditions. J. Hortic. Sci. Biotechnol. 2021, 96, 94–109. [Google Scholar] [CrossRef]
- Abd El-Gawad, H.G.; Mukherjee, S.; Farag, R.; Abd Elbar, O.H.; Hikal, M.; Abou El-Yazied, A.; Abd Elhady, S.A.; Helal, N.; ElKelish, A.; El Nahhas, N.; et al. Exogenous γ-Aminobutyric Acid (GABA)-Induced Signaling Events and Field Performance Associated with Mitigation of Drought Stress in Phaseolus Vulgaris L. Plant Signal. Behav. 2021, 16, 1853384. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of Salinity and Water Content on Soil Microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Hashim, A.M.; Elkelish, A.; Alhaithloul, H.A.; El-hadidy, S.M.; Farouk, H. Environmental Monitoring and Prediction of Land Use and Land Cover Spatio-Temporal Changes: A Case Study from El-Omayed Biosphere Reserve, Egypt. Environ. Sci. Pollut. Res. 2020, 27, 42881–42897. [Google Scholar] [CrossRef] [PubMed]
- Bin-Jumah, M.; Abdel-Fattah, A.-F.M.; Saied, E.M.; El-Seedi, H.R.; Abdel-Daim, M.M. Acrylamide-Induced Peripheral Neuropathy: Manifestations, Mechanisms, and Potential Treatment Modalities. Environ. Sci. Pollut. Res. 2021, 28, 13031–13046. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Azab, E.; Hegazy, A.K.; Gobouri, A.A.; Elkelish, A. Impact of Transgenic Arabidopsis Thaliana Plants on Herbicide Isoproturon Phytoremediation through Expressing Human Cytochrome P450-1A2. Biology 2020, 9, 362. [Google Scholar] [CrossRef]
- Din, A.F.; Ibrahim, M.F.; Farag, R.; El-Gawad, H.G.A.; El-Banhawy, A.; Alaraidh, I.A.; Rashad, Y.M.; Lashin, I.; El-Yazied, A.A.; Elkelish, A.; et al. Influence of Polyethylene Glycol on Leaf Anatomy, Stomatal Behavior, Water Loss, and some Physiological Traits of Date Palm Plantlets Grown In Vitro and Ex Vitro. Plants 2020, 9, 1440. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Salem, S.S.; El-Belely, E.F.; Niedbała, G.; Alnoman, M.M.; Hassan, S.E.-D.; Eid, A.M.; Shaheen, T.I.; Elkelish, A.; Fouda, A. Bactericidal and In-Vitro Cytotoxic Efficacy of Silver Nanoparticles (Ag-NPs) Fabricated by Endophytic Actinomycetes and Their Use as Coating for the Textile Fabrics. Nanomaterials 2020, 10, 2082. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M. Salinity Stress Induced Alterations in Antioxidant Metabolism and Nitrogen Assimilation in Wheat (Triticum Aestivum L.) as Influenced by Potassium Supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef]
- Ibrahim, M.F.; El-Samad, A.; Ashour, H.; El-Sawy, A.M.; Hikal, M.; Elkelish, A.; El-Gawad, H.A.; El-Yazied, A.A.; Hozzein, W.N.; Farag, R. Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy 2020, 10, 1212. [Google Scholar] [CrossRef]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Farmer, E.E.; Mueller, M.J. ROS-Mediated Lipid Peroxidation and RES-Activated Signaling. Annu. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar] [CrossRef]
- Soliman, M.; Qari, S.H.; Abu-Elsaoud, A.; El-Esawi, M.; Alhaithloul, H.; Elkelish, A. Rapid Green Synthesis of Silver Nanoparticles from Blue Gum Augment Growth and Performance of Maize, Fenugreek, and Onion by Modulating Plants Cellular Antioxidant Machinery and Genes Expression. Acta Physiol. Plant. 2020, 42, 148. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Rathinasabapathi, B. Oxidative Stress Tolerance in Plants. Plant Signal. Behav. 2013, 8, e25761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, e217037. [Google Scholar] [CrossRef] [Green Version]
- Rashad, Y.; Aseel, D.; Hammad, S.; Elkelish, A. Rhizophagus Irregularis and Rhizoctonia Solani Differentially Elicit Systemic Transcriptional Expression of Polyphenol Biosynthetic Pathways Genes in Sunflower. Biomolecules 2020, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, B.C.; Oelmüller, R. Reactive Oxygen Species Generation and Signaling in Plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.H.; Abdulmajeed, A.M.; Alhaithloul, H.; Alharbi, B.M.; El-Esawi, M.A.; Hasanuzzaman, M.; Elkelish, A. Saponin Biopriming Positively Stimulates Antioxidants Defense, Osmolytes Metabolism and Ionic Status to Confer Salt Stress Tolerance in Soybean. Acta Physiol. Plant. 2020, 42, 114. [Google Scholar] [CrossRef]
- Panieri, E.; Santoro, M.M. ROS Homeostasis and Metabolism: A Dangerous Liason in Cancer Cells. Cell Death Dis. 2016, 7, e2253. [Google Scholar] [CrossRef]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid Seed Priming with Nitrogen Supplementation Improves Salt Tolerance in Soybean. Physiol. Mol. Biol. Plants 2020, 26, 501–511. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma Harzianum Alleviates Waterlogging-Induced Growth Alterations in Tomato Seedlings by Modulating Physiological, Biochemical, and Molecular Mechanisms. Environ. Exp. Bot. 2020, 171, 103946. [Google Scholar] [CrossRef]
- Soliman, M.H.; Alayafi, A.A.M.; El Kelish, A.A.; Abu-Elsaoud, A.M. Acetylsalicylic Acid Enhance Tolerance of Phaseolus Vulgaris, L. to Chilling Stress, Improving Photosynthesis, Antioxidants and Expression of Cold Stress Responsive Genes. Bot. Stud. 2018, 59, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghour, M.; Gálvez, F.J.; Sánchez, M.E.; Aranda, M.N.; Venema, K.; Rodríguez-Rosales, M.P. Overexpression of LeNHX2 and SlSOS2 Increases Salt Tolerance and Fruit Production in Double Transgenic Tomato Plants. Plant Physiol. Biochem. 2019, 135, 77–86. [Google Scholar] [CrossRef]
- Ismail, H.; Maksimović, J.D.; Maksimović, V.; Shabala, L.; Živanović, B.D.; Tian, Y.; Jacobsen, S.-E.; Shabala, S.; Ismail, H.; Maksimović, J.D.; et al. Rutin, a Flavonoid with Antioxidant Activity, Improves Plant Salinity Tolerance by Regulating K+ Retention and Na+ Exclusion from Leaf Mesophyll in Quinoa and Broad Beans. Funct. Plant Biol. 2015, 43, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Falahi, H.; Sharifi, M.; Ahmadian Chashmi, N.; Zare Maivan, H. Water Stress Alleviation by Polyamines and Phenolic Compounds in Scrophularia Striata Is Mediated by NO and H2O2. Plant Physiol. Biochem. 2018, 130, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Du, S.-T.; Liu, Y.; Zhang, P.; Liu, H.-J.; Zhang, X.-Q.; Zhang, R.-R. Atmospheric Application of Trace Amounts of Nitric Oxide Enhances Tolerance to Salt Stress and Improves Nutritional Quality in Spinach (Spinacia oleracea L.). Food Chem. 2015, 173, 905–911. [Google Scholar] [CrossRef]
- Floryszak-Wieczorek, J.; Arasimowicz-Jelonek, M.; Izbiańska, K. The Combined Nitrate Reductase and Nitrite-Dependent Route of NO Synthesis in Potato Immunity to Phytophthora Infestans. Plant Physiol. Biochem. 2016, 108, 468–477. [Google Scholar] [CrossRef]
- Freschi, L. Nitric Oxide and Phytohormone Interactions: Current Status and Perspectives. Front. Plant Sci. 2013, 4, 398. [Google Scholar] [CrossRef] [Green Version]
- Gaber, A.; Alsanie, W.F.; Kumar, D.N.; Refat, M.S.; Saied, E.M. Novel Papaverine Metal Complexes with Potential Anticancer Activities. Molecules 2020, 25, 5447. [Google Scholar] [CrossRef]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed Priming by Sodium Nitroprusside Improves Salt Tolerance in Wheat (Triticum Aestivum L.) by Enhancing Physiological and Biochemical Parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Fatma, M.; Khan, N.A. Nitric Oxide Protects Photosynthetic Capacity Inhibition by Salinity in Indian Mustard. J. Func. Environ. Bot. 2014, 4, 106. [Google Scholar] [CrossRef]
- Melo, N.K.G.; Bianchetti, R.E.; Lira, B.S.; Oliveira, P.M.R.; Zuccarelli, R.; Dias, D.L.O.; Demarco, D.; Peres, L.E.P.; Rossi, M.; Freschi, L. Nitric Oxide, Ethylene, and Auxin Cross Talk Mediates Greening and Plastid Development in Deetiolating Tomato Seedlings. Plant Physiol. 2016, 170, 2278–2294. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Hey, S.J. The Contribution of Wheat to Human Diet and Health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium Protects Wheat Seedlings against Salt Stress-Mediated Oxidative Damage by up-Regulating Antioxidants and Osmolytes Metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Ke, Q.; Wang, Z.; Ji, C.Y.; Jeong, J.C.; Lee, H.-S.; Li, H.; Xu, B.; Deng, X.; Kwak, S.-S. Transgenic Poplar Expressing CodA Exhibits Enhanced Growth and Abiotic Stress Tolerance. Plant Physiol. Biochem. 2016, 100, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Egbichi, I.; Keyster, M.; Ludidi, N. Effect of Exogenous Application of Nitric Oxide on Salt Stress Responses of Soybean. South Afr. J. Bot. 2014, 90, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Shokat, S.; Großkinsky, D.K. Tackling Salinity in Sustainable Agriculture—What Developing Countries May Learn from Approaches of the Developed World. Sustainability 2019, 11, 4558. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Dutta, T.; Neelapu, N.R.R.; Surekha, C. Transgenic Approaches to Enhance Salt and Drought Tolerance in Plants. Plant Gene 2017, 11, 219–231. [Google Scholar] [CrossRef]
- Shi, Q.; Ding, F.; Wang, X.; Wei, M. Exogenous Nitric Oxide Protect Cucumber Roots against Oxidative Stress Induced by Salt Stress. Plant Physiol. Biochem. 2007, 45, 542–550. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.S.P. Nitric Oxide Mediates Hydrogen Peroxide- and Salicylic Acid-Induced Salt Tolerance in Rice (Oryza Sativa L.) Seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Freschi, L.; Corpas, F.J.; Khan, N.A. Nitric Oxide Signaling and Its Crosstalk with Other Plant Growth Regulators in Plant Responses to Abiotic Stress. Environ. Sci. Pollut. Res. Int. 2017, 24, 2273–2285. [Google Scholar] [CrossRef]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought Stress in Maize Causes Differential Acclimation Responses of Glutathione and Sulfur Metabolism in Leaves and Roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-F.; Du, C.-X.; Ding, L.; Xu, Y.-L. Effects of Nitric Oxide on the Germination of Cucumber Seeds and Antioxidant Enzymes under Salinity Stress. Acta Physiol. Plant. 2013, 35, 2707–2719. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous Application of Nitric Oxide Modulates Osmolyte Metabolism, Antioxidants, Enzymes of Ascorbate-Glutathione Cycle and Promotes Growth under Cadmium Stress in Tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef]
- Marvasi, M. Potential Use and Perspectives of Nitric Oxide Donors in Agriculture. J. Sci. Food Agric. 2017, 97, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.J.; Jinc, S.S.; Liu, S.; Xu, L.L.; Kong, J. Effects of Exogenous Nitric Oxide on Growth of Cotton Seedlings under NaCl Stress. J. Soil Sci. Plant Nutr. 2014, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Basalah, M.; Ali, H.; Al-whaibi, M.; Siddiqui, D.M.H.; Sakran, A.; Alsahli, A. Nitric Oxide and Salicylic Acid Mitigate Cadmium Stress in Wheat Seedlings. J. Pure Appl. Microbiol. 2013, 7, 139–148. [Google Scholar]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-María, G.E. Plant Survival in a Changing Environment: The Role of Nitric Oxide in Plant Responses to Abiotic Stress. Front. Plant Sci. 2015, 6, 977. [Google Scholar] [CrossRef]
- Lu, S.; Zhuo, C.; Wang, X.; Guo, Z. Nitrate Reductase (NR)-Dependent NO Production Mediates ABA- and H2O2-Induced Antioxidant Enzymes. Plant Physiol. Biochem. 2014, 74, 9–15. [Google Scholar] [CrossRef]
- Kolbert, Z. Implication of Nitric Oxide (NO) in Excess Element-Induced Morphogenic Responses of the Root System. Plant Physiol. Biochem. 2016, 101, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Pottosin, I. Regulation of Potassium Transport in Plants under Hostile Conditions: Implications for Abiotic and Biotic Stress Tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.M.; Elkelish, A. Beneficial Features of Biochar and Arbuscular Mycorrhiza for Improving Spinach Plant Growth, Root Morphological Traits, Physiological Properties, and Soil Enzymatic Activities. JoF 2021, 7, 571. [Google Scholar] [CrossRef]
- Elkelish, A.; Ibrahim, M.F.M.; Ashour, H.; Bondok, A.; Mukherjee, S.; Aftab, T.; Hikal, M.; El-Yazied, A.A.; Azab, E.; Gobouri, A.A.; et al. Exogenous Application of Nitric Oxide Mitigates Water Stress and Reduces Natural Viral Disease Incidence of Tomato Plants Subjected to Deficit Irrigation. Agronomy 2021, 11, 87. [Google Scholar] [CrossRef]
- Turan, S.; Tripathy, B.C. Salt-Stress Induced Modulation of Chlorophyll Biosynthesis during de-Etiolation of Rice Seedlings. Physiol. Plant. 2015, 153, 477–491. [Google Scholar] [CrossRef]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric Oxide Alleviates Salt Stress Inhibited Photosynthetic Performance by Interacting with Sulfur Assimilation in Mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar] [CrossRef] [Green Version]
- Procházková, D.; Haisel, D.; Wilhelmová, N.; Pavlíková, D.; Száková, J. Effects of Exogenous Nitric Oxide on Photosynthesis. Photosynthetica 2013, 51, 483–489. [Google Scholar] [CrossRef]
- Azooz, M.; Youssef, A.; Ahmad, P. Evaluation of Salicylic Acid (SA) Application on Growth, Osmotic Solutes and Antioxidant Enzyme Activities on Broad Bean Seedlings Grown under Diluted Seawater. Inter. J. Plant Physiol. Biochem. 2011, 3, 253–264. [Google Scholar] [CrossRef]
- Bahrami-Rad, S.; Hajiboland, R. Effect of Potassium Application in Drought-Stressed Tobacco (Nicotiana Rustica L.) Plants: Comparison of Root with Foliar Application. Ann. Agric. Sci. 2017, 62, 121–130. [Google Scholar] [CrossRef]
- Munns, R. Comparative Physiology of Salt and Water Stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Kaldenhoff, R.; Ribas-Carbo, M.; Sans, J.F.; Lovisolo, C.; Heckwolf, M.; Uehlein, N. Aquaporins and Plant Water Balance. Plant Cell Environ. 2008, 31, 658–666. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of Salinity Stress on Vicia Faba L. Plants via Seed Priming with Melatonin. Acta Biológica Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Abdelgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, A.; Abu-Elsaoud, A.; El Kelish, A.; Sahadad, M.; Mohamed, E. Role of External Proline on Enhancing Defence Mechanisms of Vicia Faba L. against Ultraviolet Radiation. Am. Eurasian J. Sustain. Agric. 2015, 9, 22–34. [Google Scholar]
- Elkahoui, S.; Hernández, J.A.; Abdelly, C.; Ghrir, R.; Limam, F. Effects of Salt on Lipid Peroxidation and Antioxidant Enzyme Activities of Catharanthus Roseus Suspension Cells. Plant Sci. 2005, 168, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.; Fernandez-Garcia, N.; Diaz-Vivancos, P.; Olmos, E. A Different Role for Hydrogen Peroxide and the Antioxidative System under Short and Long Salt Stress in Brassica Oleracea Roots. J. Exp. Bot. 2010, 61, 521–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco-Ríos, L.; Pinto, M. Effect of Salt Stress on Antioxidant Enzymes and Lipid Peroxidation in Leaves in Two Contrasting Corn, ‘Lluteno’ and ‘Jubilee’. Chil. J. Agric. Res. 2014, 74, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Lamattina, L.; García-Mata, C.; Graziano, M.; Pagnussat, G. Nitric Oxide: The Versatility of an Extensive Signal Molecule. Annu. Rev. Plant Biol. 2003, 54, 109–136. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Functions of Nitric Oxide (NO) in Roots during Development and under Adverse Stress Conditions. Plants 2015, 4, 240–252. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Dutta, D.; Esmaili, M.; Overduin, M.; Fliegel, L. Expression and Detergent Free Purification and Reconstitution of the Plant Plasma Membrane Na+/H+ Antiporter SOS1 Overexpressed in Pichia Pastoris. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183111. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The Roles of Aquaporins in Plant Stress Responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim; Ullah, A.; Hussain, A.; Shaban, M.; Khan, A.H.; Alariqi, M.; Gul, S.; Jun, Z.; Lin, S.; Li, J.; et al. Osmotin: A Plant Defense Tool against Biotic and Abiotic Stresses. Plant Physiol. Biochem. 2018, 123, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.A.; Silvestri, C.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A.; Manzoor, A.; Cristofori, V.; Rugini, E. Osmotin: A Cationic Protein Leads to Improve Biotic and Abiotic Stress Tolerance in Plants. Plants 2020, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Kale, R.; Hebert, A.E.; Frankel, L.K.; Sallans, L.; Bricker, T.M.; Pospíšil, P. Amino Acid Oxidation of the D1 and D2 Proteins by Oxygen Radicals during Photoinhibition of Photosystem II. Proc. Natl. Acad. Sci. USA 2017, 114, 2988–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, V.; Jhanghel, D.; Mehrotra, S. Emerging Warriors against Salinity in Plants: Nitric Oxide and Hydrogen Sulphide. Physiol. Plant. 2021, 171, 896–908. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Inafuku, M.; Nahar, K.; Fujita, M.; Oku, H. Nitric Oxide Regulates Plant Growth, Physiology, Antioxidant Defense, and Ion Homeostasis to Confer Salt Tolerance in the Mangrove Species, Kandelia Obovata. Antioxidants 2021, 10, 611. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Hiscox, J.D.; Israelstam, G.F. Erratum: A Method for the Extraction of Chlorophyll from Leaf Tissue without Maceration. Can. J. Bot. 2011, 58, 403. [Google Scholar] [CrossRef]
- Smart, R.E.; Bingham, G.E. Rapid Estimates of Relative Water Content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International: New Delhi, India, 1996; ISBN 978-81-224-0976-5. [Google Scholar]
- Shields, R.; Burnett, W. Determination of Protein-Bound Carbohydrate in Serum by Modified Anthrone Method. Anal. Chem. 1960, 32, 885–886. [Google Scholar] [CrossRef]
- Ibrahim, M.; Bondok, A.; Al-Senosy, N.K.; Younis, R. Stimulation some of defense mechanisms in tomato plants under water deficit and Tobacco mosaic virus (TMV). World J. Agric. Sci. 2015, 11, 289–302. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Cheeseman, J.M. Hydrogen Peroxide Concentrations in Leaves under Natural Conditions. J. Exp. Bot. 2006, 57, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Oxygen Radicals in Biological Systems; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant. Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. In Methods in Enzymology; Glutamate, Glutamine, Glutathione, and Related Compounds; Academic Press: Cambridge, MA, USA, 1985; Volume 113, pp. 484–490. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of Water Stress-Induced Changes in the Levels of Endogenous Ascorbic Acid and Hydrogen Peroxide in Vigna Seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Wolf, B. A Comprehensive System of Leaf Analyses and Its Use for Diagnosing Crop Nutrient Status. Commun. Soil Sci. Plant. Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Subbiah, B.V.; Asija, G.L. A Rapid Procedure for the Estimation of Available Nitrogen in Soils. Curr. Sci. 1956, 25, 259–260. [Google Scholar]
- Togawa, T.; Dunn, W.A.; Emmons, A.C.; Nagao, J.; Willis, J.H. Developmental Expression Patterns of Cuticular Protein Genes with the R&R Consensus from Anopheles Gambiae. Insect Biochem. Mol. Biol. 2008, 38, 508–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoot Length (cm) | Shoot Fresh Weight (gm) | Shoot Dry Weight (gm) | |
|---|---|---|---|
| Control | 22.26 ± 1.006c | 4.573 ± 0.417c | 1.206 ± 0.103bc |
| NaCl (100 mM) | 14.09 ± 0.899e | 2.725 ± 0.453f | 0.7071 ± 0.012e |
| NO 50 µM | 25.96 ± 2.65b | 4.805 ± 0.116b | 1.386 ± 0.055b |
| NO 100 µM | 28.73 ± 1.08a | 6.068 ± 0.073a | 2.034 ± 0.06a |
| NaCl + NO 50 µM | 16.06 ± 0.125d | 3.256 ± 0.169e | 0.944 ± 0.05d |
| NaCl + NO 100 µM | 17.97 ± 0.047d | 4.025 ± 0.18d | 1.103 ± 0.1c |
| N (mg g−1) | Na (mg g−1) | K (mg g−1) | Ca (mg g−1) | Na/K % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Root | Leaf | Root | Leaf | Root | Leaf | Root | Leaf | Root | |
| Control | 35.37 ± 1.05c | 17.46 ± 1.02c | 7.072 ± 0.51d | 9.98 ± 0.87d | 45.64 ± 0.74c | 26.55 ± 1.44c | 6.78 ± 0.502c | 4.91 ± 0.54c | 0.154 | 0.375 |
| NaCl (100 mM) | 25.71 ± 1.05f | 13.06 ± 0.96d | 15.39 ± 1.15a | 17.69 ± 0.6a | 25.07 ± 0.27f | 17.92 ± 0.89f | 4.02 ± 0.327ef | 3.21 ± 0.29ef | 0.613 | 0.987 |
| NO 50 µM | 38.59 ± 0.61b | 19.3 ± 1.12b | 6.55 ± 0.37d | 7.66 ± 0.42e | 51.24 ± 1.22b | 30.02 ± 1.04b | 7.35 ± 0.168b | 5.56 ± 0.38b | 0.127 | 0.255 |
| NO 100 µM | 43.69 ± 1.13a | 25.31 ± 0.97a | 4.86 ± 0.08e | 5.51 ± 0.55f | 61.13 ± 1.12a | 34.75 ± 1.03a | 9.5 ± 0.465a | 6.93 ± 0.14a | 0.079 | 0.158 |
| NaCl + NO 50 µM | 27.95 ± 0.11e | 14.92 ± 0.97d | 13.18 ± 0.61b | 15.29 ± 1.01b | 31.41 ± 0.6e | 19.43 ± 0.65e | 4.58 ± 0.302e | 3.6 ± 0.51e | 0.419 | 0.786 |
| NaCl + NO 100 µM | 33.48 ± 0.44cd | 16.68 ± 1.08c | 10.6 ± 0.53c | 12.71 ± 0.54c | 37.85 ± 0.9d | 24.25 ± 0.58cd | 5.93 ± 0.153d | 4.56 ± 0.4cd | 0.28 | 0.524 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alnusairi, G.S.H.; Mazrou, Y.S.A.; Qari, S.H.; Elkelish, A.A.; Soliman, M.H.; Eweis, M.; Abdelaal, K.; El-Samad, G.A.; Ibrahim, M.F.M.; ElNahhas, N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants 2021, 10, 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081693
Alnusairi GSH, Mazrou YSA, Qari SH, Elkelish AA, Soliman MH, Eweis M, Abdelaal K, El-Samad GA, Ibrahim MFM, ElNahhas N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants. 2021; 10(8):1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081693
Chicago/Turabian StyleAlnusairi, Ghalia S. H., Yasser S. A. Mazrou, Sameer H. Qari, Amr A. Elkelish, Mona H. Soliman, Mohamed Eweis, Khaled Abdelaal, Gomaa Abd El-Samad, Mohamed F. M. Ibrahim, and Nihal ElNahhas. 2021. "Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat" Plants 10, no. 8: 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081693